不同郁闭度控制下马尾松(Pinusmassoniana)人工林土壤动物群落特征
2017-05-15周泓杨张丹桔魏大平赵燕波李川北
周泓杨,张 健,,*,张丹桔,张 捷,魏大平,赵燕波,赵 波,李川北
1 四川农业大学生态林业研究所, 林业生态工程省级重点实验室 成都 611130 2 四川农业大学林学院, 成都 611130
不同郁闭度控制下马尾松(Pinusmassoniana)人工林土壤动物群落特征
周泓杨1,张 健1,2,*,张丹桔1,张 捷1,魏大平1,赵燕波1,赵 波2,李川北2
1 四川农业大学生态林业研究所, 林业生态工程省级重点实验室 成都 611130 2 四川农业大学林学院, 成都 611130
马尾松人工林; 郁闭度; 土壤动物; 群落结构
土壤动物作为森林生态系统的重要组成部分,与地下植物根系、微生物等一起构成完整稳定的食物网络,在人工林土壤活性及养分循环方面发挥了积极作用[1]。土壤动物取食、掘穴等活动直接促进枯落物分解和粉碎,增加了土壤有机元素的有效性,促进微生物多样性和土壤微环境异质性,能够加速养分循环[2],影响土壤有机质稳定性和碳、氮平衡[3]。土壤动物与土壤微生物及地上部分的相互关系有利于森林生态系统功能的发挥[4]。因此,研究森林生态系统土壤动物群落结构和功能具有十分重要的意义。目前,关于人工林经营与生态系统中土壤动物群落结构和功能的研究已有大量报道,如Andrey[5]等研究了云杉纯林经过4个演替阶段发展成混交林对土壤甲螨群落结构的影响。张良辉[6]研究了川西亚高山米亚罗林区云杉人工林不同大小林窗以及不同强度抚育间伐调控下植物和土壤动物多样性。而关于人工林不同郁闭度的生态功能研究还相对较少[7]。
马尾松(Pinusmassoniana)作为我国南方和长江流域低山丘陵区主要造林树种,面积超过200万hm2,在人工林中占有很大比例[8]。然而,马尾松人工林林下植被结构单一、生物多样性降低[9];马尾松凋落物腐化分解产生的有机酸等导致土壤酸化,降低了部分养分元素有效性[10];连栽马尾松林使土壤速效氮、速效磷、代换性钙镁离子含量明显下降,土壤酶类活性及微生物区系随之相应变化,从而影响整个马尾松林下土壤代谢活动[11]。因此,采取科学的抚育措施加强马尾松人工林经营的要求非常迫切。林分郁闭度作为反映森林结构和环境状态的重要因子,在林分结构构建与调控过程中为确定抚育措施和抚育强度提供参考[12]。通过郁闭度调节林内光照和水分,减小森林生态系统中光合有效辐射、土壤温度、土壤含水量、空气湿度和温度的极端分布,为林下植被发育和生物活动创造稳定良好的小生境[13]。土壤动物对生存环境的感受与适应,决定了郁闭度变化也显然会影响到土壤生态系统中土壤动物的群落结构组成。而目前关于不同郁闭度对土壤生态系统尤其是对土壤动物的研究还未见报道,难以完整全面地解释郁闭度生态效应及其响应机制。
土壤动物群落受气温和降雨的季节变化,在亚热带针叶林中通常于秋季大量发生[14]。为此,本文选择四川宜宾11年生马尾松人工林作为研究对象,通过抚育间伐结合修枝控制形成5个郁闭度梯度(0.5、0.6、0.7、0.8、0.9),于2014年10月在不同郁闭度下马尾松人工林进行土壤动物群落组成、结构及多样性研究,以期为马尾松人工林的科学经营和管理提供依据。
1 研究区概况
研究区位于四川省高县来复镇(28°11′—28°47′ N,104°21′—104°48′ E),地处四川盆地南缘。该地属中亚热带湿润季风气候,气候温和,雨量充沛。年平均气温18.1 ℃,最高气温(7月)36.8 ℃,最低气温(1月)7.8 ℃,年均降雨量1 021 mm。相对湿度81%,年日照时数1 148 h。地貌以低山丘陵为主,海拔412—567 m。土壤类型为发育在砂岩上的山地黄壤。植被绝大部分为原始植被受破坏后的次生植被,马尾松人工林林分结构简单,初植密度较大。常见林下灌木有枹栎(Quercusserrate)、黄牛奶(Symplocoslaurina)、梨叶悬钩子(Rubuspirifolius)等,草本层有淡竹叶(Lophatherumgracile),金星蕨(Parathelypterisglanduligera)、芒萁(Dicranopterisdichotoma)、山菅兰(Dianellaensifolia)等。
