马尾松木材径向与弦向抗弯性能及破坏特征的比较研究∗
2017-04-27张娅梅
张娅梅 潘 彪 王 丰
木材是典型的非均质材料,与一般钢材、混凝土及石材等材料不同,木材属生物材料,其构造的各向异性导致其力学性能的各向异性。蔡则漠的早晚材缓变树种杉木16个样本的标准力学试验数据结果表明,除横纹抗压强度外,弦向的抗弯强度、弹性模量、冲击韧性、顺纹抗剪强度及抗劈力均不同程度地高于径向[1]。近年来许多学者开始将木材的解剖特征与其力学性能联系起来,提出木材微观构造对木材力学性能的影响是造成这种差异的主要原因之一[2]。深入研究木材的破坏机理、木材构造与其力学性能相互关系,对克服木材缺点具有重要的指导意义[3]。研究木材的断裂过程是认识木材破坏机理的一个主要方面,任宁等[4]采取了连续加载的方式,实时追踪监测木材断裂过程,探讨了木材微观构造与拉伸断裂的关系。木材的破坏过程实际上就是木材中原有微裂纹的萌生、扩展、贯通直到最后宏观裂纹产生并导致木材细胞壁破裂[5]。由于裂纹以声速在木材中扩展[6],直接监测裂纹在木材中的扩展较为困难,裂纹以不同方式扩展,必然会在破坏面上留下不同形态。因此,利用扫描电镜分析破坏面上裂纹扩展时留下的残痕,对破坏面进行观察是研究木材破坏过程的一种有效途径[7]。
F.Thuvander等利用光学显微镜观察到樟子松横切面的拉伸断裂主要源于细胞壁中间层的撕裂或剥离[8]。S.W.J. BOATRIGHT用扫描电子显微镜(SEM)观察南非松破坏面,研究表明试件中存在不可逆的细胞变形,包括脱粘,轴向扭曲和屈曲[9]。
细胞是构成木材微观的基本形态单位,针叶材的管胞、木射线、细胞壁上的纹孔以及早晚材结构的差异都可能对木材破坏有很大的影响,因此破坏面微观的观察分析是研究木材破坏必不可少的部分。笔者对比早晚材急变的马尾松木材在弯曲荷载作用下抗弯强度、抗弯弹性模量及破坏形态的差异,希望为探究木材微观构造对弯曲破坏的影响提供一些理论依据。
1 试验材料与方法
1.1 材料
马尾松试材按国家标准GB/T 1927—1991《木材物理力学试材采集方法》于1994年12月采自贵州龙里林场的标准试验林(位于东经106°58′,北纬26°27′,海拔1 150 m,坡向东南,坡度5°~10°)。树龄30年,取基部向上1.3~2.3 m木段的气干板材用于试件的加工,根据GB/T 1936-1—2009《木材抗弯强度试验方法》加工试件,尺寸为300 mm×20 mm×20 mm,长度为顺纹方向。
1.2 试验方法
试验前挑选出年轮规整、表面为标准三切面的马尾松木材试件,用于径、弦向抗弯试验的试件各50个备用。设备采用最大试验力为30 kN的UTM4304三思纵横万能力学试验机,径、弦向抗弯试验按GB/T 1936-1—2009测定。在抗弯试验后测定所有马尾松试件的含水率与气干密度,抗弯强度与抗弯弹性模量均换算成含水率为12%时的值。
试验后将试件在XZX7体视显微镜拍下破坏形态。然后在破坏部位分别截取剪切破坏处和拉伸断裂处小块样品,破坏部位朝上,用导电胶粘在样品台上,接着用离子喷金仪进行喷金处理,最后用日立TM-1000台式扫描电子显微镜对样品进行观察,并调整到适合的倍数拍照。将体视显微镜与扫描电子显微镜下拍到的照片分类整理,观察并分析径、弦向受力时破坏处细胞被破坏的差异。
