低温寡照对番茄花期植株生长及干物质分配的影响
2017-04-05邹雨伽高冠杨再强肖芳李军
邹雨伽 +高冠 +杨再强 +肖芳 李军
摘要:为研究低温寡照复合灾害对番茄花期植株形态指标及干物质分配的影响,以金粉5号为试材,于2015年2—7月在南京信息工程大学利用人工气候室设置低温、寡照双因素控制试验,昼温/夜温分别设置18/8、16/6、14/4、12/2 ℃ 4个水平,光合有效辐射(photosynthetically active radiation,PAR)设置400、200 μmol/(m2·s) 2个水平,以昼温28 ℃/夜温18 ℃、PAR 1000 μmol/(m2·s)为对照(CK),研究不同低温寡照水平对设施番茄植株生长指标及器官干物质分配的影响。结果表明,随低温寡照胁迫加强及处理时间的增加番茄花期植株各生长指标平均增长量均有降低,昼温/夜温12 ℃/2 ℃下,PAR 200 μmol/(m2·s) 10 d处理的株高、叶面积平均增长量为最小,分别较CK低 70.12%、85.39%,PAR 400 μmol/(m2·s) 10 d处理茎粗增长量最小,较CK低94.92%。当温度相同时,PAR为400 μmol/(m2·s)[JP2]处理下的茎粗及叶面积平均增长量分别比PAR 200 μmol/(m2·s)处理高7.70%和14.28%;番茄植株地上部分干物质分配比例总体随温度升高而增大,最大值为PAR 200 μmol/(m2·s)、温度为18 ℃/8 ℃ 8 d的处理,较CK高4.57%。各处理中叶干物质分配大于茎分配比例,叶分配比例最小值为PAR 400 μmol/(m2·s)、温度 18 ℃/8 ℃ 10 d的处理,较CK低18.15%。茎干物质分配比例最小值为12 ℃/2 ℃、400 μmol/(m2·s)下2 d处理,较CK低1368%,最大值为18 ℃/8 ℃、400 μmol/(m2·s) 10 d的处理,较CK高46.11%。花器官干物质分配比例仅为3%~5%,各个处理间差异不明显。研究证实番茄花期低温寡照胁迫下干物质先向叶片分配,叶干物质分配比例随温度升高、PAR增加而降低,茎干物质分配比例变化趋势与叶相反,可为设施作物的低温寡照复合灾害防御及环境调控提供支持。
关键词:番茄;低温寡照;植株生长;干物质分配比例
中图分类号: S641.204文献标志码: A
文章编号:1002-1302(2016)12-0178-06
收稿日期:2015-10-23
基金项目:国家自然科学基金(编号:41475107);“十二五”国家科技支撑计划(编号:2014BAD10B07)。
作者简介:邹雨伽(1992—),女,四川成都人,硕士研究生,主要研究方向为设施农业气象灾害预警。E-mail:zyjxyyj@163.com。
通信作者:杨再强,博士,教授,博士生导师,主要研究设施环境调控。E-mail:yzq6751@163.com。
番茄(Lycopersicon escylentu Mill.)属茄科番茄属,为喜温性蔬菜,在世界范围内番茄产量约占到蔬菜生产总量的1356%,贸易量位居第3[1],是我国冬、春最主要的设施作物之一。番茄生长适宜温度为20~30 ℃(昼温)/15~20 ℃(夜温)。15~20 ℃/10~15 ℃为亞适温,此时番茄除植株生长减缓之外并未受到伤害;当昼/夜温度低于15 ℃/10 ℃时植株生长受到严重影响,形态开始萎蔫,内部各种生理生化反应受到影响,植株生长迟缓,持续时间过长,植株甚至死亡[2]。产量低、品质差、难以周年生产等是中国设施番茄面临的主要问题,因此如何优化调控设施小气候以提高番茄产量及品质,是生产部门亟待解决的难题。