2 材料与方法
2.1 样地设置
本次实验样地位于高县来复镇森林经营所毛颠坳V1林班大窝场。2013年9月,在已有马尾松人工林(2.84 hm2,郁闭度0.9,林龄11 a,栽植密度1.5 m×1.5 m)的基础上通过间伐、修枝形成郁闭度为0.5、0.6、0.7、0.8、0.9梯度系列样地。郁闭度采用数字式植物冠层分析仪(CI- 110)测定。每个处理林分面积约为20 m×20 m。每个郁闭度设置3个重复样地。每个样地间隔10 m,样地边界离林缘不小于10 m。样地基本情况见表1,其中土壤性质的测定于2014年10月与土壤动物样品采集同步进行,以“品”字形采样法采集0—15 cm层土壤混合样品并测定。
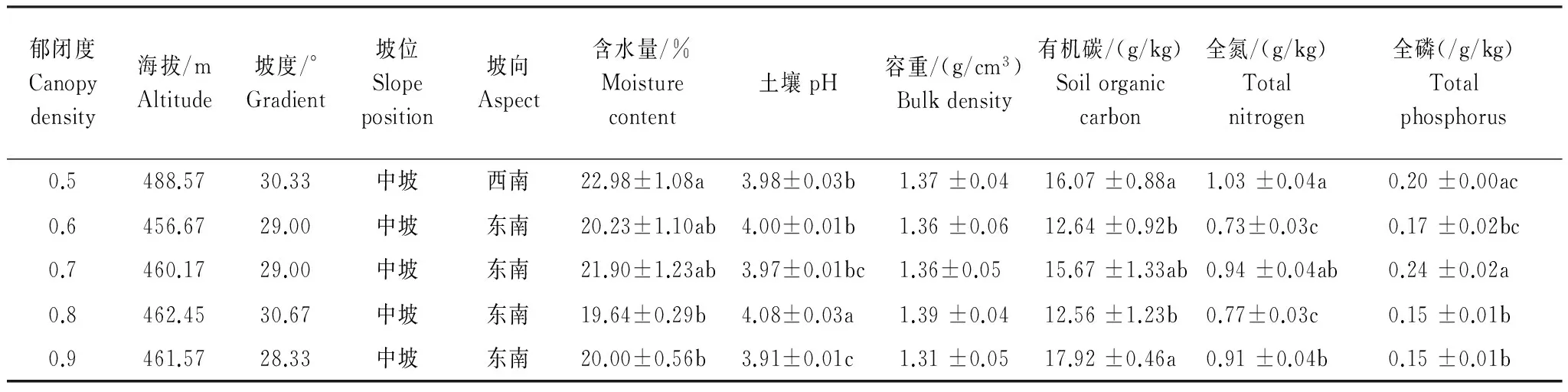
表1 高县来复不同郁闭度马尾松人工林样地概况(平均值±标准误)
同列不同小写字母表示差异显著(P<0.05)
2.2 样品采集与分离
2014年10月,在样方内以“五点法”进行采样,样点选择较为平坦、人为活动少且避开斜坡地、洼地、岩石、倒木和大树根。每个采样点分为枯落物层、0—5 cm、5—10 cm 3个不同层次取样。土壤动物按体形、习性及其在食物分解过程中的作用分为大型和中小型两类[15- 16]。其中,大型土壤动物主要是指野外调查中肉眼可见的在浅层土壤活动的大型种类,采用手捡法,采样面积为50 cm×50 cm(0.25 m2),将收集到的土壤动物放入盛有浓度为75%的酒精容器中杀死后带回实验室在解剖镜下分类计数;中小型土壤动物包括用湿漏斗法(Baermann法)收集的小型湿生土壤动物和用干漏斗法(Tullgren法)收集的中小型节肢动物。首先在采样点收集10 cm×10 cm(0.01 m2)枯落物带回实验室用Tullgren法分离土壤动物,再用环刀(100 mL)自下往上顺次取土,每个土层取两个土样,带回实验室分别用Tullgren干漏斗和Baermann湿漏斗分离。由于湿生土壤动物平均密度较大,用于分离湿生的土壤样品采用“四分法”取四分之一(25 mL)进行分离[17]。
土壤动物的分离在烘虫箱中进行,控制烘虫箱温度在35—50 ℃,烘虫时间为48 h,其中湿生土壤动物在分离4 h后观察一次,以防线虫自溶。干漏斗法分离出的土壤动物用盛有75%浓度的酒精收集于培养皿中,湿生土壤动物则用清水。对分离出的土壤动物进行分类鉴定并计数。
土壤动物的分类鉴定,将捕获所得的土壤动物置于双目解剖镜(Leica,EZ4HD)下,主要参考《中国土壤动物检索图鉴》、《中国亚热带土壤动物》、《昆虫分类检索》和《幼虫分类学》进行分类鉴定。受分类限制,采用大类进行分类。
2.3 数据处理与分析
土壤动物类群数量等级划分依据:个体数量占捕获总量的10.0%以上者为优势类群(+++), 占1.0%—10.0%者为常见类群(++), 不足1.0%者为稀有类群(+)。
群落多样性分析:土壤动物多样性特征主要采用Shannon-Wiener多样性指数(H′)、Margalef丰富度指数(D)、Pielou均匀度指数(J)和Simpson优势度指数(C)来计算。
Simpson优势度指数(C):
Shannon-Wiener多样性指数(H′):
Pielou均匀度指数(E):
E=H′/lnS
Margalef丰富度指数(D):
D=(S-1)/lnN
式中,Ni为第i类群的个体数;N为群落中所有类群的个体数;Pi=Ni/N,S为类群数。
群落相似性分析:采用Jaccard相似性指数
Q=c/a+b-c
式中,a为A样地全部类群数,b为B样地全部类群数,c为A、B样地共有的类群数。主要用于反映林分之间土壤动物类群上的相似程度。
数据的整理与分析主要采用Excel 2010和SPSS 22.0,采用Origin 8.1 绘制图形。利用单因素方差分析(one-way ANOVA)比较不同郁闭度间土壤动物群落的差异性,用最小显著差异法(LSD)进行两两比较。用Canoco for Windows 4.5对土壤动物和主要土壤环境因子进行CCA分析。
3 结果与分析
3.1 土壤动物群落组成
3.2 土壤动物功能类群
