TcLr35小麦中抗病相关基因S2A2的抗叶锈性分析
2017-05-17张艳俊张家瑞栗小英王海燕刘大群
张艳俊 张家瑞 栗小英 王海燕 刘大群


摘要:NBS-LRR是已克隆植物抗病基因的高度保守氨基酸区域。前期工作中,笔者成功克隆获得了1个通读的NBS类抗病同源基因S2A2的cDNA序列,该序列含有NB-ARC保守结构域和多个LRR结构域,且在小麦叶片中为低丰度组成型表达。进一步根据S2A2基因在TcLr35与Thatcher中扩增获得的基因序列差异位点设计特异性引物,分别以TcLr35、Thatcher为模板进行扩增,筛选出了具有较高稳定性和可重复性的3对引物。利用3个多态性引物对TcLr35、Thatcher及其F2代群体进行扩增和遗传性分析,并用Mapmanager软件计算分子标记与抗叶锈病基因之间的遗传距离,结果发现这3对引物获得的标记与Lr35基因遗传距离较远。利用这3个多态性引物扩增33个不同小麦抗叶锈病近等基因系材料,并回收测序,结果表明该基因序列在不同近等基因系材料中广泛存在。
关键词:NBS-LRR;抗病基因;小麦叶锈病;分子标记
中图分类号:S435.12 文献标志码:A 文章编号:1002—1302(2016)01—0148—04
小麦是世界上分布范围最广,栽培面积最大,总产量最高的粮食作物。病害是威胁小麦稳产高产的主要因素之一。由专性寄生真菌——小麦叶锈菌(Puccinia triticina)引起的小麦叶锈病是最严重的小麦叶部病害之一,在世界各产麦区均有发生。在北非、东南亚、中亚、东欧和南北美洲等广大地区较为严重。该病也是威胁我国小麦生产的一种常发病害,20世纪70年代,北方麦区曾发生3次中度以上流行(1973年、1975年和1979年),给我国小麦生产造成严重的损失。过去几年中我国虽然没有叶锈病大规模暴发的相关报道,但在华北及黄淮麦区,叶锈病经常对小麦生产造成危害,近年来该病害呈上升趋势。开发利用寄主本身的抗病性是解决病害问题的根本途径,但由于叶锈菌毒性基因的高度变异性和抗锈品种的不合理使用,导致传统抗病品种的抗性丧失。因此,利用包括DNA分子标记技术在内的多种手段提高抗病基因和抗病类型的丰富度、保持品种抗锈性的持久有效,是一项十分紧迫的任务。
成株抗叶锈病基因Lr35最初来源于拟斯卑尔脱山羊草(Aegilops spehoides),通过与二倍体的Triticum monococcum回交转到小麦中,定位在2B染色体上,与Sr39紧密连锁。国内外至今尚未发现对它表现毒性的菌株存在,是一个应用潜力很大的由主效基因控制的抗病基因。目前人们已经找到了与Lr35紧密连锁的BCD260和UBC836这2个分子标记,并分别转化为更为稳定的STS和SCAR特异性标记。笔者所在实验室王海燕等在TcLr35小麦中获得$2A2基因全长,序列比较结果表明,与Thatcher中获得的该基因序列差异明显,含有多个插入/缺失或者SNP位点,为了能快速且恰当的评价该基因的功能及其与小麦叶锈病的抗病相关性,本研究利用特异性引物筛选抗感材料和不同近等基因系材料差异,以期明确S2A2基因抗叶锈性。
1材料与方法
1.1材料
供试小麦叶锈菌致病菌株99-8-11-5-3由河北农业大学小麦叶锈病研究中心提供。小麦抗叶锈病近等基因系材料TcLr35、感病亲本Thatcher和郑州5389及其他抗叶锈病近等基因系均来自河北农业大学小麦叶锈病研究室。
1.2特异性引物的筛选
围绕Thatcher和TcLr35中获得的序列差异位点设计特异性引物(表1),将其自由组合成不同引物组合,并以TcLr35和Thatcher为模板进行筛选。引物筛选的24μL反应体系为:1.0μL模板,18.7μL ddH2O,2.5μL 10×buffer,0.5μLdNTP,正反向引物各0.5μL,0.3μL Taq酶。反应程序为94℃预变性1 min;94℃变性30 s,58℃退火1 min,72℃延伸2 min,30个循环;最后72℃延伸10 min。扩增产物用1.0%琼脂糖凝胶电泳检测,用凝胶成像分析系统进行拍照及分析。
