三种植物气体交换参数的日变化特征
2017-04-12丁旭李宏侠刘建国霍举松刘
丁旭++李宏侠++刘建国++霍举松++刘卫国++李春
摘要:以三工河流域3種植物胡杨(Populus euphratica)、芦苇(Phragmites communis)和柽柳(Tamarix chinensis)为研究对象,利用LI-6400便携式光合作用测定系统测定了3种植物的光合速率、蒸腾速率、气孔导度等气体交换参数,分析3种植物对不同土壤条件的响应以及植物水分利用效率和光能利用效率的日变化特征。结果表明,①芦苇对不同的土壤环境适应能力最强,胡杨主要受水分影响,柽柳受到水盐及其他环境要素的共同影响,适合于水盐不要过高的地方;②芦苇和胡杨的水分利用效率在水分充足时低于水分较低时,而柽柳的水分利用效率变化随土壤水分变化不大;③3种植物的光能利用效率均随水盐含量减少而增加。芦苇在不同生境中均表现极强的生态适应性,胡杨的生理特性和分布范围主要受水分影响,柽柳在水盐生境中也表现为极强的适应性,其主要受干旱胁迫作用,在胁迫生境中3种植物主要通过提高自身的光能利用效率来抵御胁迫危害。
关键词:光合速率;蒸腾速率;气孔导度;水分利用效率
中图分类号:Q945.1 文献标识码:A 文章编号:0439-8114(2016)22-5802-05
DOI:10.14088/j.cnki.issn0439-8114.2016.22.019
Diurnal Variations Characteristics of Three Plants Gas Exchange Parameters under Different Water Conditions
DING Xu, LI Hong-xia, LIU Jian-guo, HUO Ju-song, LIU Wei-guo, LI Chun
(College of Resources and Environment Science/Key Laborary of Oasis Ecology,Ministry of Education,Xinjiang University,Urumqi 830046,China)
Abstract: Taking three native dominant desert plants Populus euphratica, Phragmites communis, Tamarix chinensis of Sangong River basin as the research object, different gas exchange parameters, such as photosynthetic rate, transpiration rate,stomatal conductance and so on of these plants had been measured using the LI-6400 portable photosynthesis system. The response of plant to different soil conditions, the diurnal variation characteristics of water use efficiency(WUE) and light use efficiency (LUE) of these plants were investigated. The results showed that:①P. australis had strongest resilience towards different soil environment. P. euphratica was mainly influenced by moisture, while T. chinensis was influenced by both soil water salt and other environment factors, which was not suitable for high water salt. ②Under sufficient water condition, P. australis and P. euphratica higher water use efficiency rather than low moisture contents, but water use efficiency of T. chinensis could not change a lot with changes of soil moisture.③Radiation use efficiency of all three plants could increase with decreased water salt. P. australis for different habitats showed strong ecological adaptability, P. euphratica mainly by moisture on physiological characteristics and distribution. T. chinensis habitat in the water and salt also showed strong adaptability, its role was mainly affected by drought stress. In the stress habitats, three plants mainly to improve their energy efficiency to withstand stress hazards.