2 结果与分析
2.1 径、弦向抗弯强度及抗弯弹性模量差异分析
试验测得所有试件的平均气干密度为0.547g/cm3。由于密度与抗弯强度呈现显著的相关性,密度对抗弯性能有很大的影响,为减小两组试件密度相差过大对试验结果的影响,从备用试件中选择密度在0.50~0.62 g/cm3范围内的试件用于比较径、弦向的抗弯强度及抗弯弹性模量的差异,用于径、弦向抗弯试验的试件数分别为47个和36个。
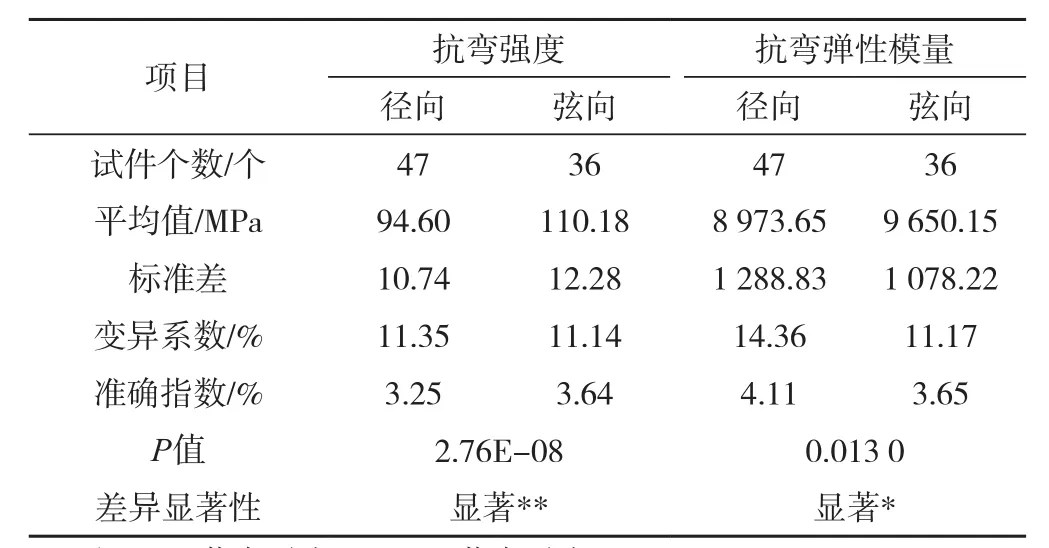
表1 马尾松木材抗弯强度及抗弯弹性模量的变异统计及差异分析Tab.1 Variance statistics and variation analysis of MOR and MOE of masson pine
试验结果如表1所示:马尾松弦向平均抗弯强度大于径向平均抗弯强度,在显著水平为0.01下即呈现显著差异;弦向平均抗弯弹性模量大于径向平均抗弯弹性模量,在显著水平为0.05下即呈现显著差异。径、弦向受力对抗弯强度的影响大于抗弯弹性模量,这与节子[10]、含水率[11]、密度[12]等影响因子对抗弯强度的影响大于抗弯弹性模量的结论一致。主要原因是:
1)径向受力时,剪切层或为早材,或为晚材。在某些位置只有早材受到拉伸和剪切应力,由于早材细胞壁较薄,早晚材边界处组织构造差异大,早材先于晚材破坏,裂纹扩展到早晚材边缘时,会沿着边界处的早材扩展,因此剪切破坏通常发生在相邻年轮早晚材边界处,抗弯强度、抗弯弹性模量相对较小。
2)弦向受力时,试件底部早、晚材同时受到拉伸作用力,剪切层由早、晚材间隔组成,晚材致密,不容易破坏,因此木材的抗弯强度、抗弯弹性模量较大。宏观可以看到剪切破坏处有大量的木射线,说明试件受到剪切应力后沿木射线开裂。
径向抗弯强度及抗弯弹性模量的离散程度均大于弦向,如表1。若径向受力试件的密度平均值、变异系数与弦向相等,可以排除密度的影响。但是所选择径、弦向受力试件的密度变异系数不同且径向小于弦向,如表2,若弦向受力试件的密度的变异系数减小至与径向相等,相应的弦向抗弯强度及抗弯弹性模量的变异系数会随之减小,故实际径、弦向抗弯强度及抗弯弹性模量的变异系数差值应比表1所示的数值大,离散程度相差得也就更大。
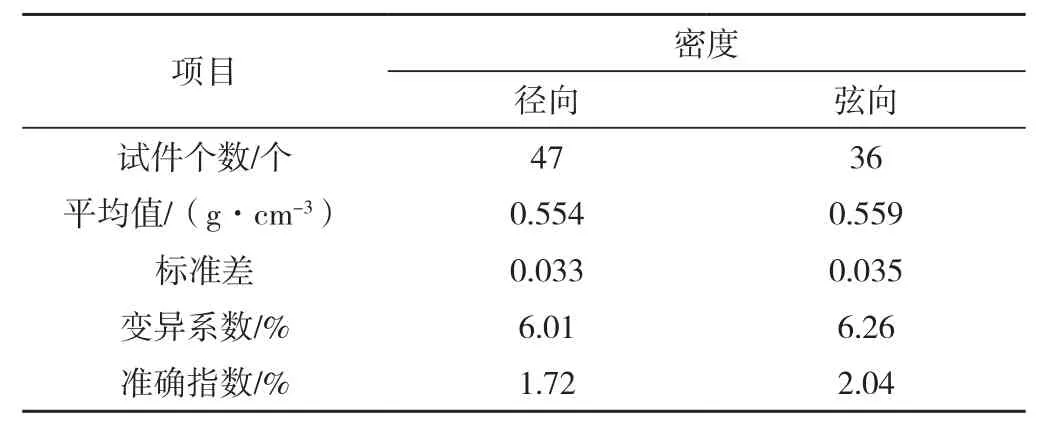
表2 马尾松木材径、弦向受力弯曲试件的密度变异统计Tab.2 Variance statistics of density of the specimen under radial and tangential bending load
2.2 木材弯曲破坏面的微观分析
发生的木材弯曲破坏往往在拉伸区和剪切区明显,受压区破坏不明显,故选取拉伸区和剪切区的破坏面进行观察。在试件径、弦向受力时均观察其受到拉伸应力而断裂的面,即与荷载方向平行的横切面。径向受力时观察试件受到剪切应力而破坏的面,即与荷载方向垂直的弦切面。而弦向受力时观察试件受到剪切作用力而破坏的面,即与荷载方向垂直的径切面。
2.2.1 拉伸断裂面的显微观察