根据林英华[18]等,将该研究区土壤动物划分为腐食性、杂食性、植食性、捕食性、菌食性、枯食性和粪食性土壤动物。大型土壤动物以杂食性为主,占56.03%,其次是捕食性(19.19%)>植食性(10.19%)>腐食性(8.49%)>枯食性(5.94%)>粪食性(0.17%)(图1A)。其中杂食性和捕食性在0.5—0.8无显著差异,在0.8—0.9捕食性显著减少,杂食性显著增加。中小型土壤动物以菌食性(45.08%)和腐食性(40.19%)为主,其次为杂食性(9.90%)>植食性(4.23%)>捕食性(0.48%)>枯食性(0.11%)(图1B)。其中捕食性在0.5—0.7显著增加,0.7—0.9无显著变化;随郁闭度增大,腐食性土壤动物逐渐增加,菌食性逐渐减少,且都在0.9郁闭度下形成显著性差异(P<0.05;P<0.01)。
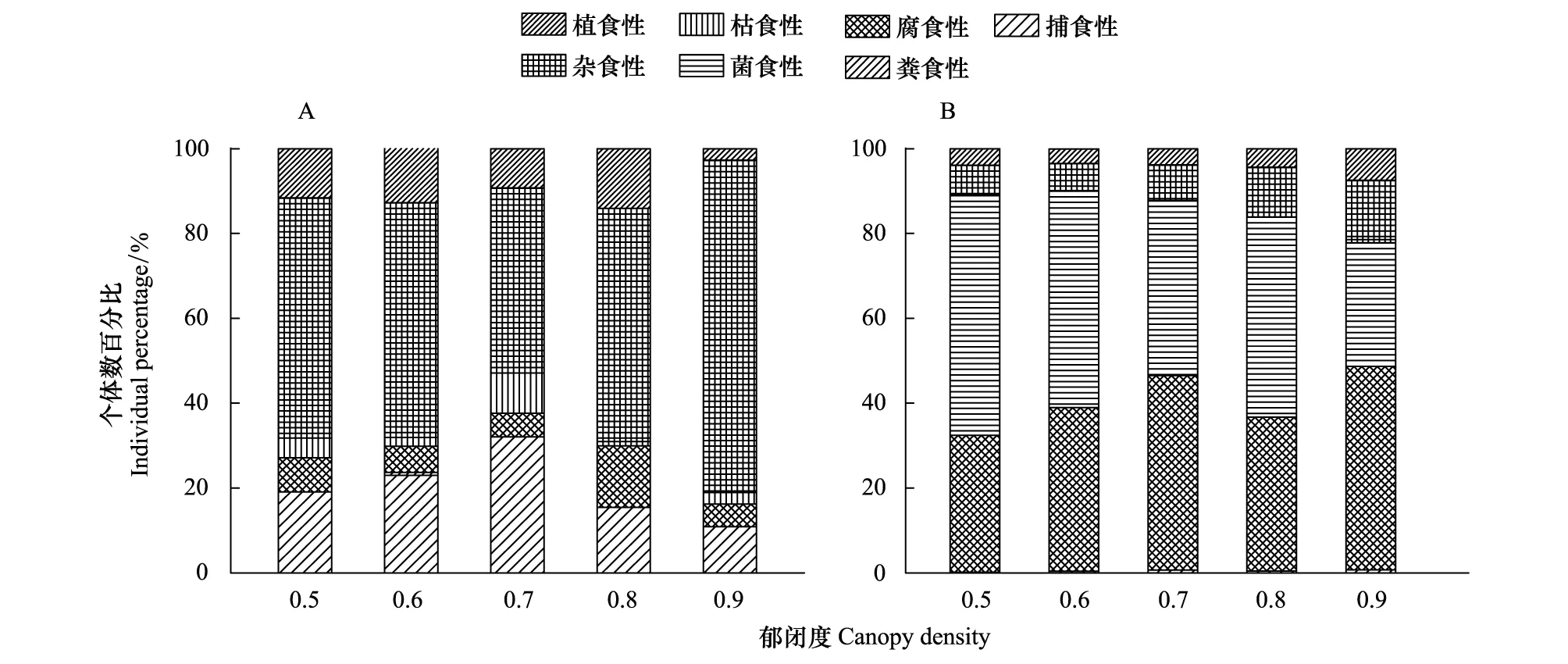
图1 不同郁闭度马尾松人工林土壤动物功能类群Fig.1 Functional group of soil fauna for different canopy density of a Pinus massoniana PlantationA:大型土壤动物各功能群个体数百分比;B:中小型土壤动物各功能群个体数百分比
3.3 土壤动物分布特征
统计分析结果显示,不同郁闭度处理对大型土壤动物个体数和类群数均无显著影响(P>0.05)。中小型土壤动物个体数随郁闭度在0.5—0.7下显著增加(P<0.05),在0.7—0.9郁闭度下显著降低(P<0.05)。中小型土壤动物类群数在0.6—0.9郁闭度无显著变化(P>0.05),在0.5郁闭度则显著降低(P<0.05)(图2)。
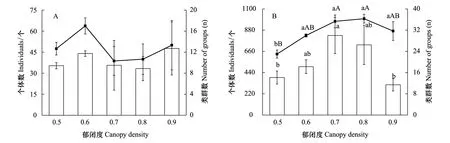
图2 不同郁闭度马尾松人工林土壤动物类群及数量(平均值±标准误)Fig.2 Soil fauna individuals and number of groups for different canopy density of a Pinus massoniana Plantation (mean±SE)A:大型土壤动物类群及数量;B:中小型土壤动物类群及数量; 不同小写字母表示显著差异(P < 0.05);不同大写字母表示极显著差异(P < 0.01)
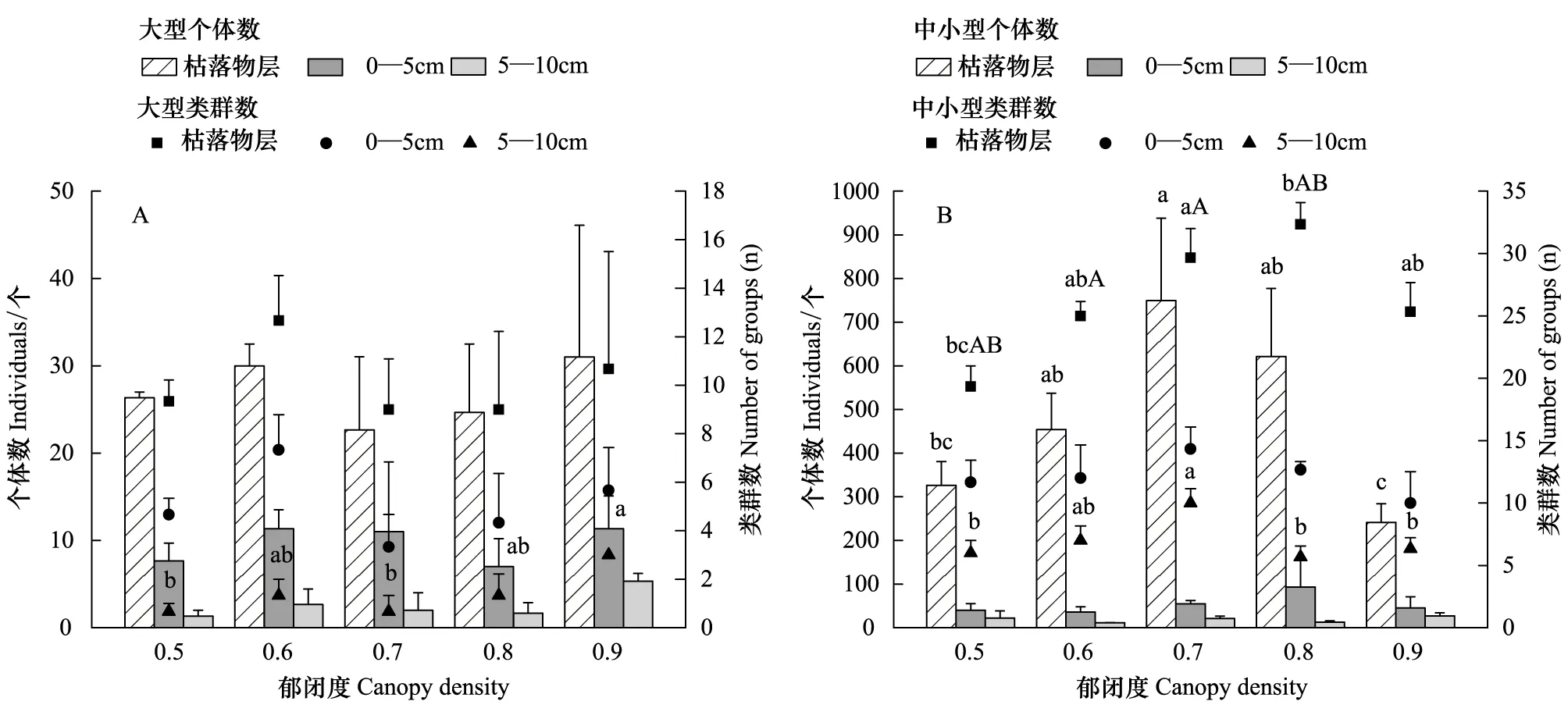
图3 不同郁闭度马尾松人工林土壤动物剖面分布(平均值±标准误)Fig.3 Profile distribution of soil fauna for different canopy density of a Pinus massoniana Plantation (mean±SE)A:大型土壤动物类群及数量剖面分布;B:中小型土壤动物类群及数量剖面分布; 不同小写字母表示显著差异(P <0.05);不同大写字母表示极显著差异(P <0.01)
郁闭度变化对不同土壤层次间大型土壤动物个体数以及枯落物层、0—5 cm层类群数没有显著影响(P>0.05),对5—10 cm层大型土壤动物类群数影响显著(P<0.05)。其中,在0.5—0.8郁闭度下大型土壤动物类群数无显著变化(P>0.05),至0.9郁闭度则显著增多(P<0.05)(图3)。枯落物层中小型土壤动物个体数和5—10 cm土层类群数在0.5—0.7郁闭度显著增高(P<0.05),0.7—0.9郁闭度显著降低(P<0.05)。枯落物层类群数在0.5—0.8郁闭度显著升高(P<0.05),且在0.7郁闭度达到极显著水平(P<0.01),0.8郁闭度下枯落物层中小型土壤动物类群数最为丰富,而0.8—0.9郁闭度则显著降低(P<0.05)。0—5 cm土层中小型土壤动物类群数随郁闭度变化不显著(P>0.05)(图3)。
3.4 土壤动物多样性与均匀性
5种郁闭度处理对大型土壤动物多样性影响不显著(P>0.05),对中小型土壤动物群落多样性产生显著影响(P<0.05)。其中,中小型土壤动物Simpson优势度指数随着郁闭度增大而显著减小(P<0.05),Shannon-Wiener多样性指数和Pielou均匀性指数则显著增大(P<0.05)。0.9郁闭度下优势度最低,而多样性指数和均匀性指数则显著高于其他郁闭度(P<0.05)。Margalef丰富度指数在0.5—0.6郁闭度显著增高(P<0.05),在0.6—0.9郁闭度则无显著变化(P>0.05)(图4)。