1.3抗叶锈病分析
Lr35基因为成株抗叶锈病基因,待供试小麦生长至4叶期,采用撒粉法接种新鲜小麦叶锈菌种。接种后15 d左右,待发病充分时进行抗感鉴定,按0、;、1、2、3、4等6级标准调查记载表现型,0~2级为抗病,3~4级为感病。同时利用Lr35的SCAR引物进行抗病性鉴定。24μL反应体系为:模板1.0μL,ddH20 18.7μL,10×buffer 2.5μL,dNTF0.5μL,正反向引物各0.5μL,Taq酶0.3μL。反应程序为:94℃预变性1 min;94℃变性30 s,64℃退火1 min,72℃延伸2 min,35个循环;最后72℃延伸10 min。扩增产物用1.0%琼脂糖凝胶电泳检测,用凝胶成像分析系统进行拍照及分析。
利用小麦近等基因系寻找TcLr35的多态条带,再用F2分离后代单株,验证该多态性条带与目标基因连锁程度。用Mapmanager软件对数据进行连锁分析,计算遗传距离。
2结果与分析
2.1特异性引物的筛选
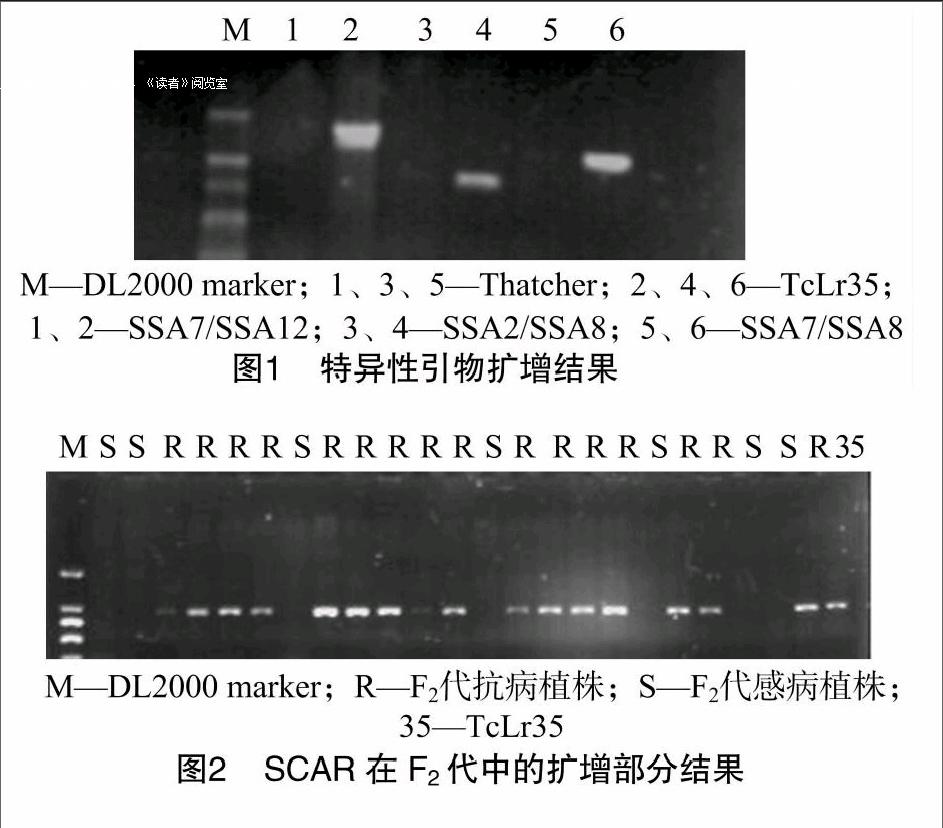
为了进一步明确该基因与Lr35的关系,根据S2A2基因在TcLr35和Thatcher中序列的差异性位点设计引物组合进行筛选,获得3条特异性条带,即3对引物组合能扩增出小麦抗叶锈病近等基因系TcLr35特异性条带,而在Thatcher中条带缺失,这3对引物分别是SSA7/SSA8、SSA2/SSA8、SSAT/SSA12。引物对SSA7/SSA8能在TcLr35中稳定扩增出1条大小为1200 bp左右的条带;引物对SSA2/SSA8能在TcLr35中稳定扩增出1条大小为890 bp左右的条带;引物对SSA7/SSA12能在TcLr35中稳定扩增出1条大小为1300 bp左右的条带(图1)。经多次試验,扩增结果具有很高的稳定性和可重复性。
2.2小麦叶锈病的鉴定及群体遗传分析
叶锈菌生理小种99-8-11-5-3接种TcLr35、感病亲本Thatcher及杂交F1代10个单株。抗病鉴定结果表明,TcLr35和10个杂交F1代单株对生理小种99-8-11-5-3表现为高抗(反应型为0),而Thatcher表现为高感(反应型为4)。
成株期对192个F2代单株接种小麦叶锈菌生理小种99-8-11-5-3后,抗病146株,感病46株,用卡方测验(x2-test)检验其适合度,其卡方值是0.11,检验结果表明,该基因符合3:1的遗传规律,说明F2代中的Lr35基因以单显性基因方式存在。为保证鉴定结果的可靠性,用Lr35的SCAR引物对192株F2(TcLr35×Thatcher)代群体抗病性进行鉴定,结果在抗病品种中扩出了Lr35基因的特异条带而在感病品种中没有(图2),结果与接菌鉴定一致。
为检测目的基因位点与所获得标记位点的遗传连锁性,利用3个多态性引物检测F2代群体单株,其结果显示,引物对SSA7/SSA8在46株感病中有4株扩出大小为1200 bp的差异性条带,而在146株抗病中有4株没有扩出差异性条带(图3);引物对SSA7/SSA12在46株感病中有5株扩出大小为1290 bp的差异性条带,而在146株抗病中有3株没有扩出差异性条带(图4);引物对SSA2/SSA8在46株感病中由5株扩出890 bp差异性条带,而在146株抗病中有4株没有扩出差异性条带(图5),表明在后代群体中3个标记交换值均较高。用Mapmanager软件计算分子标记与抗叶锈病基因之间的遗传距离,结果发现这3对引物获得的标记与Lr35基因遗传距离较远。
2.3特异性引物扩增近等基因系