Key words: photosynthetic rate; transpiration rate; stomatal conductance; water use efficiency
土壤条件影响植物的生理和生长特征,限制植物的光合和个体生长发育[1],而光合作用和蒸腾作用是反映植物对各种内外因子响应的生理指标。近年来,在植物生理生态方面,随着便携式测定仪器测试技术的发展,实现了在野外自然状态下测定植物的气体交换过程[2]。胡杨(Populus euphratica)、柽柳(Tamarix ramosissima)、芦苇(Phragmites communis)是三工河流域的主要荒漠植物,研究这3种植物的生理生态特性对三工河流域植被的生态恢复有重要的理论和实践意义。通过植物的气体交换参数研究植物的光合作用及其与环境的关系,国内外已有报道,但主要围绕某一种植物在试验条件(遮阴、盐分胁迫以及干旱条件)下的研究[3-7],或是植物不同生态型以及不同时期之间的特征比较[8-12],而对于多种植物在自然条件下对不同土壤环境条件的气体交换参数特征的报道较少。因此,本试验通过对三工河流域不同土壤环境条件下3种植物的气体交换参数的测定和分析,研究它们对不同土壤条件的生理生态适应性和调节机理,为研究该区荒漠植物对不同水盐的生态响应提供理论依据。
1 研究区概况
阜康县位于博格达峰北麓,准格尔盆地东南缘。三工河流域位于天山南麓中段东部,自西向东主要有3条河流,分别是水磨沟河、三工河、四工河,发源地均为博格达山北麓的冰雪林[13]。地势南高北低,分为南部山地,中部平原,北部沙漠。冬季寒冷漫长,夏季干旱炎热,主要为大陆性干旱气候。境内的主要河流大都发源于天山水源,是全县灌溉农业的主要水源。全县植被呈明显的规律性,自上而下为高山寒漠、高山草甸、森林、草原、荒漠植被、沙漠[14-16]。
2 材料与方法
2.1 研究材料