图4 不同郁闭度马尾松人工林土壤动物多样性特征(平均值±标准误)Fig.4 Diversity features for different canopy density of a Pinus massoniana Plantation (mean±SE)不同小写字母表示显著差异(P <0.05);不同大写字母表示极显著差异(P <0.01)
大型土壤动物多样性指数在枯落物层随郁闭度变化无显著影响(P>0.05)。但在0—5 cm层大型土壤动物 Simpson优势度和Shanon-Wiener多样性指数差异均显著(P<0.05)。5—10 cm土层Shanon-Wiener多样性指数、Pielou均匀性和Margelef丰富性差异显著(P<0.05),其中,均匀性随郁闭度增大而显著增加(P<0.05)(图5A)。中小型土壤动物多样性特征在0—5 cm均无显著影响(P>0.05)。Simpson优势度在枯落物层随郁闭度增大而显著减小(P<0.05),Shanon-Wiener多样性和Pielou均匀性显著增大(P<0.05),Margelef丰富度在0.5—0.8郁闭度下显著增大(P<0.05),0.8—0.9无显著变化(P>0.05)。在5—10 cm土壤层中,优势度在0.5—0.8无显著变化(P>0.05),0.8—0.9郁闭度时显著增大(P<0.05)。Shanon-Wiener多样性在郁闭度0.5—0.7显著增大,0.7—0.9时显著降低(P<0.05),Margelef丰富度在0.5—0.6显著增大,0.6—0.9显著降低(P<0.05)(图5B)。
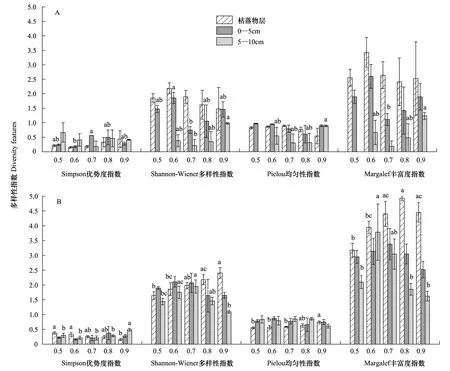
图5 不同郁闭度马尾松人工林土壤动物土壤不同层次间多样性(平均值±标准误)Fig.5 Soil fauna Diversity features for different canopy density of a Pinus massoniana Plantation (mean±SE)A:大型土壤动物多样性指数;B:中小型土壤动物多样性指数;不同小写字母表示显著差异(P <0.05);不同大写字母表示极显著差异(P <0.01)
通过计算,5个郁闭度梯度下不同体型土壤动物群落Jaccard相似性指数不同,大型土壤动物相似性指数低于中小型,在0.3—0.59之间,属于中等相似或中等不相似,并以0.7与0.8相似程度最高;中小型土壤动物群落Jaccard相似性指数在0.51—0.69之间,属中等相似,并以0.6与0.8相似程度最高(表2)。此外,在枯落物及土壤不同层次之间Jarcard相似性指数也有所差异。其中大型土壤动物除了枯落物层0.8郁闭度与0.7郁闭度、5—10 cm层0.8郁闭度与0.9、0.5郁闭度相似性指数等于0.5(中等相似)以外,在其他各层均属中等不相似或极不相似,最低相似性指数达到0.18(0—5 cm,郁闭度0.7与0.8;极不相似)。中小型土壤动物0.9郁闭度与0.5、0.6郁闭度Jaccard相似性系数在5—10 cm层极不相似,在其他各层均属中等不相似或中等相似。
3.5 土壤动物与土壤环境因子CCA分析
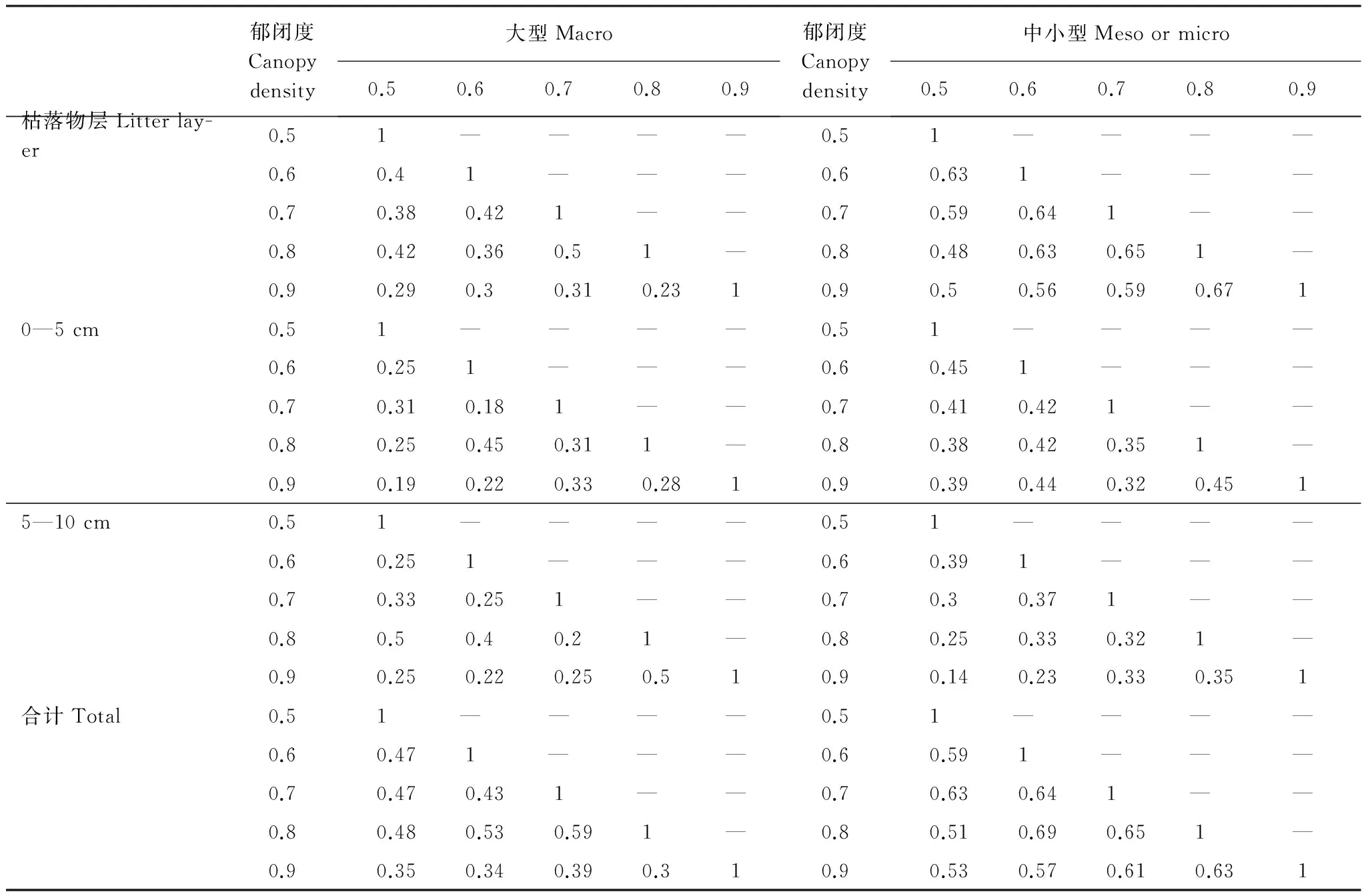
表2 不同郁闭度马尾松人工林大、中小型土壤动物Jarcard相似性系数

表3 环境变量与排序轴的相关性
*P<0.05, **P<0.01
4 讨论
在森林生态系统中,由于郁闭度不同,改变了系统的群体结构,相应地引起林内微生境的变化,包括光强、空气温度、空气相对湿度、土壤含水率、地温的日变化特和空间分布规律,以及降水的再分配等[19]。研究表明,林下微生态环境因子与林分郁闭度的相关性达到了显著水平,且高郁闭度林下极值变化范围更小[20- 21]。郁闭度的减小对应着林隙的增大和增多,改变了光照在林分中的分布,进而改变了林木各部位对光资源的利用和再分配。同时,林内光照增强使土壤温度升高,加快了土壤中有机质的分解,使土壤速效养分增加,从而提高了林地土壤肥力[22]。土壤环境因子的变化引起土壤动物类群和个体数分布差异,决定了郁闭度变化也会影响到土壤生态系统中土壤动物的群落结构组成[23- 24]。具体来讲,土壤动物个体密度与土壤有机质含量、TN、TP呈极显著正相关,与土壤含水量呈显著正相关,与土壤容重呈显著负相关[25]。本研究表明,在不同的郁闭度马尾松人工林中,土壤动物主要类群与林下土壤P含量和含水量显著相关,同时也受有机质、pH、容重和TN的共同影响。可见,郁闭度对林下微环境的改变是土壤动物群落结构发生变化的主要原因。