用3对特异性引物组合分别对33个小麦抗叶锈基因系材料进行PCR扩增(图6、图7、图8),结果显示,引物组合SSA7/SSA8、SSA7/SSA12除在TcLr14a、TcLrl4b、TcLr14ab、TcLr41、TcLr42和TcLrl8中没有获得扩增条带外,其余27个近等基因系中均扩增出目的条带;引物组合SSA2/SSA8除在TcLr14a、TcLr14b、TcLr14ab、TcLr41、TcLr42、TcLr11和TcLr18中没有条带外,其余26个近等基因系中均扩增出目的条带,说明3个特异性条带在大多数近等基因系材料中存在,选取不同来源近等基因系材料中获得的差异性条带回收测序,所得序列与TcLr35的差异性条带序列差异不大,表明获得的序列在近等基因系材料中普遍存在。
3讨论
抗叶锈病基因的利用,在小麦抗叶锈病育种中发挥了重要的作用。迄今为止,国际上已发现100余个小麦抗叶锈基因,正式命名编号至Lr72,其中大部分已分子作图并获得紧密连锁或共分离的分子标记(Lr1、Lr2c、Lr12、Lr25、Lr32、Lr37、Lr38、Lr41、Lr42、Lr48和LrZH84,但由于气候条件的变化、生产上的局限性及新的叶锈菌生理小种的形成,导致许多抗锈品种反应型由近免疫到了1或2型甚至4型。如Lr1基因对小麦大部分叶锈菌生理小种已表现感病,Lr2c基因毒性频率也下降等等。培育成株抗性品种将会成为控制小麦叶锈病经济与环保相结合的有效方法。小麦抗叶锈病基因Lr35是一个成株抗性基因,且尚未报道发现其毒性小种,是有效的成株抗性基因,吸引了大量的研究者。本实验室先后对其抗病机制、分子标记、辅助育种等方面进行了研究,并取得了一定的进展。本研究旨在前期工作的基础上进一步明确S2A2基因与小麦抗叶锈性的关系。
研究发现试图应用特异性引物在抗感材料间发现带型上的差异不太可能。可能是单碱基的差异,而不是碱基数目的差异,这是不能用普通的电泳带型区分的,这与已克隆的小麦抗叶锈病基因Lr34特点类似。本研究根据S2A2基因序列设计全长引物,在Thatcher和TcLr35基因组中分别获得该基因扩增条带,进行了全长测序,发现S2A2基因在抗感材料问序列差异明显,尤其3′端差异显著,含有多个较长的插入、缺失序列,而大多数基因的功能主要决定于3′端。
为了揭示$2A2基因与Lr35相关性,根据抗感材料中序列差异明显区域,尤其围绕2个内元设计特异性引物,有3个引物能够揭示TcLr35和Thatcher抗感亲本间差异,而在F2代验证中感病单株也出现了与抗病亲本相同的特异条带,郭楠等和闫红飞等分别在获得Lr45、Lr38的分子标记时也具有该现象,可能是由单核苷酸位点突变引起的重组突变或染色体交叉导致的,但其交换值较高,这些标记与Lr35基因遗传距离较远。当然这些引物扩增的仅是S2A2基因全长中的一部分序列,可能为遗传背景中共同的遗传信息,由于小麦抗叶锈病基因来源比较复杂,如Lr19来源于长穗偃麦草(Thinopyrum elongatum),Lr35来源于拟斯比尔脱山羊草(Ae-gilops speltoides),Lr38来源于中间偃麦草(T.interrnedium),Lr41和Lr42基因来源于粗山羊草(T.tauschii)等等,转入普通小麦后,经过6代回交,可能会产生一些共同的遗传物质,如果筛选更多的特异性引物,可能会获得与Lr35紧密连锁或共分离的分子标记。同时利用这些特异性引物在一部分小麦抗叶锈病近等基因系材料中可以获得相同的序列,在一部分小麦抗叶锈病近等基因系材料中不能获得有效扩增片段,说明这些序列在多数近等基因系基因组中广泛存在,而且这种差异能在抗感材料间表现。由于小麦基因组的复杂性,以及Lr35目的基因本身来源于外源导入片段,从以上的研究结果推测,S2A2基因可能不是Lr35目的基因,但其中的一些序列在多数小麦抗叶锈病近等基因系材料中普遍存在,而且可能与小麦抗叶锈性相关,可能为多数小麦抗叶锈病反应中的调控基因,该結果尚待在稳定转化中进行进一步的功能验证。