本试验以三工河流域胡杨、芦苇和柽柳3种植物为研究对象,于2014年7月至8月初前往研究区采集试验数据。观测时间为2014年7月底至8月初,植物生长旺盛期。结合三工河流域地下水监测位置,在研究区布设多个试验点,本研究采用其中的4个点即14号点(绿洲内部)、16号点(盐化沙地)、18号点(过渡带)和23号点(沙漠内部)。试验点植被覆盖率较高,主要以胡杨、芦苇和柽柳为主,并且有红柳、沙枣、锦鸡儿、骆驼蓬、蒿子等。虽然植被覆盖率较高,但风沙较大降水较少。
2.2 测量方法
利用LI-6400便携式光合作用测定系统(美国LI-COR公司)进行净光合速率(Pn,μmol CO2/m2·s)、蒸腾速率(Tr,mmol H2O/m2·s)等生理指标测定,该仪器同时也可记录气温(Ta,℃)、叶温(Tl,℃)、空气相对湿度(RH,%)、光合有效辐射(PAR,μmol/m2·s)等指标参数。数据测量选择晴朗无云的天气进行,样地设在植被未受破坏,长势均匀,并具有代表性的地方,设置面积为30 m×30 m的样方。每种植物(胡杨、芦苇和柽柳)随机选择3株长势良好的植株,在植株向阳面大致相同部位选取生长状况良好、叶龄相似且互不遮光的叶片,將枝条拉至同一高度进行活体测定,每片叶每次测定重复3次。在北京时间8:00~20:00(文中时间均为北京时间)每2 h测定一次[2]。测量时尽量将植物叶片平铺于叶室内,并避免相互遮挡,整个叶室能充满发散光,使叶室中所有的叶表面均能同等地接受光照[1]。
水分利用效率(Water use efficiency,WUE)是反映植物耐旱性的指标,显示植物有效利用水分的能力。水分利用效率还是重要的生理特征,指示植物对干旱的适应[17-20],表达式为WUE=Pn/Tr,单位为μmol CO2/mmol H2O,Pn为单位叶面积上叶片的净光合速率;Tr为单位叶面积的蒸腾速率[1]。光能利用效率(LUE)是单位土地面积上植物通过光合作用所产生的有机物中所含的能量与这块土地所接受的太阳能的比,植物可以通过提高光能利用效率来适应干旱环境[1,21,22],光能利用效率(LUE)是植物光合作用量子效率的固定参数,表达式为LUE=Pn/PAR,单位为μmol CO2/μmol,PAR为光合有效辐射[1]。
2.3 统计分析
采用SPSS19.0、Origin 8.0等工具对数据加以处理和分析,绘制出3种植物的气体交换参数的日变化曲线图。数据分析采用单因素方差分析和Duncan多重比较。