图6 不同郁闭度马尾松人工林土壤动物与土壤环境因子CCA分析Fig.6 CCA analysis between soil fauna groups and environmental variables for different canopy density of a Pinus massoniana Plantation CD:郁闭度 canopy density;pH:土壤pH;MC:含水量 moisture content;SOC:有机碳 soil organic carbon;TN:全氮 total nitrogen;TP:全磷 total phosphorus;Pygm:矮蒲螨;Oryi:奥地蜈蚣;Oppi:奥甲螨;Cabi:步甲;Cabo:步甲螨;Galu:大翼甲螨;Isot:等节;Blat:蜚蠊;Para:副铗叭;Phla:管蓟马;Parasi:寄螨;Campo:康叭;Noth:懒甲螨;Liac:丽甲螨;Amer:美绥螨;Asci:囊螨科;Paradox:奇马陆;Cole:鞘翅目幼虫;Lumb:大蚓;Hypogas:球角;Orib:若甲螨;Dipt:双翅目幼虫;Podu:跳虫;Salt:跳蛛;Term:螱科;Bdel:吸螨;Nema:线虫;Thom:蟹蛛;Psel:蚁甲;Form:蚁科;Stap:隐翅甲;Nean:疣;Aran:圆蛛
本研究中郁闭度显著改变了中小型土壤动物群落结构,引起类群数、个体数及多样性指数发生变化,但对大型土壤动物个体数、类群数均无显著性影响,只有5—10 cm土层类群数在0.9郁闭度表现出显著增多的趋势。这说明中小型土壤动物能够更敏感地反映林分郁闭度变化,而大型土壤动物移动力较强,营养结构更为复杂,在生境中具有相对稳定性[29],受郁闭度影响较弱。
郁闭度处理使林分内生物种间关系发生了变化。杂食性土壤动物生态位广泛,生存适应能力强,在本研究中所占比例较高。捕食性动物是土壤生物群落中的消费者,以植食性动物和腐食性动物为食,营养级别高。本研究所采集到的捕食性土壤动物个体所占的比例高于植食性,说明群落内部竞争和捕食作用使群落内各物种达到动态平衡,减少部分植食性害虫的生存和繁衍[30]。而韩宝瑜[31]等的研究表明马尾松纯林(林下植被盖度0.9)中植食性昆虫种数最多、个体数最大,天敌的种数和个体数较少。这主要是受到林下植被丰富程度的影响。随着郁闭度增大,中小型腐食性土壤动物个体数百分比显著增加,而菌食性显著减少。这是因为郁闭度越大,土壤温度降低,土壤微生物数量和活性均下降[32],导致菌食性土壤动物可食性食物源减少,故而随之减少。腐食性随郁闭度增大其比例逐渐增多,是因为高郁闭度下枯落物大量堆积[33]。而郁闭度越小,林下温度升高使枯落物很快分解,加之林下植被恢复消耗了土壤中的养分,故囊螨、维螨及其他受有机质影响的腐食性土壤动物减少。
土壤动物种类的多样性反映了其生活环境的稳定性和群落营养通道的复杂程度,是衡量群落结构水平和生态学特征的重要量度指标[34]。多样性越高,代表采样区生态结构越稳定,则有利于土壤动物的生存与繁衍[35]。在本次实验中,郁闭度变化对大型土壤动物在各土壤层次中的多样性影响显著,且均匀性逐渐增强。随着郁闭度增大,中小型土壤动物群落的Simpson优势度指数逐渐减小,Shannon-Wiener多样性指数和Pielou均匀性指数则显著增大。可能是因为林冠荫蔽作用使林下环境条件得到改善,能够适宜更多类群的土壤动物生存,因此多样性指数也逐渐增大,均匀性增强。Margalef指数也表现在高郁闭度下更加丰富。然而,郁闭度过大会限制林下植被恢复,植食性土壤动物所占比例太小,使整个林分的营养结构单一,不利于生态系统的长期稳定。
5 结论