近年来,国内外关于低温寡照对设施作物生长发育及生理特性的影响有一定研究。国外在对甜瓜[3]、番茄[4]、辣椒[5]等作物的研究中提出低温胁迫对植物的伤害主要是对地上部分,即对叶片及茎的伤害大于根系。Krause等研究认为温室作物产量对光照有很强的依赖性,温室光照强度下降,作物产量随之降低[6]。寡照对光合产物的分配运输也有影响。El-Gizawy等通过对苗期番茄进行不同程度的遮光处理,认为番茄幼苗叶面积、茎粗、总干物质含量随着光照强度的降低而降低[7]。Cockshull等则认为寡照条件与自然光环境下的番茄植株相比,株高没有太大的变化,叶面积也有所增加[8]。国内研究指出寡照会使植株叶片变大变薄,叶片叶色变淡,角度平展,枝梢变长变细,同时会抑制根系生长,植株总干质量也降低[9],这在大豆[10]、番茄[11]、甜椒[12]、黄瓜[13]等作物的研究中都已得到证实。低温胁迫程度越严重对植物伤害程度越重。余纪柱等在黄瓜低温寡照研究中指出低温弱光处理 6 d 后,与对照相比,黄瓜干质量、干鲜质量、茎粗、叶面积、叶质量、比叶面积和鲜质量都有所增加[14];在番茄[15]上的研究表明,低温弱光胁迫后,番茄茎的增长和叶面积扩张减缓,而其受影响程度取决于品种和低温弱光逆境的强度[16]。
迄今为止,国内外的研究主要集中在低温或寡照单因子对作物的影响,而忽略了两者间的协同作用,同时关于低温寡照的研究多着眼于对作物幼苗期的研究,关于低温寡照复合因子对番茄花期生长影响的研究少有报道,因此,本研究利用低温寡照双因素控制试验,研究不同低温寡照组合及不同持续时间对番茄花期生长指标及干物质分配的影响,为番茄花期的低温寡照灾害防御及环境调控提供参考。
[BT1#]1材料与方法
1.1试验设计
试验于2015年在南京信息工程大学人工气候箱(TPG-2009,Australian)进行。供试品种为金粉5号,前期在玻璃温室中用有机土分批育苗,在苗4~5张真叶时移栽于花盆中并定期定量浇水施肥,保持水分和养分在适宜的水平,待番茄出现花蕾时,选择长势相近的植株放入人工气候箱处理,每个处理重复3次。试验设置低温寡照双因素处理,各因素水平见表1。其中温度变化模拟自然气温变化趋势设置动态变温,最高温度设置在14:00,最低温度设置在5:00,各时间段温度通过人工气候箱程序控制。处理时间分别为连续2、4、6、8、10 d。处理结束后对处理植株各项生长指标进行测量,每个指标测量3次取平均值,随后放入南京信息工程大学玻璃温室内(Venlo)恢复,恢复期温光条件与对照相同,并在恢复 15 d 后分别测量各项生长指标。
1.2试验测定项目与方法
1.2.1生长指标测定
植株生长指标分别在处理及恢复结束后测量,每次取样3株,用直尺和游标卡尺分别测其株高(根茎部到生长点的距离)和茎粗(根茎部的粗度),测量3次取平均值。叶面积用直尺测量每片叶片长宽最大处长度为叶长、叶宽,并对各植株每片叶片的测量值与通过便携式叶面积仪(LI-3000C)测量值拟合得出拟合公式:
[JZ(]S=L×D×0.546 8。[JZ)][JY](1)