1)在弯曲荷载作用下,试件与荷载方向平行的横截面任意点的拉伸应力与该点至中性面的距离成正比,即沿截面高度按直线规律变化,因此截面上的同一管胞不同高度受到拉伸应力的作用大小不同;管胞的径、弦面壁有较明显的差异,管胞径面壁上纹孔的数量多于弦面壁,径面壁木质素含量高于弦面壁,导致了管胞径、弦面壁的性能差异,因此径、弦面壁在受到拉伸作用力时的抗拉能力不同,在拉伸应力作用下拉伸断裂面表层管胞会出现扭转现象。
2)弦向受力时,剪切层由早晚材共同组成,破坏强度较大,即试件的破坏主要是由于拉伸应力的作用,如图1a,反应在微观上为单个细胞的拉断。因此弦向受力时试件受到拉伸应力产生的断裂面表层管胞扭转现象明显,管胞的扭转导致管胞壁沿着纤丝角方向产生了裂隙,如图2a。且拉伸断裂面比较粗糙。从横断面观察如图2b。从顺纹的方向能更明显地观察到管胞扭转现象,如图2c。

图1 径、弦向受力时破坏形态Fig.1 Failure surface under radial and tangential bending load

图2 弦向受力时拉伸断裂面细胞形态Fig.2 Cell morphology of tensile fracture surfaces under tangential bending load
3)径向受力时,剪切层与年轮平行,裂纹通常沿着早晚材边界处扩展,即破坏主要是由于剪切应力的作用,如图1b,微观上表现为细胞间滑移或撕裂分离。因此径向受力时试件因拉伸应力产生的断裂面与弦向受力时相比,管胞无明显的扭转现象,且没有产生裂隙,如图3a。拉伸断裂面比较粗糙,这与弦向受力时的情况是一致的, 从横断面观察如图3b,从顺纹的方向观察如图3c。
2.2.2 剪切破坏面的显微观察

图3 径向受力时拉伸断裂面细胞形态Fig.3 Cell morphology of tensile fracture surfaces under radial bending load
1)弦向受力时,试件受到顺纹方向的剪力而破坏的面在径切面上,破坏发生在木射线细胞处,主要原因是射线薄壁细胞较为薄弱,如图4a。剪切应力使细胞之间产生滑移,观察剪切破坏面上的管胞,如图4b所示,管胞壁上微纤丝呈平行排列且与细胞轴呈50°~70°角,可见管胞内壁上的纹孔,判断是次生壁外层S1层附着在管胞外壁上。故认为剪切破坏通常是在复合胞间层处分离,即在管胞间产生破坏。但由于初生壁通常只有细胞壁厚度的1%,复合胞间层很薄,分离后有部分S1层附着在管胞壁上。细胞壁被破坏后,依旧能看到完整纹孔附着在相邻管胞的细胞壁,如图4c。纹孔虽然是木材中比较薄弱的部分,却能抵抗裂纹的扩展,主要是由于微纤丝的排列方向在纹孔周围发生改变,微纤丝绕过纹孔口而排列,与裂纹扩展方向垂直。在马尾松试件断裂时,裂纹会沿着纹孔的边缘扩展,因此纹孔仍能保持完整形态。

图4 弦向受力时剪切破坏面细胞形态Fig.4 Cell morphology of shear breaking surfaces under tangential bending load
2)径向受力时,试件受到顺纹方向的剪力而破坏的面在弦切面上,观察管胞的结果同弦向受力的情况一致,剪切破坏是管胞间的分离,且裂纹会沿着纹孔的边缘扩展,纹孔仍能保持完整形态,如图5。