3 结果与分析
3.1 各试验点土壤理化特征
从表1可以看出,盐化沙地的土壤水分含量最高达24.7%,其次是绿洲内部,水分较充足,土壤含水量为10.6%,过渡带和沙漠内部的土壤水分含量较低,分别为2.59%和1.70%,即水分含量由大到小依次为盐化沙地、绿洲内部、过渡带、沙漠内部。各试验点土壤pH均呈碱性,绿洲内部最高,达9.19,盐化沙地、过渡带和沙漠内部依次减少,分别为8.78、8.19和7.85。对于有机碳和有机质含量,均为沙漠内部最高,其他3个试验点含量相差不大。而对于电导率、总盐以及各离子含量(K+、Na+、Cl-),盐化沙地均明显高于其他各点,沙漠内部均为最低。绿洲内部和过渡带的各离子含量(K+、Na+、Cl-)相差不大,绿洲内部略高,但电导率和总盐相差较大,电导率分别为4.69、2.84 ms/cm,总盐分别为20.700、8.650 g/kg。
3.2 三种植物的气体交换参数的日变化特征
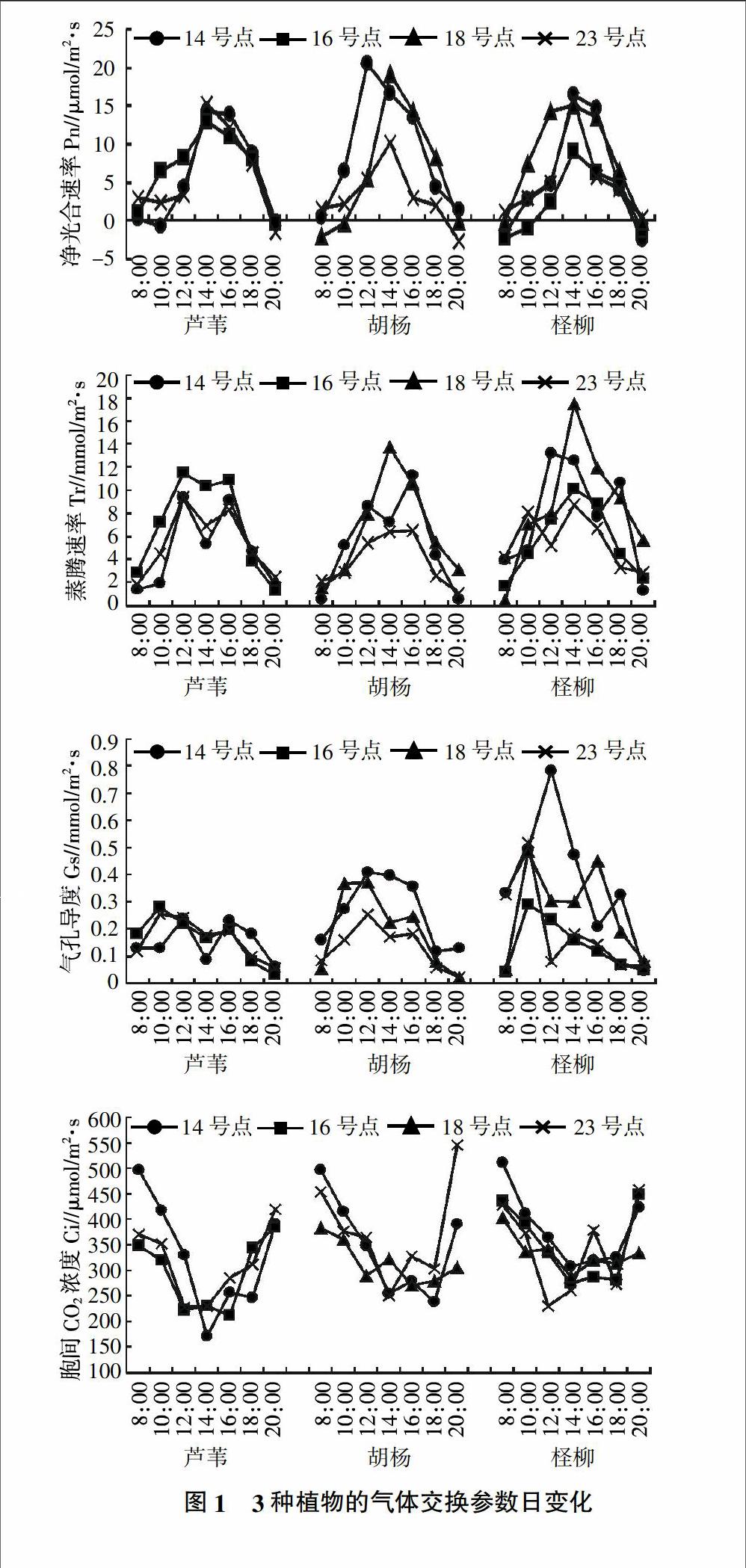
不同土壤环境条件下3种主要荒漠植物的气体交换参数日变化见图1。由图1可知,在不同土壤条件下,3种植物的净光合速率变化均呈单峰曲线。土壤环境条件的变化对芦苇的净光合速率影响并不是很大,峰值无显著差异且均出现在14:00左右。胡杨净光合速率在不同土壤环境下有较大的变化,沙漠内部净光合作用较低,明显低于其他试验点;过渡带和沙漠内部的峰值出现在14:00左右,但在绿洲内部,光合速率的峰值出现在12:00左右,可能与土壤含盐量有关,有研究指出,随盐分增大,峰值时间提前[23]。柽柳的净光合速率也均呈单峰,峰值出现在14:00左右,各试验点的净光合速率日变化有差异,盐化沙地的净光合速率最低,与其他试验点有明显差异,而其他试验点间差异不大。各植物在不同的土壤条件下,净光合速率达到峰值前差异明显,在峰值过后无明显差异。3种植物中,同样土壤条件下胡杨的光合速率最大,达到20.47 μmol/m2·s,其次是柽柳,最后是芦苇,但差异不明显。在所研究的试验点中,盐化沙地植物的净光合速率峰值并不是最高,这与盐化沙地的土壤盐含量有关,盐分增大,会使峰值降低。净光合速率不仅与土壤水盐有关,还与光有效辐射、温度以及胞间CO2浓度有关系。