式中,S为单片叶面积(cm2),L和D分别为叶片长和宽(cm)。
1.2.2各器官干物质的测定与分配比例的计算
分别对低温寡照处理2、4、6、8、10 d的番茄植株进行破坏取样,并将各部分器官放入电热烘箱烘干至恒定质量后测定干质量,每个处理重复3次。地上部分分配比例是指地上部分干质量占植物总干质量的比例,地上部分各器官干物质分配比例是指植株各器官的干质量占地上部分总干质量的比例。干物質分配指数按下列公式计算[17]:
[JZ(]PIS=WSH/WT;[JZ)][JY](2)
[JZ(]PIR=1-PIS;[JZ)][JY](3)
[JZ(]PIL=WL/WSH;[JZ)][JY](4)
[JZ(]PIST=WST/WSH;[JZ)][JY](5)
[JZ(]PIF=1-PIL-PIST。[JZ)][JY](6)
式中,PIS 为地上部分干物质分配比例,PIST、PIL、PIF别为茎、叶、花器官的干物质分配比例,PIR为根干物质分配比例,WT为单株总干质量(g/株),WSH为单株地上部分干质量(g/株),WST、WL分别为单株茎、叶干质量(g/株)。
1.3数据分析
本研究数据利用Excel软件进行相关计算作图,DPS 705软件进行统计分析。
2结果与分析
2.1低温寡照对番茄花期生长指标的影响
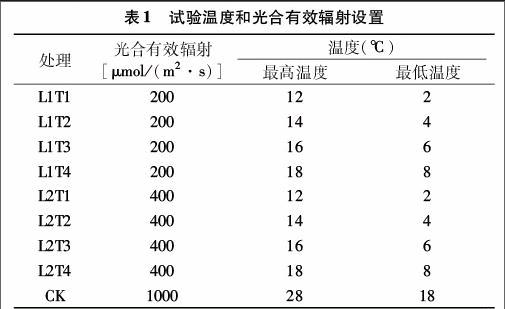
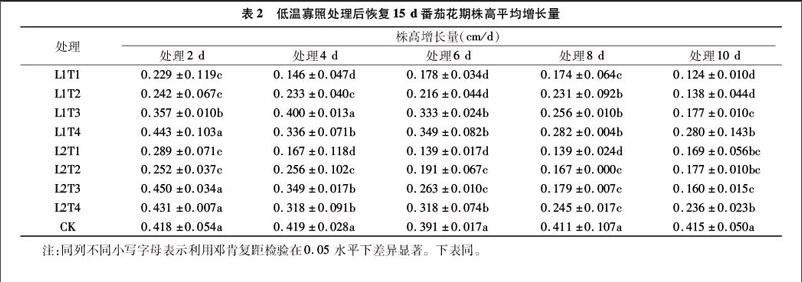
2.1.1低温寡照对番茄花期株高影响
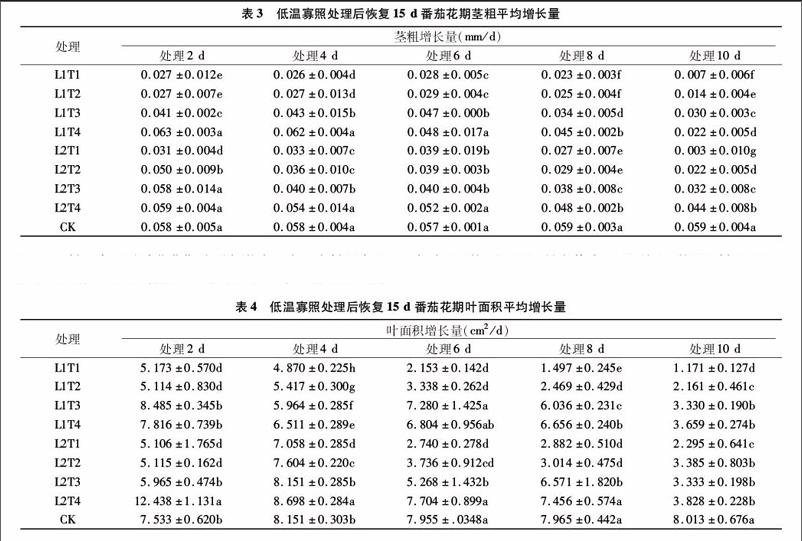
株高是对温度和光照反应十分敏感的主要农艺性状之一。表2为各个处理恢复15 d后平均株高增长量,可看出不同处理下株高的增长表现出很大的不同。低温寡照持续2 d,株高平均增长量最大值为L2T3的0.450 cm/d,较CK大7.66%;最小值为L1T1处理,为0.229 cm/d,较CK小45.21%。温度为T1、T2处理时,株高平均增长量显著低于CK。低温寡照持续4 d时,温度T1、T2、T4处理下株高平均增长量显著低于CK,最小值为L1T1处理,较CK低65.16%。当低温寡照处理持续6、8 d,各个处理平均株高增长量均显著低于CK,6 d处理最小值为L2T1,较CK小64.45%。8 d处理最小值为L2T1处理,最大值为L1T4处理,2个处理较CK分别减少66.18%、31.39%。持续10 d低温寡照,各个处理平均株高增长量显著低于CK,其中L1T1、L1T2显著低于其他处理,且T1、T2温度下,L1处理株高增长量显著低于L2处理株高增长量;L1T1为所有处理中最小,最大为L1T4处理,日平均株高增长量分别为0.124、0.280 cm/d,分别较CK低70.12%、32.53%,其中最大值为最小值的2.26倍。处理持续时间6、8 d时,相同光照条件下日平均株高增长量顺序均为T1 2.1.2低温寡照对番茄花期茎粗影响 表3为低温寡照处理恢复15 d后各个处理日平均茎粗增长量。由表3可以看出,在持续2 d的处理中T1、T2温度及L1T3处理下平均茎粗增长量显著低于CK,其余处理与CK差异不显著,甚至有大于CK的情况。其中最大值为L1T4处理,较CK大8.62%。低温寡照处理持续4 d,除L1T4处理其余各个处理均低于CK,且T1、T2、T3温度下各个处理平均茎粗增长量与CK差异显著,最小值L1T1处理为0.026 mm/d,较CK减小5517%。