芦苇根际土壤可培养真菌多样性的季节变化
2017-04-17李燕超
姜 华, 李燕超, 姜 一
(1.辽宁师范大学 生命科学学院,辽宁 大连 116081; 2.天津出入境检验检疫局 动植物与食品检测中心,天津 300456)
芦苇根际土壤可培养真菌多样性的季节变化
姜 华1, 李燕超1, 姜 一2
(1.辽宁师范大学 生命科学学院,辽宁 大连 116081; 2.天津出入境检验检疫局 动植物与食品检测中心,天津 300456)
以稀释平板法和形态学、分子生物学鉴定法对芦苇(Phragmitesaustralis)进行了根际土壤可培养真菌的分离鉴定及季节变化对其种群结构影响的分析.结果表明:春、夏、秋、冬四季芦苇根际土壤可培养真菌菌落总数分别是5 243、5 003、6 388和4 970,种类数分别为9属21种、8属18种、11属22种和6属21种;真菌的多样性指数、丰富度指数值,从大到小的次序均为秋>春>夏>冬,而均匀度指数为春>夏>冬>秋,多样性指数和丰富度指数在秋季表现最高,但数值波动较大,均匀度指数差别较小.芦苇根际土壤中的真菌资源丰富,共分离出15 属54 种真菌,优势属为曲霉属(Aspergillus)、青霉属(Penicillium)、拟青霉属(Paecilomyces)、木霉属(Trichoderma)和链格孢属(Alternaria);金孢属(Chrysosporium)、伏革菌属(Corticium)、粘帚霉属(Gliocladium)、踝节菌属(Talaromyces)和根霉属(Rhizopus)是无植被土壤中没有的菌属.芦苇对土壤真菌多样性具有较强的保护作用,其根际土壤真菌的种类是无植被土壤的1.2倍,数量是无植被土壤的4倍.
芦苇;根际土壤;真菌多样性
芦苇(Phragmitesaustralis)属于禾本科(Gramineae)芦苇属(PhragmitesAdans)多年生草本植物,主要生长于温带和热带地区,世界各地分布广泛,我国主要分布在东北、内蒙古、新疆和华北等地.芦苇是造纸及建材原料,被誉为“第二森林”,具有较高的经济价值;根可入药,有解毒、利尿、清凉、镇呕等药用功能;也具有重要的生态价值,能涵养水源、调节气候,保护湿地生态环境,抗逆性强、生态幅广,可在湖滨、池塘、沼泽、河岸、滩涂、河口等浅水湿地形成密集的单优群落,甚至在荒漠、盐碱地也广泛分布[1-3].土壤微生物影响着矿物质营养的吸收与释放、土壤有机质的分解、碳、氮循环和成土过程[4].丰富多样的微生物群落是保持土壤生态系统稳定的重要因素,其数量与分布在一定程度上反映了土壤肥力状况.因此,研究和保护土壤微生物多样性对保护生态系统具有重要意义.鉴于芦苇的生态及经济价值,前人对此曾进行了芦苇耐盐基因的研究[5]、芦苇对重金属污染土壤的生态修复功能研究[6],以及芦苇病虫害防治、芦苇资源及分布与生长状况调查、芦苇对环境保护及污水处理、芦苇的生理及生态结构、芦苇资源的开发利用、芦苇加工设备等研究[7],但有关芦苇微生物等方面的研究报道较少.本文旨为查明芦苇根际土壤可培养真菌的种群构成,以及它们对季节变化的响应,为芦苇资源养护及污染土壤的芦苇-微生物修复提供理论依据及优势菌株.
1 材料与方法
1.1 材料
1.1.1 试材 芦苇(P. australis)采自辽宁省大连市甘井子区营城子镇盐场.
1.1.2 培养基与试剂 采用虎红固体培养基进行真菌分离,马铃薯培养基(PDA)进行真菌纯化;DNA Marker(DL2000)、dNTP、Taq DNA 聚合酶、buffer等均购自大连TaKaRa 公司;引物合成、DNA测序由上海生工生物工程公司完成;所需试剂均为国产分析纯.