由图1可以看出,不同土壤条件下植物的蒸腾速率变化各不相同。芦苇的蒸腾速率全部呈双峰曲线,峰值分别在12:00和16:00左右,在14:00降低;胡杨在绿洲内部出现双峰,沙漠内部呈现平滑的单峰曲线,过渡带出现明显的单峰,出现单峰有可能是因为测量时间间隔不够密集,没有检测到;柽柳的蒸腾速率在不同的土壤环境下变化较大,峰值较高,曲线有双峰也有单峰。盐化沙地和过渡带呈现单峰曲线,峰值出现在14:00左右,沙漠内部和绿洲内部的曲线呈双峰,峰值出现的时间不同,沙漠内部的峰值出现在10:00和14:00,绿洲内部的峰值出现在12:00和18:00,盐化沙地和沙漠内部的峰值较低,绿洲内部和过渡带的峰值较高。柽柳和芦苇在12:00~18:00之间水分蒸腾速率持续较大。胡杨的蒸腾作用主要集中在14:00左右。从气孔导度的日变化和蒸腾速率的日变化图中可以看出,植物的蒸腾速率与气孔导度密切相关,气孔导度降低,说明植物在此时关闭了气孔防止水分蒸发。
3种植物在不同的土壤环境条件下,胞间CO2浓度的日变化规律基本一致,早晨和晚上较高,中午较低,这是因为早晨光合作用很弱,而白天随着光合作用的增强,胞间CO2浓度逐渐减少。植物的光合速率和蒸腾速率与气孔有关,气孔通过张开闭合控制胞间CO2浓度和植物体内水分来调节光合作用和蒸腾作用。
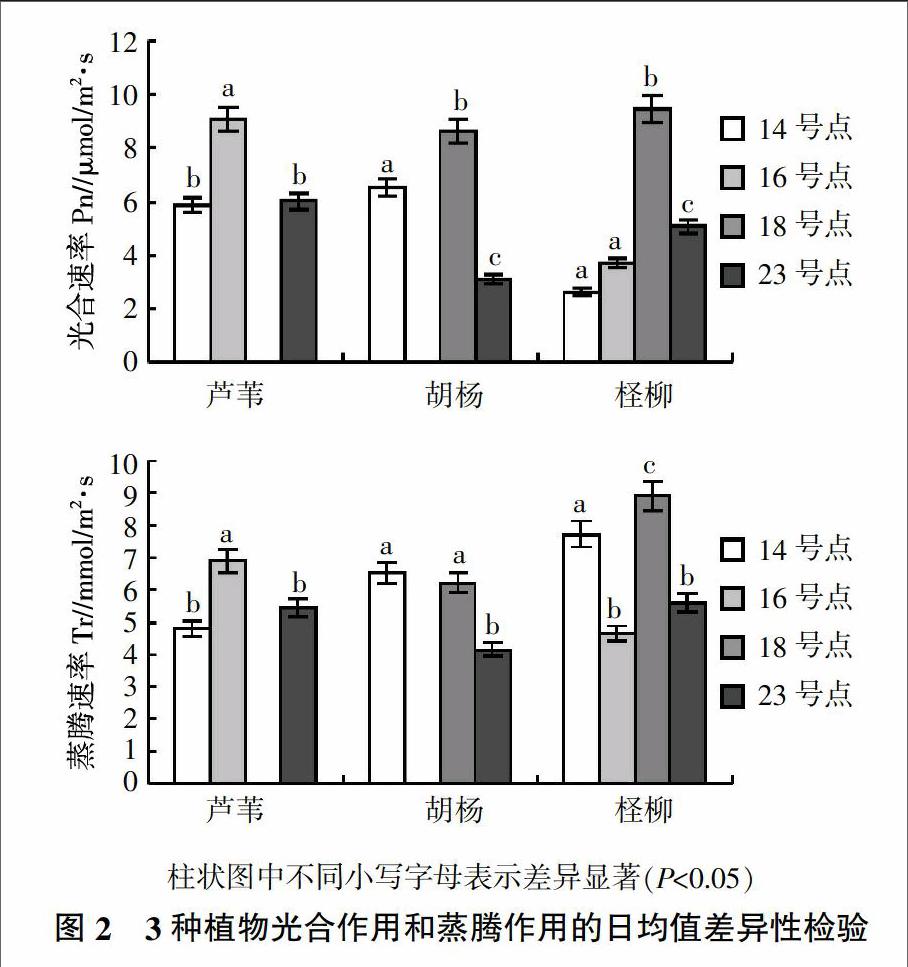
3种荒漠植物的光合速率和蒸腾速率日均值的差异性检验见图2。由图2可知,不同植物对不同环境的响应差异明显。芦苇的光合速率和蒸腾速率的日均值在盐化沙地与其他样点差异显著,芦苇对水盐的响应较明显,随水盐变化而变化,适应性比较好;胡杨的光合速率日均值在绿洲内部、过渡带、沙漠内部差异显著,蒸腾速率日均值在绿洲内部和盐化沙地差异不显著,在沙漠内部显著低于与其他两点,胡杨的光合速率比蒸腾速率对水盐更敏感些;柽柳的光合速率日均值在绿洲内部和盐化沙地无显著性差异,在过渡带日均值最高,达9.45 μmol/m2·s,明显高于其他各点,蒸腾速率日均值在盐化沙地和沙漠内部无显著性差异,在过渡带最高,说明过渡带的环境更适合柽柳生长,过高的水分和盐分会抑制柽柳的生长。