低温寡照处理持续6 d,各个处理平均茎粗增长量均低于CK,T1、T2、T3温度下处理与CK差异显著,最大值为L2T4处理,较CK低8.77%,最小值为L1T1处理,较CK低50.88%。低温寡照处理持续8 d,各个处理平均茎粗增长量均显著低于CK,最大值最小值分别为L2T4、L1T1处理,日平均茎粗增长量分别为0048、0.023 mm/d,较CK分别小1864%、61.02%。处理持续10 d,各个处理茎粗平均增长量均显著低于CK,其中处理L2T1日均增长量为所有处理中最小,较CK小94.92%。各个处理下同温度L2处理较L1茎粗增长量平均增加了 7.70%。在相同光照处理下,茎粗平均增长量整体随温度升高而增加,处理时间持续6 d以上时随着处理时间的延长而减小。 2.1.3低温寡照对番茄花期叶面积影响 表4为低温寡照处理后叶面积的平均增长量。在低温寡照2 d处理中T1、T2温度下平均叶面积增长量与CK差异显著,最大值为L2T4处理,较CK高65.11%,最小值为L2T1处理,平均叶面积增长量为5.106 cm2/d,较CK低32.22%。低温寡照处理持续 4 d,除L2T4、L2T3外各个处理均低于CK,其中最小值为L1T1处理,平均叶面积增长量为4.870 cm2/d。低温寡照持续6 d处理下,各个处理平均叶面积增长量均低于CK,最大值为L2T4处理,平均增长量为7.704 cm2/d,最小值为L1T1处理,平均增长量为2.153 cm2/d,2个处理分别较CK低 3.15%、7294%。低温寡照持续8 d,除L2T4各个处理日平均叶面积增长量均显著低于CK,其中L1T1平均增长量最小,仅为CK的18.79%,最大值为L2T4处理,较CK低6.39%。低温寡照持续10 d,各个处理平均叶面积增长量显著低于CK,最小值L1T1为所有处理中最小,为1.171 cm2/d,比CK低8539%,最大值为L2T4处理,较CK低52.23%。番茄花期叶面积随低温寡照处理时间的延长整体呈下降趋势,处理时间6 d及以上时,各个处理平均增长量均低于CK,且在相同光照条件下温度越低,叶面积平均增长量整体越低,处理时间持续8 d及以上时,在同一温度处理条件下L2光照下[PAR为400 μmol/(m2·s)]日平均叶面积增长量高于L1光照下[PAR为 200 μmol/(m2·s)]叶面积的增长量,同温度下各个处理L2较L1平均增加了14.28%。
2.2低温寡照对番茄花期分配比例的影响
2.2.1低温寡照对番茄花期地上部分分配比例的影响
不同处理对地上部分干物质分配如图1所示,其中,图1-a为光照强度200 μmol/(m2.s)下不同处理温度的地上部分干物质分配比例,图1-b为光照强度400 μmol/(m2.s)下不同处理温度的地上部分干物质分配比例。各个处理地上部分干物质分配比例在0.76~0.89之间,最大值为L1T4低温寡照处理8 d,为0.892,较CK高4.57%,最小值出现在L2T1持续 6 d 处理,为0.759。低温寡照持续2 d处理最小值为L2T1处理,地上部分分配比例为0.800,较CK低6.73%。低温寡照4 d,T1、T2处理地上部分分配比例均低于CK,其中L2T1、L2T2处理与CK差异显著,分别较CK低6.81%、6.00%。