1.2 方法
1.2.1 土样的采集 采样地位于辽宁省大连市甘井子区营城子镇盐场.该区除了大面积的芦苇外,还有辽宁碱蓬等植物.分别于2015年的春(5 月9日)、夏(7 月9日)、秋(9月8日)、冬(12月14日)四季,采用五点采样法,采集深10 cm左右的根系上的土壤(以无植被土壤为对照)[8],3次重复.
1.2.2 真菌的分离纯化与计数 土样进行10倍浓度梯度稀释,以涂布平板法接种土样稀释液.用移液枪吸取200 μL的10-4~10-2土壤稀释液于虎红(孟加拉红)平板上,涂布,3次重复.培养3~5 d 后进行真菌菌落计数[8],再以PDA培养基进行真菌的纯化.
1.2.3 真菌鉴定
(1)形态学鉴定:依据菌落和个体形态特征,参照真菌分类鉴定指标进行[9].
(2)分子生物学鉴定:对于难以产生分生孢子、以形态学特征难以确认的可疑菌株进行分子鉴定[10].提取菌株DNA,琼脂糖凝胶电泳检测DNA纯度;使用真菌通用引物ITS1和ITS4,进行菌株rDNA-ITS区的PCR扩增;PCR 纯化产物委托生物公司进行DNA序列测定,测序结果在 NCBI网站上进行BLAST序列比对和同源性分析,构建分子系统发育树,确定菌株种类.
1.2.4 统计分析[8]
(1)分离频率
分离频率(%)= 某一真菌分离物的总数(株)/所有分离物的总数(株)×100%.
(2)真菌种群多样性:采用Shannon多样性指数(H′)、Pielou均匀度(J)指数和Margalef丰富度指数(R)来测定.公式为
H′=-∑PilnPii=1,2…n;J=H′/ln(S);R=(S-1)/lnN.
式中:Pi为第i种的个体数占总个体数的比例,可用Pi=Ni/N求出;Ni为第i种物体个体数,N为总个体数,S为每个样品的物种总数.
(3)Jaccard相似性指数:是用来比较2个样本之间真菌种类组成的相似程度,公式为:Cj=c/(a+b-c).当Cj为0.00~0.25时,为极不相似;当Cj为0.25~0.50时,为中等不相似;当Cj为0.50~0.75时,为中等相似;当Cj为0.75~1.00时,为极相似.
2 结果及分析
2.1 芦苇根际土壤可培养真菌种群构成对季节的响应
于春、夏、秋、冬4个季节采样,检测芦苇根际土壤可培养真菌菌落总数,其结果分别为5 243、5 003、6 388和4 970;真菌种类数分别为9属21种、8属18种、11属22种和6属21种;共分离鉴定出15属54种真菌.4个季节的芦苇根际土壤中,分离出的真菌种类数量依次是秋季>春季>冬季>夏季,秋季种类数量最多、夏季最少,且四季的优势菌属也略有不同.春季的9个属分别是青霉属(Penicillium)、曲霉属(Aspergillus)、拟青霉属(Paecilomyces)、木霉属(Trichoderma)、毛壳属(Chaetomium)、帚霉属(Scopulariopsis)、金孢属(Chrysosporium)、毛霉属(Mucor)和镰孢属(Fusarium);优势属为青霉属(Penicillium)、曲霉属(Aspergillus)和拟青霉属(Paecilomyces).夏季8个属分别有木霉属(Trichoderma)、曲霉属(Aspergillus)、链格孢属(Alternaria)、青霉属(Penicillium)、毛霉属(Mucor)、伏革菌属(Corticium)、拟青霉属(Paecilomyces)和镰孢属(Fusarium);优势属为木霉属(Trichoderma)、曲霉属(Aspergillus)和链格孢属(Alternaria).秋季11属包括木霉属(Trichoderma)、曲霉属(Aspergillus)、青霉属(Penicillium)、粘帚霉属(Gliocladium)、根霉属(Rhizopus)、枝孢属(Cladosporium)、踝节菌属(Talaromyces)、毛霉属(Mucor)、毛壳属(Chaetomium)、拟青霉属(Paecilomyces)和镰孢属(Fusarium);优势属为木霉属 (Trichoderma)、曲霉属(Aspergillus)和青霉属(Penicillium).冬季6属包括木霉属(Trichoderma)、曲霉属(Aspergillus)、青霉属(Penicillium)、枝孢属(Cladosporium)、镰孢属(Fusarium)和帚霉属(Scopulariopsis);优势属为木霉属(Trichoderma)和曲霉属(Aspergillus).结果见表1.