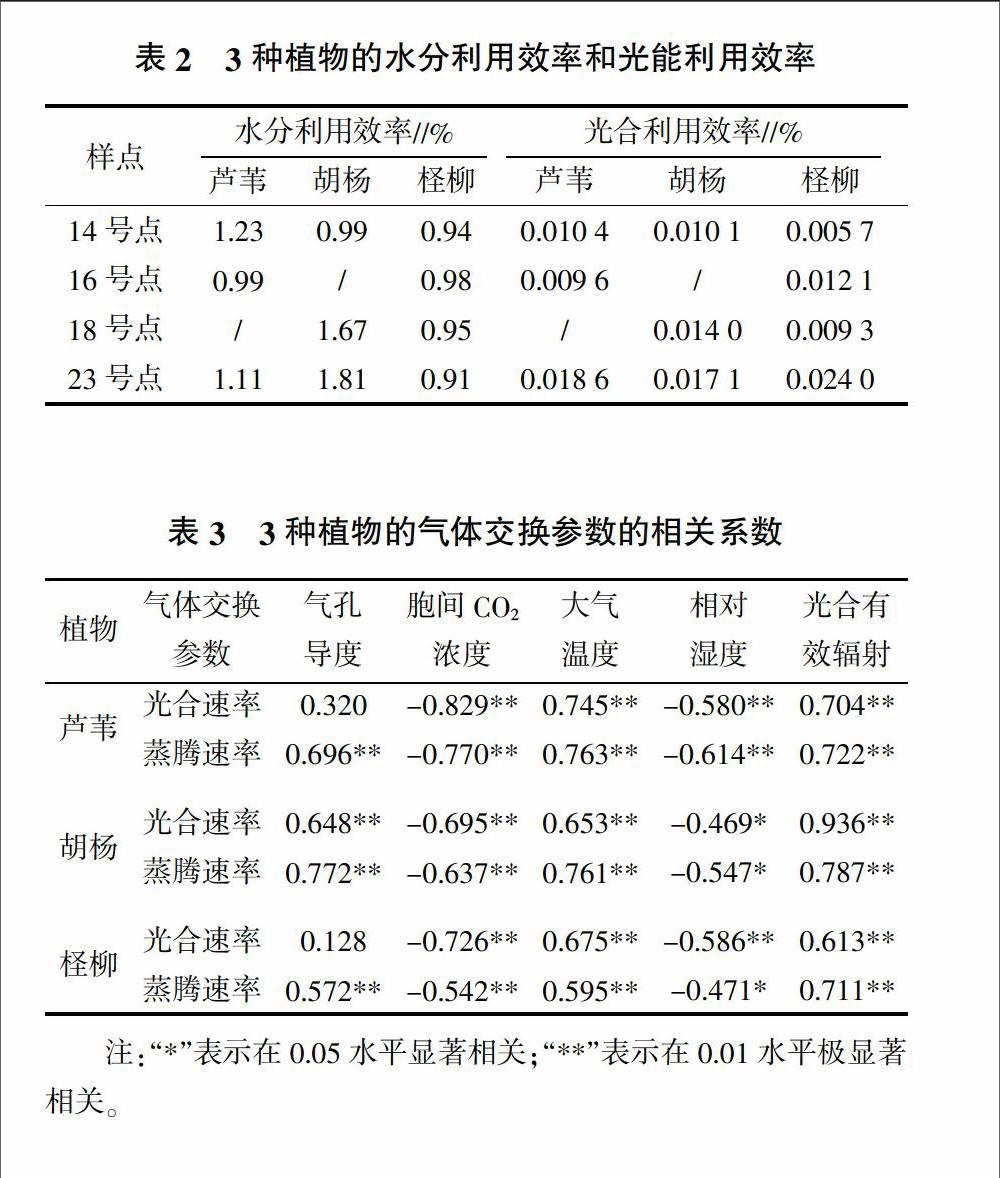
3.3 水分利用效率和光能利用效率
由表2可知,当水分条件较差时,植物会采取降低蒸腾,提高水分利用效率以适应环境。芦苇的水分利用效率在水分充足时低于水分较低时,在绿洲内部水分利用效率最高。胡杨的水分利用效率更符合上述结论。柽柳的水分利用效率变化随土壤水分变化不大,由于其是深根系植物,可利用更深层的土壤水分,本试验土壤研究层较浅,未达到影响柽柳的深度。3种植物中胡杨的水分效率最高,达1.81,可得胡杨最适应土壤水分的变化。光能利用效率规律同水分利用效率表现相似,3种植物的光能利用效率均隨水分减少而增加,这体现植物利用提高光能利用率以适应更干旱的环境,其中芦苇和胡杨的光能利用效率高于柽柳,这与其自身强生态适应性有关。
3.4 3种植物的气体交换参数的相关性分析
从表3可以看出,植物的光合速率和蒸腾速率均受到多种环境因子影响,光合速率与胞间CO2浓度、大气温度、相对湿度、光合有效辐射等密切相关,蒸腾速率除与上述因子相关外还同气孔导度密切相关。3种植物的光合速率和蒸腾速率与大气温度和光合有效辐射呈正相关,且相关程度达极显著水平;与胞间CO2浓度和相对湿度呈负相关,其与胞间CO2浓度的相关程度达极显著水平;与相对湿度的相关程度中,芦苇达极显著相关水平,胡杨达显著相关水平,柽柳的光合速率和蒸腾速率分别与相对湿度呈极显著负相关与显著负相关;3种植物的蒸腾速率与气孔导度呈极显著正相关,光合速率与气孔导度呈正相关,但只有胡杨达极显著水平。
4 小结与讨论
不同植物对不同土壤环境条件的响应各不相同。环境变化对芦苇的光合作用影响较小;柽柳的光合速率峰值受到盐分影响,会受到抑制;而胡杨的净光合速率受水分影响较大。芦苇的蒸腾速率全部呈双峰曲线;胡杨的蒸腾速率曲线随着水盐的增加,由单峰转变为双峰且峰值增大;柽柳的蒸腾速率曲线有双峰也有单峰,蒸腾速率的峰值均出现在正午左右。