图5 径向受力时剪切破坏面细胞形态Fig.5 Cell morphology of Shear breaking surfaces under radial bending load
3 结论
对比径、弦向弯曲荷载作用下马尾松木材抗弯强度、抗弯弹性模量,使用扫描电子显微镜观察径、弦向破坏面,结果表明:
1)马尾松弦向抗弯强度、抗弯弹性模量均大于径向,差异显著。晚材的影响使弦向受力时破坏强度较大,剪切破坏则是沿木射线开裂。径向受力时,试件剪切破坏通常发生在早晚材边界处,破坏强度相对较小。受力方向对抗弯强度的影响大于抗弯弹性模量。径向抗弯强度及抗弯弹性模量的离散程度均大于弦向,由于径、弦向受力试件的密度变异系数不同的影响,实际径、弦向抗弯强度及抗弯弹性模量的离散程度应相差更大。
2)径、弦向受力时拉伸断裂面均比较粗糙,剪切破坏通常发生在管胞间。弯曲断裂时裂纹会沿着纹孔的边缘扩展,纹孔仍能保持完整形态。
3)弦向受力时,拉伸断裂面表层管胞扭转现象明显,并产生裂隙;剪切破坏为沿比较薄弱的木射线开裂。径向受力时,拉伸断裂面表层管胞无明显扭转现象;剪切破坏发生在相邻年轮的早晚材边界处。
[1]蔡则漠. 杉木径面及弦面的木材力学性质的差异[J]. 林业科学,1996, 32(3):254-259.
[2]任宁. 木材微观构造对受载断裂的影响方式研究[D]. 哈尔滨:东北林业大学, 2007.
[3]邵卓平. 木材损伤断裂与木材细观损伤基本构元[J]. 林业科学,2007, 43(4):107-110.
[4]任宁, 刘一星, 巩翠芝, 等. 木材微观构造与拉伸断裂的关系[J]. 东北林业大学学报, 2008, 36(2):33-35.
[5]李大纲.木材细胞壁细观断裂及其损伤机理[J]. 科学技术与工程,2004, 4(1):24-27.
[6]任海青, 江泽慧. 人工林杉木、马尾松木材的断裂特性[J]. 林业科学,2001, 37(3):118-123.
[7]江泽慧, 任海青, 胡一贯, 等. 木材断裂过程的研究[J]. 核技术, 2000,23(8):572-576.
[8]THUVANDER F, BERGLUND L A. In situ observations of fracture mechanisms for radial cracks in wood[J]. Journal of Materials Science,2000, 35(24):6277-6283.
[9]BOATRIGHT S W J , GARRETT G G. The effect of microstructure and stress state on the fracture behaviour of wood[J]. Journal of Materials Science,1993,18(7):2181-2199.
[10]阙泽利. SPF规格材密度与节子变异特性对抗弯性能的影响[J]. 林产工业, 2016, 43(3):16-18.
[11]成俊卿. 木材学[M]. 北京:中国林业出版社, 1985:65-66.
[12]骆秀琴, 管宁, 张寿槐, 等. 杉木材性株内变异的研究[J]. 林业科学,1997, 33(4):349-355.