表1 季节对芦苇根际土壤可培养真菌数量及种群构成的影响
在春、夏、秋、冬4个季节中,芦苇根际土壤可培养真菌的Shannon多样性指数、Margalef丰富度指数,从大到小的次序排列为秋季>春季>夏季>冬季,而Pielou均匀度指数则不同,为春季>夏季>冬季>秋季;Shannon多样性指数和Margalef丰富度指数在秋季均为最高,而均匀度指数则是春季最高,并且在4个季节中,多样性指数和丰富度指数的数值波动较大,均匀度指数相对差异较小且秋季和冬季的数值相近.说明季节对芦苇根际土壤可培养真菌的多样性指数和丰富度指数影响较大、对均匀度指数影响较小.比较4个季节可培养真菌种类的相似性指数,可得出相似性指数(Cj)值春季和夏季为 0.545 5、春季和秋季为 0.538 5、春季和冬季为0.500 0、夏季和秋季为 0.461 5、夏季和冬季为0.400 0、秋季和冬季为0.307 7,其中,春季与夏季和秋季为中等相似,其他季节为中等不相似(见表2).
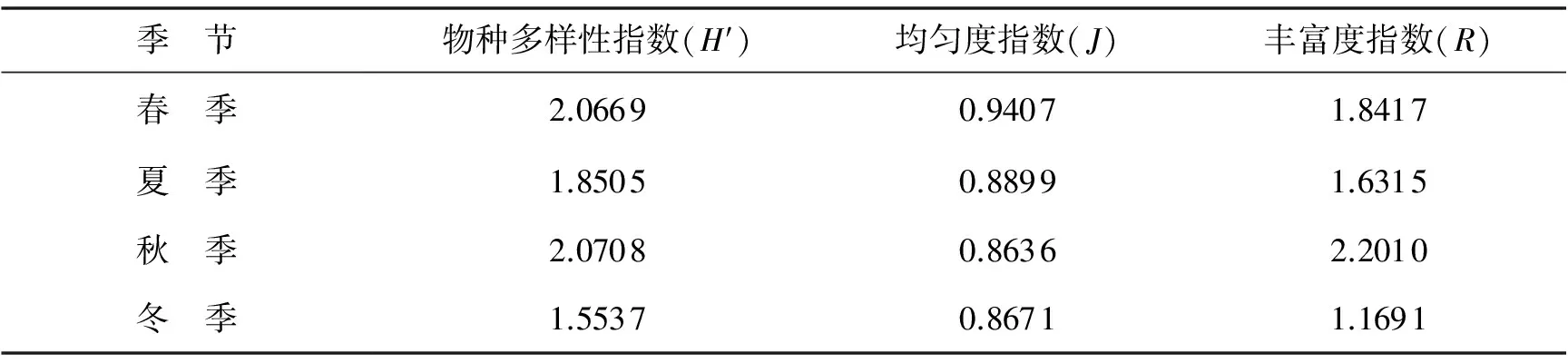
表2 季节对芦苇根际土壤可培养真菌种群多样性指数的影响
2.2 芦苇对土壤可培养真菌多样性保护作用的分析
表3为不同季节下的无植被土壤中的可培养真菌种群构成及数量.对比芦苇根际土壤和无植被土壤真菌种群构成可知,芦苇根际土壤含有伏革菌属、粘帚霉属、踝节菌属、金孢属和根霉属等真菌,它们是无植被土壤中没有的菌属;而无植被土壤中也含有一些芦苇根际土壤中没有的菌属:枝顶孢属、散孢菌属和白僵菌属.对比表1和表3可知,芦苇根际土壤可培养真菌种类是无植被土壤的1.2倍,数量是无植被土壤的4.0倍.芦苇根际土壤可培养真菌与无植被土壤可培养真菌多样性指数(H′)的R/S值为1.03~2.20,均匀度指数的R/S值为0.93~1.20;芦苇根际土壤可培养真菌的多样性指数高于无植被土壤,但均匀度指数与无植被土壤的相似.2种土壤所含的真菌种类差异性较大,且种群构成也不同(见表4).因此,芦苇根际土壤相对于无植被土壤具有丰富的真菌资源,芦苇对土壤真菌多样性的保护具有重要作用.

