橡胶林土壤微生物碳代谢功能多样性及其变化
2017-03-10周玉杰李建华张广宇谭文丽王永鹏王春燕曹启民
周玉杰,李建华,张广宇,王 宁,谭文丽,王永鹏,王春燕,曹启民
(1. 海南省农垦科学院,海南 海口 570206;2. 海南省农垦农业科学研究所,海南 海口 570311)
土壤微生物是土壤的重要组成部分,是生态系统的分解者,它积极参与生态系统中的物质循环和能量流动,在维持生态系统的结构、功能方面起着极其重要的作用,研究土壤微生物群落功能多样性是理解生态系统过程的核心内容之一[1]。随着全球生态系统CO2气体浓度的持续升高,如何减少碳释放及增强固碳能力成为当前人们关注和研究的热点。而土壤是陆地生态系统中最大的碳库,同时也是最活跃的碳库之一,土壤微生物影响着土壤有机碳的转化,是整个生态系统碳循环的“关键”和“动力”[2],但两者之间的相互作用复杂而微妙,土壤微生物与土壤碳循环以及其他生态服务功能之间的关系及相互作用机制一直以来都是土壤生态学研究的难点和热点。开展土壤微生物碳代谢功能多样性研究,将有利于推进微生物介导的土壤生态系统乃至整个陆地生态系统碳循环机制及其他生态服务功能研究。
我国自1904年开始引种橡胶树以来,经过长期的艰苦奋斗,打破了橡胶树种植的传统禁区,形成了海南、云南、广东三大主要植胶区。橡胶树成为我国热带地区的主要经济作物,橡胶林生态系统已成为较大的人工生态系统之一,橡胶林对维持区域生态系统平衡发挥着重要的作用。但我国的第一代橡胶林基本上都是在自然植被的土地上建立起来的,橡胶树作为多年生乔木,在长期大规模替代自然植被背景下,随着时间的不断推移,是否对土壤微生物群落结构和功能产生影响?在实际研究工作当中,土壤生态系统中起关键作用的微生物往往被忽视。
土壤微生物对不同碳源利用能力的差异,是描述土壤微生物群落特征的一个重要指标,土壤微生物群落功能多样性可反映群落总体的动态变化。以往土壤微生物功能多样性的测定方法主要有磷脂脂肪酸法 (PLFA)[3]和基于PCR的分子生物学分析方法[4],但这两种方法技术要求高,耗费时间长,实验灵活性较差,又难以在较短时间内进行大量样品处理。基于碳代谢的 Biolog-ECO 技术为微生物群落功能多样性定量研究提供了一种灵敏度高、分辨性强、更加简便快捷的方法,该方法已广泛应用于不同土壤类型、不同植被物种下土壤及不同管理策略下的农业土壤微生物群落功能多样性研究中[5-8]。因此,我们以海南大丰农场6个不同林龄橡胶林样地为研究对象,运用Biolog-ECO技术对橡胶林土壤微生物群落碳代谢功能多样性进行研究,分析橡胶林土壤微生物群落碳代谢功能多样性在割胶前后的变化,以及对不同林龄橡胶林土壤微生物群落碳代谢功能多样性的差异,为橡胶林土壤生态系统健康提供理论基础。
1 材料与方法
1.1 研究区概况
本研究区域位于海南省琼中黎族苗族自治县大丰农场。该区域为热带海洋季风气候,年平均温度22.8℃,最冷月平均气温为 16~18℃,年平均降雨量2 278 mm。试验选取林龄5、10、13、18、25、30年常规栽培橡胶林作为试验样地,位于 19°14′~19°16′E、109°44′~109°45′N,海拔 175~207 m,每个样地面积 1 hm2。橡胶树栽培品系为PR107,株行距3 m×7 m。各样地的立地条件、栽培管理措施和割胶制度基本一致。
1.2 土壤样品采集
土壤样品采集时间为割胶前(5月)和割胶后(11月)。在每个林龄橡胶林样地内随机选取5个采样点,采集0~20 cm土层土壤,充分混合,迅速装入样品袋并置于冰块上,土样带回实验室后去掉植物根系和石块,放入4℃恒温冰箱内低温保存。
1.3 土壤微生物功能多样性测定
取样后48 h内采用含有31种碳源的Biolog-ECO微平板培养技术测定土壤微生物群落碳代谢功能多样性。具体方法为:称取新鲜土样5.00 g,加入45 mL无菌水后在摇床上振荡30 min,然后将土壤样品稀释至10-3,再从中取150 μL该悬浮液接种到Biolog-ECO 微平板的每一个孔中,1 h内测定第一次吸光值,最后将接种的微平板放在28℃的恒温培养箱中暗培养216 h,每隔24 h采用BIOLOG读数仪(BIOLOG,Hayward,USA)测定各孔在750 nm和590 nm 波长下的光吸收值。
1.4 数据分析
在一定培养时间内Biolog-ECO 微平板的孔数变化和每孔颜色变化的程度与土壤微生物群落功能密切相关。根据相关研究文献的方法[9],测试板孔中溶液吸光值平均颜色变化率(AWCD),计算方法为:

式中,C为每个培养基孔的光密度值,R为对照孔的光密度值,n为培养基碳源种类数,Biolog-ECO 微平板n为31。
采用培养至72 h的Shannon-Wiener指数表征土壤中微生物群落多样性,McIntosh指数表征土壤中微生物群落丰富度。
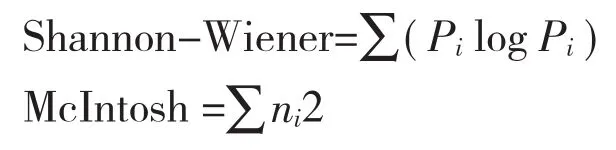
式中,Pi为第i孔的相对吸光值与所有反应孔相对吸光值总和的比值,即;ni为第i孔的相对吸光值(C-R)。
根据有关Biolog-ECO微平板培养时间对微生物相关数据分析的影响研究结果[10],以及本研究中各样地土壤微生物的AWCD增长曲线图,多样性指数选择AWCD值处于快速增长时(72 h)的数据进行分析。采用EXCEL2003计算土壤微生物AWCD值及多样性指数,DPS7.05进行方差分析,SPSS19.0进行主成分分析,Canoca4.5软件进行典范对应分析(CCA)。
2 结果与分析
2.1 土壤微生物AWCD值及变化
AWCD值可作为衡量微生物群落整体活性的有效指标,AWCD值的变化速率和最终达到值反映了土壤微生物群落利用碳源的能力。AWCD值越大,表明微生物的碳代谢活性越高,利用碳源的能力越强。培养0~24 h,各样地土壤微生物AWCD值变化均较小,培养24~72 h土壤微生物AWCD值快速增长,培养72 h后增长速度减慢并一直延续到144 h,培养144 h 后增长速率趋于平稳(图1)。
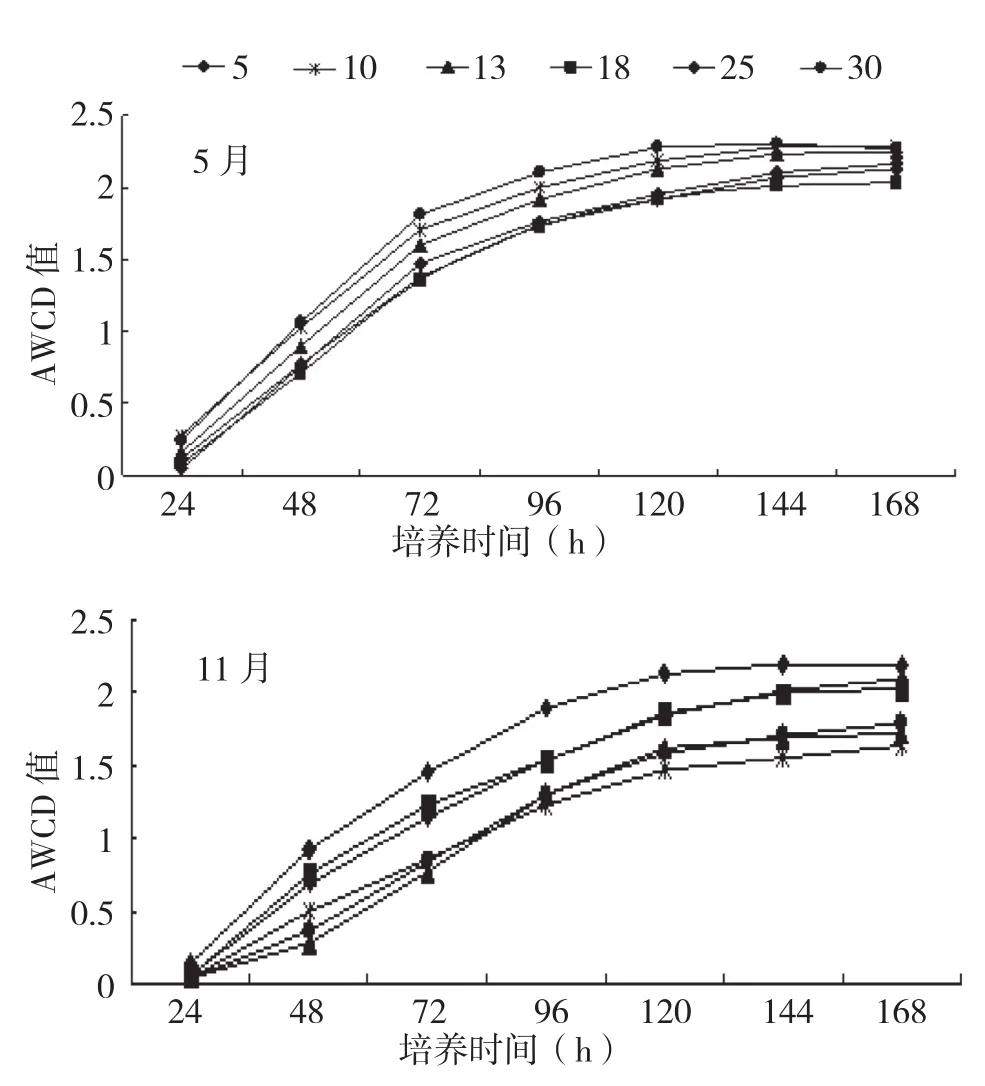
图1 橡胶林土壤微生物AWCD增长曲线
培养至72 h 的AWCD值,除林龄5年橡胶林割胶前后差异不明显外,林龄10、13、18、25、30年橡胶林5月份均高于11月份。不同林龄橡胶林AWCD值在割胶前后的变化不同。割胶前(5月)林龄10、30年橡胶林极显著高于林龄5、18、25年橡胶林,林龄13年橡胶林极显著高于林龄18年橡胶林,其他林龄橡胶林间差异不显著;而割胶后(11月)AWCD值以林龄5年橡胶林最高,其次为林龄8年和25年橡胶林,林龄10、13、30年橡胶林较低。其中林龄5年橡胶林显著高于林龄25年橡胶林,极显著高于林龄10、13、25、30年橡胶林;林龄18年和25年橡胶林极显著高于林龄10、13、30年橡胶林(图2)。