处理持续6 d,T1处理下地上部分分配指数均低于CK,L1T1较CK低2.25%,L2T1较CK低10.96%。低温寡照处理8 d,T1、T2温度下地上部分分配比例均小于CK,T4温度处理则大于CK,最小值为L2T1处理下的0.776,最大值為L1T4处理的 0.892。持续处理10 d,T1、T2、T3处理下地上干物质分配比例均小于CK,最小值为L2T1处理,较CK低11.88%,最大L1T4处理,地上部分干物质分配比例为0.882,但与CK差异不显著。各个处理地上部分分配比例随处理时间的延长有一个上升再下降的趋势,处理持续6 d及以上时,T4温度下地上部分分配比例均高于CK,在相同温度处理下L1[PAR为200 μmol/(m2·s)]光照处理地上部分干物质分配比例总体大于L2[PAR为400 μmol/(m2·s)]光照处理。相同光照条件下地上部分干物质分配比例总体随温度升高而增加。
[FK(W24][TPZYJ1.tif]
2.2.2低温寡照对番茄花期叶片干物质分配比例的影响
不同处理对番茄叶片干物质分配比例如图2所示,图2-a为光照强度200 μmol/(m2·s)处理下的各个温度处理的叶干物质分配比例,图2-b为光照强度为400 μmol/(m2·s)处理下各个温度处理的叶干物质分配比例。各个处理番茄叶片干物质分配比例在0.43~0.60之间。低温寡照处理2 d时,T1、T2温度下叶干物质分配比例均高于CK,且T1处理下差异显著,其中最大值为L1T1,较CK高9.37%。当低温寡照持续6 d时,T1、T2处理下叶干物质分配比例大于CK,T4处理小于CK,但与CK差异不显著,其中最大值为L2T1处理,较CK大 9.89%,最小值为L1T4处理,较CK低6.80%。处理持续 8 d,T4处理下叶干物质分配比例则显著低于CK,最大值为L2T1处理,较CK高8.26%,最小值为L1T4处理,较CK低10.64%。持续低温寡照10 d,T1、T2处理下叶干物质分配比例大于CK,其余温度下分配比例小于CK,其中L2T4处理叶干物质分配比例为所有处理中最低,较CK低1815%。低温寡照下叶干物质分配比例与地上部分分配比例变化趋势相近,均随处理时间的延长有一个上升再下降的趋势,且在T1温度处理下的叶干物质分配比例高于CK。在相同光照条件下,叶片干物质分配比例整体随温度的升高而下降。
2.2.3低温寡照对番茄花期茎干物质分配比例的影响
不同处理对番茄茎干物质分配比例如图3所示,图3-a为光照强度200 μmol/(m2·s)下各温度处理茎干物质分配比例,图3-b为光照强度400 μmol/(m2·s)下各温度处理茎干物质分配比例,可知叶干物质分配比例大于茎干物质分配比例,各个处理间茎干物质分配比例趋势与叶片干物质分配比例相反,其中茎干物质分配比例最大值为持续10 d的L2T4处理,为0.542,较CK高46.11%,最小值为连续处理2 d下L2T1处理,为0.360,较CK低13.68%。2 d处理下,茎干物质分配比例最大为L1T4处理,为0.446,较CK大7.06%,最小值与最大值均与CK差异显著。在6、8 d的处理中,T2、T3温度处理下均表现为L1[PAR为400 μmol/(m2·s)]茎干物质分配比例小于L2[PAR为200 μmol/(m2·s)]处理。相同处理天数及光照条件下,整体T4温度处理下的茎干物质分配比例最大,T1温度处理下茎干物质分配比例最小。在持续 10 d 相同光照下茎分配比例随温度升高而升高。不同处理间分配比例总体随处理时间的延长而增大。