表3 季节对无植被土壤可培养真菌种群构成及数量的影响

表4 芦苇根际土壤与无植被土壤中的可培养真菌多样性指数与均匀度指数的比较
注:R/S为根际土壤/无植被土壤的数值比
3 讨 论
本课题组前期研究得知[11],芦苇内生真菌的优势属为镰孢菌属(Fusarium)、青霉属(Penicillium)和链格孢属(Alternaria),而本文研究的芦苇根际土壤真菌的优势属有链格孢属(Alternaria)、青霉属(Penicillium)、曲霉属(Aspergillus)、拟青霉属(Paecilomyces)和木霉属(Trichoderma).可见芦苇内生真菌种类和根际土壤真菌种类相似,但根际土壤中真菌种类的多样性较高.本课题组成员邵璐等人[8]曾对生长于同一地域的辽宁碱蓬根际土壤真菌多样性进行过研究,查明了辽宁碱蓬根际土壤真菌的优势属有青霉属(Penicillium)、枝孢属(Cladosporium)、葡萄穗霉属(Stachybotrys)、木霉属(Trichoderma)、 曲霉属(Aspergillus)和镰孢属(Fusarium);本文采集的芦苇根际土样与辽宁碱蓬根际土样来自于同一采样区,2种植物的根际是否有它们独特的菌种资源,本文予以查明.对比芦苇和辽宁碱蓬根际土壤中的真菌种类,优势菌属都有青霉属(Penicillium)、曲霉属(Aspergillus)和木霉属(Trichoderma);无植被土壤中也含有这些菌属,说明这些菌属对环境适应性较强,受生态条件影响较小,是土壤中重要的优势种群[12].芦苇不同于辽宁碱蓬的菌属有帚霉属(Scopulariopsis)、粘帚霉属(Gliocladium)、毛壳属(Chaetomium)、踝节菌属(Talaromyces)、伏革菌属(Corticium)、根霉属(Rhizopus)和金孢属(Chrysosporium);本研究还查明了芦苇根际土壤中的真菌种类资源丰富,其中金孢属、伏革菌属、粘帚霉属、踝节菌属和根霉属是无植被土壤中没有的菌属.
芦苇的抗逆性较强,素有“禾草森林”之称[13],是构建人工湿地、修复污染土壤的常用植物[14].芦苇的修复是以根际效应为主,其根际恰当的微生物类群为土壤污染物降解提供了有力的保障[8].本研究从芦苇根际土壤中分离出了15 属54 种真菌,说明了真菌资源的多样性和丰富性.在芦苇根系分泌物滋养下,一定孕育着丰富的、有经济价值的微生物种类资源,期望今后能从中筛选到更多的真菌、细菌、放线菌等有益菌种,为芦苇生境的养护及以芦苇-微生物修复被污染土壤提供理论依据及有益菌株.Ma Fang等人[14]曾以变性梯度凝胶电泳(DGGE)法研究了湿地芦苇根际的微生物群落特征,证明了微生物在芦苇根际的恒常性和可变性与湿地生态系统的时空变化有关,根际的微生物群落似乎在很大程度上受营养生长期的影响.本文对芦苇根际土壤中可培养的真菌进行了检测及季节对其多样性的影响分析,其结果也说明了芦苇根际土壤中的可培养真菌数量及种类受季节即营养生长期的影响较大.
[1] CHAMBERS R M,MCYERSON L A,SALTONATALL K.Expansion ofPhragmitesaustralisinto tidal wetlands of north America[J].Aquatic Botany,1999,64(3/4):261-273.
[2] ZHU X Y,WANG S M,ZHANG C L.Responses of different ecotypes of reed growing in the Hexi Corridor to natural drought and salinity[J].Plant Physiology Communications,2003,39(4):371-376.
[3] LI J G,LI G B,LIU F,et al.Reed resource and its ecological function and utilization in Baiyangdian Lake[J].South-to-North Water Transfers and Water.Science and Technology,2004,2(5):37-40.