郁闭度变化引起林下环境条件相应改变,对大型土壤动物群落结构影响较小,主要对中小型土壤动物类群及数量、多样性、功能类群等在各郁闭度下及不同土壤层次之间影响显著,郁闭度过高或过低都不利于保持土壤动物群落多样性和稳定性。0.7郁闭度马尾松人工林下土壤动物优势度、丰富度都保持在适中水平,虽然均匀性和多样性还不太理想,但随着林下植被恢复的时间尺度增加,该郁闭度马尾松人工林可能更有利于土壤动物结构和功能的发挥。但是,本研究仅是郁闭度控制后马尾松人工林土壤动物群落的初期调查,研究结果反应的仅是一种趋势,这种趋势是否会随着郁闭度控制时间的推移而发生改变还有待进一步研究。
[1] Brussaard L, Pulleman M M, Ouédraogo É, Mando A, Six J. Soil Fauna and Soil Function in the Fabric of the Food Web. Pedobiologia, 2007, 50(6): 447- 562.
[2] 武海涛, 吕宪国, 杨青, 姜明. 土壤动物主要生态特征与生态功能研究进展. 土壤学报, 2006, 43(2): 314- 323.
[3] Peng Y, Yang W Q, Li J, Wang B, Zhang C, Yue K, Wu F Z. Contribution of Soil Fauna to Foliar Litter-Mass Loss in Winter in an Ecotone between Dry Valley and Montane Forest in the Upper Reaches of the Minjiang River. Plos one, 2015, 10(4): e0124605.
[4] Huang W, Siemann E, Xiao L, Yang X F, Ding J Q. Species-Specific Defence Responses Facilitate Conspecifics and Inhibit Heterospecifics in Above-Belowground Herbivore Interactions. Nature Communications, 2014, 5: 4851 doi:10.1038/ncomms5851.
[5] Zaitsev A S, Chauvat M, Wolters V. Spruce Forest Conversion to a Mixed Beech-Coniferous Stand Modifies Oribatid Community Structure. Applied Soil Ecology, 2014, 76(2): 60- 67.
[6] 张良辉. 川西亚高山云杉人工林分结构调控下植物与土壤动物多样性研究[D]. 雅安: 四川农业大学, 2010.
[7] 刘茂秀, 史军辉, 王新英, 彭秋梅. 林分郁闭度和龄级对防护林碳汇功能及固碳价值的影响. 东北林业大学学报, 2011, 39(6): 30- 32.
[8] 国家林业局. 中国森林资源简况--第八次全国森林资源清查, 2013[2015]. http://www.forestry.gov.cn/gjslzyqc.html
[9] 洪伟, 吴承祯. 马尾松人工林经营模式及其应用. 北京:中国林业出版社, 1999.
[10] 杨平平, 徐仁扣, 黎星辉. 淋溶条件下马尾松针对土壤的酸化作用. 生态环境学报, 2012, 21(11): 1817- 1821.
[11] 杨承栋, 孙启武, 焦如珍, 卢立华. 大青山一二代马尾松土壤性质变化与地力衰退关系的研究. 土壤学报, 2003, 40(2): 267- 273.
[12] 李永宁, 张宾兰, 秦淑英, 李帅英, 黄选瑞. 郁闭度及其测定方法研究与应用. 世界林业研究, 2008, 21(1): 40- 56.
[13] 周彬, 韩海荣, 康峰峰, 程小琴, 宋娅丽, 刘可, 李勇. 太岳山不同郁闭度油松人工林降水分配特征. 生态学报, 2013, 33(5): 1645- 1653.
[14] 王邵军, 阮宏华, 汪家社, 徐自坤, 吴焰玉. 武夷山典型植被类型土壤动物群落的结构特征. 生态学报, 2010, 30(19): 5174- 5184.