2.2.4低温寡照对番茄花的干物质分配比例的影响图4-a为光照强度200 μmol/(m2·s)下各温度处理花干重分配比例,图4-b为光照强度400 μmol/(m2·s)下各温度处理花干重分配比例。通过试验观察到在低温寡照处理下番茄植株花器官的干物质分配比例较少,仅占植株地上部分干物质的3%~5%,各个处理间差异不显著,不具有明显变化趋势。
3结论与讨论
大多数研究表明,生长受抑是作物对温光反应最敏感的生理过程,在不适宜的温度光照度下,植株生长减缓甚至停止[CM(25][18]。本研究结果表明,低温寡照处理后,番茄花期各个生[CM)]
[FK(W22][TPZYJ4.tif]
长指标均有增长,但较CK增长缓慢,并随处理时间的延长增长速率下降,这与前人在辣椒[9]、番茄[19]、茄子[20]等作物上的研究结果一致。在光照相同的条件下,番茄茎粗平均增长随温度的升高而增大。光照为400 μmol/(m2·s)时相同温度时间处理下茎粗及叶面积日平均增长量总体大于光照 200 μmol/(m2·s) 下的处理,这可能因为番茄的生长因胁迫程度的加深而受到阻碍。
温光会影响作物库强,干扰作物干物质分配。有研究指出,弱光处理会减少番茄干物质的积累,低温对番茄干物质积累影响不显著,反而客观上有利于番茄弱光下干物质的积累[21]。本研究显示,番茄花期各个处理地上部分干物质分配比例随处理时间的延长有一个先上升再下降的趋势。相同光照条件下地上干物质分配比例随温度升高而增大,地上部分干物质分配比例在12 ℃/2 ℃及14 ℃/4 ℃处理下低于CK,在 18 ℃/8 ℃ 处理下高于CK,这与王兴银等[13]在黄瓜上的研究结论一致。其原因可能是在低温寡照前期或当低温寡照处理胁迫程度较低时,番茄根系活力逐渐下降,同时为了弥补光照不足,植株选择向上生长以获取足够的光能,这使植物体内的光合产物及养分主要用于地上部生长,减少了向地下部的输送,因此地上部分干物质分配比例上升,而低温寡照处理时间过长或低温寡照胁迫严重时,为了保持一定的根系活力和维持植株的存活,需要增加根系功能单位来修复或降低逆境的影响,光合产物较多地分配到根系,从而抑制了地上部的生长,使得地上部分分配比例下降[22]。叶干物质分配比例与地上部分干物质分配比例随处理时间变化趋势相似,整体上随处理时间的延长而降低,在相同光照条件下,叶干物质分配总体随处理温度降低而升高。茎干物质分配比例整体上随处理时间的延长而增大,在持续10 d时,相同光照下茎干物质分配比例随温度的降低而降低。这说明在低温寡照下随着胁迫程度的加深,叶片生产的干物质先满足自身需求,即干物质分配顺序为叶大于茎,与前人研究结果[9]一致。
本研究中不同光照温度组合处理下对番茄各个指标影响不同,可见低温寡照对番茄综合作用机理是复杂的,且不同品种间抗性不同。本研究对番茄低温寡照处理是通过人工气候箱,并设置温度梯度进行的,但并不能准确模拟出对照所处温度变化,同时在人工气候箱中空气湿度与外部环境水分管理也有所差异,对结果存在一定的影响。
[HS2*2][HT8.5H]参考文献:[HT8.SS]
[1]张忠杰,孔媛. 世界番茄产品贸易现状及主要出口国竞争力实证研究[J]. 新疆农垦经济,2006(7):48-53.