植物通过降低蒸腾,提高水分利用效率以及光能利用效率来适应干旱环境,为节约水分,干旱生境的许多植物采取了增加气孔阻力的形态、结构和生理机制,由于蒸腾速率对气孔关闭的敏感度高于光合速率,因此一般认为干旱提高了植物的水分利用效率。
植物的光合速率和蒸腾速率受到多种环境因子的影响。通过检验,光合速率与胞间CO2浓度、大气温度、相对湿度、光合有效辐射密切相关,蒸腾速率除与上述因子相关外还与气孔导度密切相关。
三工河流域荒漠植物在光合速率、蒸腾速率及气孔导度上对环境条件的改变所做出的响应反映了不同植物的适应方式。在生态环境条件较好时,植物充分利用各种环境资源,保持较好的生活状态,而植物的净光合速率和蒸腾速率主要受光照强度和环境气候条件的影响;水分条件较差时,荒漠植物会采取降低蒸腾,提高水分利用效率和光能利用率来适应不利的生态环境。不同植物的调节机理不同,适应环境的相关参数变化程度也表现出较大的差异性[23-26]。
从以上分析结果可得以下结论:①芦苇对不同的土壤环境适应能力更强,胡杨主要受水分影响,柽柳受到水盐及其他环境要素的共同影响,适合于水盐不宜过高的环境中。②芦苇和胡杨的水分利用效率在水分充足时低于水分较低时,而柽柳的水分利用效率变化随土壤水分变化不大。3种植物的光能利用效率均随水盐减少而增加。③3种荒漠植物的光合速率和蒸腾速率与气孔导度、大气温度和光合有效辐射呈正相关,与胞间CO2浓度和相对湿度呈负相关,但不同植物的光合速率和蒸腾速率与各因子的相关程度呈现不同的显著水平。
参考文献:
[1] 王珊珊,陈 曦,王 权,等.新疆古尔班通古特沙漠南缘多枝柽柳光合作用及水分利用的生态适应性[J].生态学报,2011,31(11):3082-3089.
[2] 陈亚鹏,陈亚宁,徐长春,等.塔里木河下游地下水埋深对胡杨气体交换和叶绿素荧光的影响[J].生态学报,2011,31(2):344-353.
[3] GAO S,SU P X,YAN Q D,et al. Canopy and leaf gas exchange of Haloxylon ammodendron under different soil moisture regimes[J].Science China Life Sciences,2010,53(6):718-728.