[15] 尹文英. 中国亚热带土壤动物. 北京: 科学出版社, 1992.
[16] Swift M J, Heal O W, Anderson J M. Decomposition in Terrestrial Ecosystems. Berkeley and Los Angeles: University of California Press, 1979.
[17] 尹文英. 中国土壤动物. 北京: 科学出版社, 2000.
[18] 林英华, 贾旭东, 徐演鹏, 李慧仁, 刘学爽, 徐永波, 韦昌雷, 刘三章, 王立中. 大兴安岭典型森林沼泽类型地表土壤动物群落与生态位分析. 林业科学, 2015, 51(12): 53- 62.
[19] 宋小帅, 康峰峰, 韩海荣, 于晓文, 周彬, 程小琴. 太岳山不同郁闭度油松人工林枯落物及土壤水文效应. 水土保持通报, 2014, 34(3): 102- 108.
[20] 康文星, 邓湘雯, 赵仲辉. 林冠截留对杉木人工林生态系统物质循环的影响. 林业科学, 2006, 42(12): 1- 5.
[21] 成向荣, 冯利, 虞木奎, 吴统贵, 孙海菁, 王宗星, 张翠. 间伐对生态公益林冠层结构及土壤养分的影响. 生态环境学报, 2010, 19(2): 355- 359.
[22] 李岩. 阔叶红松林林隙小气候及土壤物理性质特征比较研究[D]. 哈尔滨: 东北林业大学, 2007.
[23] Taki H, Inoue T, Tanaka H, Makihara H, Sueyoshi M, Isono M, Okabe K. Responses of Community Structure, Diversity, and Abundance of Understory Plants and Insect Assemblages to Thinning in Plantations. Forest Ecology and Management, 2010, 259(3): 607- 613.
[24] 刘任涛, 赵哈林, 赵学勇. 科尔沁沙地不同造林类型对土壤动物多样性的影响. 应用生态学报, 2012, 23(4): 1104- 1110.
[25] 武海涛, 吕宪国, 杨青, 姜明, 佟守正. 三江平原湿地岛状林土壤动物群落结构特征及影响因素. 北京林业大学学报, 2008, 30(2): 50- 58.
[26] Ponge J F. Biocenoses of Collembola in Atlantic Temperate Grass-woodland Ecosystems. Pedobiologia, 1993, 37(4): 223- 244.
[27] Nickerl J, Helbig R, Schulz H J, Werner C, Neinhuis C. Diversity and Potential Correlations to the Function of Collembola Cuticle Structures. Zoomorphology, 2013, 132(2): 183- 195.
[28] 张瑞益. 北方主要草地类型土壤线虫群落结构及多样性的比较研究[D]. 长春: 东北师范大学, 2015.
[29] 黄丽荣, 张雪萍. 大兴安岭寒温带地区中小型土壤动物群落特征. 应用与环境生物学报, 2008, 14(3): 388- 393.
[30] 朱新玉, 高宝嘉, 胡云川. 森林草原交错带土壤节肢动物同功能种团组成及分布. 江苏农业科学, 2011, 39(3): 478- 581.
[31] 韩宝瑜. 马尾松林节肢动物群落的组成及多样性. 生物多样性, 2001, 9(1): 62- 67.
[32] 张鼎华, 叶章发, 范必有, 危廷林. 抚育间伐对人工林土壤肥力的影响. 应用生态学报, 2001, 12(5): 672- 676.
[33] 张雪萍, 侯威岭, 陈鹏. 东北森林土壤动物同功能种团及其生态分布. 应用与环境生物学报, 2001, 7(4): 370- 374.
[34] 谭波, 吴福忠, 杨万勤, 张健, 徐振锋, 刘洋, 苟小林. 不同林龄马尾松人工林土壤节肢动物群落结构. 应用生态学报, 2013, 24(4): 1118- 1124.
[35] 王振中, 张友梅, 李忠武. 黄山森林生态系统土壤动物群落结构特征及其多样性. 林业科学, 2009, 45(10): 168- 173.
Community characteristics of soil fauna for different canopy density of aPinusmassonianaplantation
ZHOU Hongyang1, ZHANG Jian1, 2,*, ZHANG Danju1, ZHANG Jie1, WEI Daping1, ZHAO Yanbo1, ZHAO Bo2,LI Chuanbei2
1SichuanProvinceKeyLaboratoryofEcologicalForestryEngineering,InstituteofEcology&Forestry,SichuanAgriculturalUniversity,Chengdu611130,China2ForestryCollegeofSichuanAgriculturalUniversity,Chengdu611130,China
Soil fauna is critical to the nutrient cycling, litter decomposition promoting and soil structure formation. The relationship between underground biota especially soil fauna and aboveground vegetation are beneficial to the forest ecosystem stability. Therefore, we studied the community structure of soil fauna inPinusmassonianaplantation controlled by different sized canopy density (proportion of the forest floor covered by the tree crowns′ vertical projection: 0.5, 0.6, 0.7, 0.8, 0.9). In October 2014, we used the hand picking up method, Tullgren funnel method and Baermann technique to collect macro, meso or micro soil fauna in the upper Yangtze River at Yibin. The results revealed that a total of 8860 individuals soil fauna were collected, belonging to 4 phyla, 11 classes,25 orders and 111 families. These include 588 macro individuals with Blattidae, Neanuridae and Campodeidae as the dominant species and with omnivorus as the main feeding habit. Besides, the rest of 8271 individuals were meso or micro fauna with Isotomidae and Liacaridae as the dominant species, the feeding habits were mainly fungivorous forms and saprozoic. Meanwhile, the percentage of macro omnivorous soil fauna increased significantly in 0.9 canopy density and predators decreased conversely. Soil fauna of meso or micro sized predators increased significantly in 0.5—0.7 canopy density and then had no significant change along with canopy density. The proportion of meso or micro saprozoic soil fauna increased significantly with canopy density increasing but opposite trend we found for fungivorous forms. In addition, canopy density treatments only impacted on the number of macro soil fauna groups, which significant increase in 0.9. With increasing canopy density, all of the total individuals, little layer individuals and 5—10 cm layer groups number of soil meso or micro fauna increased (0.5—0.7) and subsequently decreased (0.7—0.9) significantly. The total group number of meso or micro soil fauna increased in 0.5—0.6 canopy density. In litter layer the group number of meso or micro soil fauna also increased in 0.5—0.8 significantly and 0.8—0.9 decreased. We had also take diversity feature into consideration. The diversity value of macro, meso or micro soil fauna were significantly different with soil layer. Following the canopy density increased, the value of Simpson index of meso or micro soil fauna decreased although the Shannon-Wiener index and Pielou index were significantly increased. Simultaneously the Margalef abundance index were significantly higher firstly (0.5—0.6) and decreased (0.6—0.9) subsequently. Furthermore, macro soil fauna Jaccard similarity coefficient is lower than the meso or micro sized. Finally, the CCA (canonical correlation analysis) results showed that different response to environmental factors for different groups of soil fauna. Canopy density, moisture content, organic matter and total phosphorus have significant effects on these groups. So based on our research, we proposed thatPinusmassonianaplantation with 0.7 canopy density has relatively abundant soil fauna diversity and stable community.