[2]卢良恕. 21世纪中国农业科技展望[M]. 济南:山东科学技术出版社,1993:549-557.
[3]Fogelman E,Kaplan A,Tanami Z,et al. Antioxidative activity associated with chilling injury tolerance of muskmelon(Cucumis melo L.)rind[J]. Scientia Horitculturae,2011,128(3):267-273
[4]齋藤隆.番茄生理基础[M]. 上海:上海科学技术出版社,1981:38-46
[5]Martin B,Ort D R,Boyer J S. Impairment of phoyosynthesis by chilling-temperature in tomato[J]. Plant Physiology,1981,68(2):329-334.
[6]Krause G H,Weis E. ChlorophyII fluorescence and photosynthesis:the basics[J]. Plant Biol,1991,42(42):313-349
[7]El-Gizawy A M,Gomaa H M,El-Habbasha K M,et al. Effect of different shading levels on tomato plants:growth,flowering and chemical composition[J]. Acta Hortic,1992,323:341-347
[8]Cockshull K E,Graves C J,Cave R J. The influence of shading on yield of greenhouse tomatoes[J]. Journal of Horticultural Science,1992,67:11-24.
[HT8.]
[9]郭晓冬. 低温弱光对日光温室辣椒生长及其生理功能的影响[D]. 杨凌:西北农林科技大学,2008:66-67.
[10]高辉远. 大豆生长发育过程中光合作用及光合效率的调节[D]. 泰安:山东农业大学,1999.
[11]胡文海. 低温弱光对番茄生理生化影响的研究[D]. 杭州:浙江大学,2001:47-48.
[12]眭晓蕾,蒋健箴,王志源,等. 弱光对甜椒不同品种光合特性的影响[J]. 园艺学报,1999,26(5):314-318.
[13]王兴银,张福墁. 弱光对日光温室黄瓜光合产物分配的影响[J]. 中国农业大学学报,2000,5(5):36-41.
[14]余纪柱,李建吾,王美平,等. 低温弱光对不同生态型黄瓜苗期若干测定指标及光合特性的影响[J]. 上海农业学报,2003,19(4):46-50.
[15]黄伟,任华中,张福墁. 低温弱光对番茄苗期生长和光合作用的影响[J]. 中国蔬菜,2002(4):15~17.
[16]任志雨,王秀峰,魏珉,等. 不同根区温度对黄瓜幼苗生长及光合参数的影响[J]. 山东农业大学学报:自然科学版,2003,34(1):64-67.
[17]杨再强,赵翔,苏天星,等. 光质对温室甜椒干物质生产和分配指数的影响[J]. 生态学杂志,2012,31(5):1117-1122.[HJ1.95mm]
[18]赵玉萍. 不同温度光照对温室番茄生长、光合作用及产量品质的影响[D]. 杨凌:西北农林科技大学,2010:16.
[19]Janssen L H J,Wams H E,Hass E L T,et al. Temperature dependence of chlorophyll fluorescence induction and photosynthesis in tomato as affected by temperature and light[J]. Journal of Plant Physiology,1992,139(5):549-554 .
[20]刘黎军. 低温弱光对日光温室茄子生长及生理生化功能的影响[D]. 长春:吉林农业大学,2012.
[21]张强. 低温弱光对番茄形态和生理生化影响的研究[D]. 郑州:河南农业大学,2003:60.
[22]周艳虹,黄黎锋,喻景权. 持续低温弱光对黄瓜叶片气体交换、叶绿素荧光猝灭和吸收光能分配的影响[J]. 植物生理与分子生物学学报,2004,30(2):153-160.