[4] 王建华,任士福,史宝胜,等.遮荫对连翘光合特性和叶绿素荧光参数的影响[J].生态学报,2011,31(7):1811-1817.
[5] 张国伟,张 雷,唐明星,等.土壤盐分对棉花功能叶气体交换参数和叶绿素荧光参数日变化的影响[J].应用生态学报,2011, 22(7):1771-1781.
[6] 王 凯,朱教君,于立忠,等.遮阴对黄波罗幼苗的光合特性及光能利用效率的影响[J].植物生态学报,2009,33(5):1003-1021.
[7] 王伟华,张希明,闫海龙,等.盐处理对多枝柽柳光合作用和渗调物质的影响[J].干旱区研究,2009,26(4):561-568.
[8] 刘卫国,邹 杰.水盐梯度下克里雅河流域芦苇光合响应特征[J].西北植物学报,2014,34(3):572-580.
[9] 苏培玺,严巧娣.C4荒漠植物梭梭和沙拐枣在不同水分条件下的光合作用特征[J].生态学报,2006,26(1):75-82.
[10] 苏培玺,赵爱芬,张立新,等.荒漠植物梭梭和沙拐枣光合作用、蒸腾作用及水分利用效率特征[J].西北植物学报,2003,23(1):11-17.
[11] 龚吉蕊,赵爱芬,苏培玺,等.黑河流域几个主要植物种光合特征的比较研究[J].中国沙漠,2005,25(4):587-592.
[12] 黄 刚,赵学勇,崔建垣,等.水分胁迫对2种科尔沁沙地植物光合和水分利用特性的影响[J].西北植物学报,2008,28(11):2306-2313.
[13] 年福华,李 新.三工河流域水量转换特征及水资源利用[J].干旱区研究,2000,17(1):5-11.
[14] 王根绪,程国栋.干旱荒漠绿洲景观空间格局及其受水资源条件的影响分析[J].生态学报,2000,20(3):363-368.
[15] 王政权.地统计学在生态学中的应用[M].北京:科学出版社,1999.
[16] 孙司衡.再生资源遥感研究[M].北京:中国林业出版社,1991.
[17] 曹生奎,冯 起,司建华,等.植物叶片水分利用效率研究综述[J].生态学报,2009,29(7):382-389.
[18] RICHARDS R A,REBETZKE G J,CONDON A G,et al. Breeding opportunities for increasing the efficiency ofwater use and crop yield in temperate cereals[J].Crop Science,2002,42(1):111-121.
[19] CASPER B B,FORSETH I N,WAIT D A. A stage-based study of drought response in Cryptantha flava (Boraginaceae):Gas exchange,water use efficiency,and whole plant performance[J].American Journal of Botany,2006,93(7):978-987.
[20] ZHAO B Z,KONDO M,MAEDA M,et al. Water-use efficiency and carbon isotope discrimination in two cultivars of upland rice during different developmental stages under three water regimes[J].Plant and Soil,2004,261:61-75.
[21] TEZARA W,MAR?魱N O,RENGIFO E,et al. Photosynthesis and photoinhibition in two xerophytic shrubs during drought[J].Photosynthetica,2005,43(1):37-45.
[22] 李 偉,曹坤芳.干旱胁迫对不同光环境下的三叶漆幼苗光合特性和叶绿素荧光参数的影响[J].西北植物学报,2006,26(2):266-275.
[23] 吴统贵,李艳红,吴 明,等.芦苇光合生理特性动态变化及其影响因子分析[J].西北植物学报,2009,29(4):789-794.
[24] 黄金廷,侯光才,尹立河,等.干旱半干旱区天然植被的地下水水文生态响应研究[J].干旱区地理,2011,34(5):788-793.
[25] 马全林,王继和,纪永福.固沙树种梭梭在不同水分梯度下的光合生理特征[J].西北植物学报,2003,23(12):2120-2126.
[26] 高 丽,杨 劼,刘瑞香.不同土壤水分条件下中国沙棘雌雄株光合作用、蒸腾作用及水分利用效率特征[J].生态学报,2009,29(11):6025-6034.