Pinusmassonianaplantation; canopy density; soil fauna; community structure

附表1 不同郁闭度下马尾松人工林大型土壤动物群落组成
—表示无或未采集到,+++为优势类群,++为常见类群,+为稀有类群;Ph:植食性 Phytophage; F:菌食性 Fungivorous forms;Pr:捕食性 Predators; S:腐食性 Saprozoic;O:杂食性 Omnivores;D:枯食性 Debris-feeder′ s;Co:粪食性 Coprophages
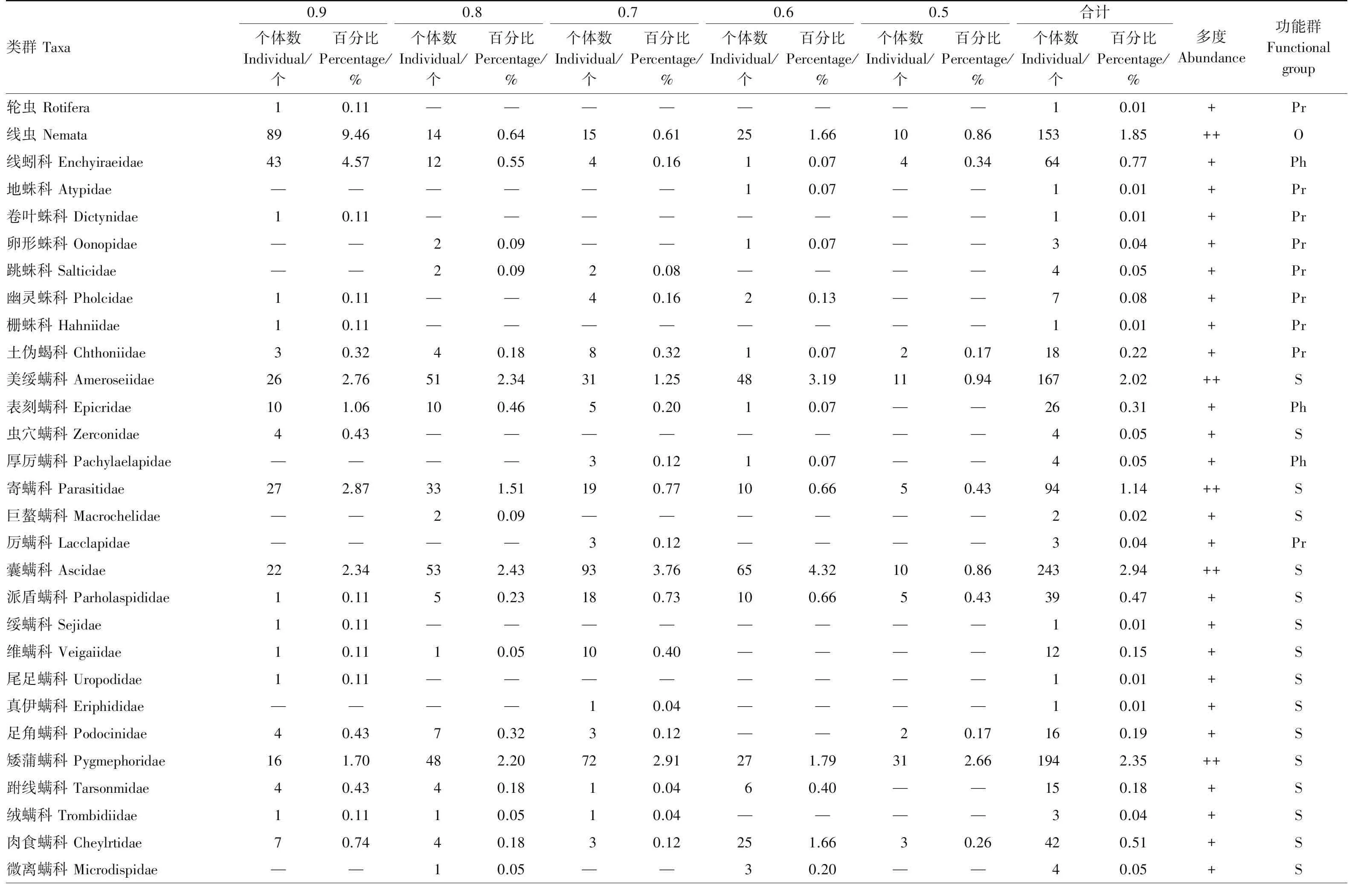
附表2 不同郁闭度下马尾松人工林大型土壤动物群落组成
—表示无或未采集到,+++为优势类群,++为常见类群,+为稀有类群; Ph:植食性 Phytophage; F:菌食性 Fungivorous forms;Pr:捕食性 Predators; S:腐食性 Saprozoic;O:杂食性 Omnivores;D:枯食性 Debris-feeder′ s;Co:粪食性 Coprophages
国家自然科学基金项目 (31370628);国家科技支撑计划项目(2011BAC09B05);四川省科技支撑计划项目(12ZC0017);四川省科技厅应用基础项目(2012JY0047);四川省教育厅科技创新团队计划项目(11TD006)
2015- 10- 23;
日期:2016- 08- 02
10.5846/stxb201510232148
*通讯作者Corresponding author.E-mail: sicauzhangjian@163.com
周泓杨,张健,张丹桔,张捷,魏大平,赵燕波,赵波,李川北.不同郁闭度控制下马尾松(Pinusmassoniana)人工林土壤动物群落特征.生态学报,2017,37(6):1939- 1955.
Zhou H Y, Zhang J, Zhang D J, Zhang J, Wei D P, Zhao Y B, Zhao B,Li C B.Community characteristics of soil fauna for different canopy density of aPinusmassonianaplantation.Acta Ecologica Sinica,2017,37(6):1939- 1955.