[4] RUTIGLIANO F A,ASCOLI R D,VIRZO DE SANTO A.Soil microbial metabolism and nutrient status in a Mediterranean area affected by plant cover[J].Soil Biology & Biochemistry,2004,36(11):1719-1729.
[5] HOLMES G D,HALL N E, GENDALL A R, et al.Using transcriptomics to identify differential gene expression in response to salinity among AustralianPhragmitesaustralisclones[J].Frontiers in Plant Science,2016,6(7):1-12.
[6] FENG H,QIAN Y, GALLAGHER F J, et al.Synchrotron micro-scale measurement of metal distributions inPhragmitesaustralisandTyphalatifoliaroot tissue from an urban brownfield site[J].Journal of Environmental Sciences,2016,41(3):172-182.
[7] 王振庆,王丽娜,吴大千,等.中国芦苇研究现状与趋势[J].山东林业科技,2006(6):85-87,74.
[8] 邵璐,姜华.辽宁碱蓬根际土壤真菌多样性的季节变化及其耐盐性[J].生态学报,2016,36(4):1050-1057.
[9] 魏景超.真菌鉴定手册[M].上海:上海科学技术出版社,1979:495-496,501.
[10] 姜华,杨策,石鑫.大连市银杏叶斑病病原菌鉴定[J].辽宁师范大学学报(自然科学版),2014,37(3):396-401.
[11] 叶馨,陆蕾,丛珊,等.芦苇内生真菌的多样性分析[J].辽宁农业科学,2016(1):5-8.
[12] BOSSIO D A,SCOW K M,GUNAPALA N,et al.Determinants of soil microbial communities:effects of agricultural management,season and soil type on phospholipids fatty acis profiles[J].Microbial Ecology, 1998,36(1):1-12.
[13] 吴洁婷.湿生芦苇根际微生物群落结构与功能研究[D].哈尔滨:哈尔滨工业大学,2011.
[14] MA F,WU J T,WANG L,et al.Characterization of the microbial community in the Rhizosphere ofPhragmitesaustralis(Cav.) Trin ex.Steudel growing in the Sun Island Wetland[J].Water Environment Research,2014,86(3):258-268.
[15] 庄瑶,孙一香,王中生,等.芦苇生态型研究进展[J].生态学报,2010,30(8):2173-2181.
Seasonal variation of soil fungal diversity in the rhizosphere ofPhragmitesaustralis
JIANGHua1,LIYanchao1,JIANGYi2
(1.College of Life Science, Liaoning Normal University, Dalian 116081, China;2.Animal & Plant & Foodstuffs Inspection Center, Tianjin Entry-Exit Inspection and Quarantine Bureau, Tianjin 300456, China)
In this paper, rhizosphere soil fungis were isolated and identified by using dilution-plate method, morphological analysis and molecular biology identification method, and the effect of season on fungi were analyzed. The results show that in spring, summer, autumn and winter, the number of fungal colonies involved in the rhizosphere soil is 5 243,5 003,6 388 and 4 970 respectively. They belong to 9 genera 21 species, 8 genera 18 species, 11 genera 22 species and 6 genera 21 species, respectively, totally there were 54 genera and 15 species of fungi isolated.The dominant genera areAspergillus,Penicillium,Paecilomyces,Trichoderma, andAlternaria.The Shannon diversity index and richness index of rhizosphere fungi from different seasons were ranked as autumn>spring>summer>winter,while the evenness index was spring>summer>winter>autumn. It can be seen that the diversity and richness index showed the highest performance in autumn, but the numerical value fluctuated greatly, and the evenness index showed a little difference.P. australis fungi resources are abundant in rhizosphere soil.Among them,Chrysosporium,Corticium,Gliocladium,TalaromycesandRhizopusare the genera which can not be found in the non-vegetation soil;The total species ofP. australis rhizosphere fungi is 1.2 times than those without vegetation covering,and the quantity is 4 times of non-vegetation soil.P. australis plays an important role in the protection of soil fungal diversity.
Phragmitesaustralis;rhizosphere soil;fungal diversity
2016-11-25 作者简介:姜华(1959-),女,黑龙江富裕人,辽宁师范大学教授,博士.
1000-1735(2017)01-0089-06
10.11679/lsxblk2017010089
S763.15
A