图2 不同林龄橡胶林土壤微生物AWCD值
2.2 土壤微生物碳代谢功能多样性指数
2.2.1 割胶前后土壤微生物群落功能多样性指数变化 割胶前林龄10、13、30年橡胶林Shannon-Wiener指数极显著高于割胶后,林龄5、18、25年橡胶林间割胶前与割胶后的Shannon-Wiener指数差异不明显。McIntosh指数除林龄5年橡胶林外,其他林龄橡胶林间割胶前McIntosh指数均极显著高于割胶后(图3)。

图3 不同林龄橡胶林土壤微生物Shannon-Wiener指数
2.2.2 不同林龄橡胶林土壤微生物群落功能多样性指数变化 割胶前Shannon-Wiener指数以林龄30年橡胶林最高,林龄18年橡胶林最低,林龄5、10、13、25年橡胶林间差异不显著。McIntosh指数以林龄30年橡胶林最高,林龄5年橡胶林最低,林龄10、13、18、25年橡胶林间差异不显著(图3、图4)。
割胶后Shannon-Wiener指数以林龄5年橡胶林最高,林龄13年橡胶林最低,林龄10、18、25、30年橡胶林间差异不显著。McIntosh指数以林龄5年橡胶林最高,其次为林龄18年和25年橡胶林,林龄10、13、30年橡胶林较低(图3、图4)。

图4 不同林龄橡胶林土壤微生物McIntosh指数
2.3 橡胶林土壤微生物对碳源的利用及变化
2.3.1 橡胶林土壤微生物对碳源的利用 对31种碳源的利用情况进行主成分分析,其中前3个主成分累积贡献率为86.077%,说明主成分1、2、3是变异的主要来源。3个主成分的特征值分别为15.649、7.663、1.976,贡献率分别为50.480%、4.720%和6.373%(图5)。

图5 橡胶林土壤微生物利用碳源的主成分分析
通过计算31种碳源的综合得分,可以看出橡胶林土壤微生物群落利用D-甘露醇、腐胺、苯乙胺、R-羟丁酸及衣康酸能力较强(综合得分>0.5),而对环式糊精、吐温80、甘氨酰-L-谷氨酸、α-D乳糖、肝糖、D-木糖和赤藓糖醇利用能力较差,综合得分低于平均水平(图6)。

图6 橡胶林土壤微生物利用31种碳源的综合得分
土壤微生物对不同类型碳源的利用能力程度,有助于更全面地了解土壤微生物群落功能特性[11-12]。将Biolog-ECO微平板上的31种碳分为糖及衍生物类(Carbohydrates)、氨基酸类(Amino acids)、羧酸类(Carboxylic acids)、酚酸类(Phenolic acids)、多聚物类(Polymers)和胺类(Amines)六大类型[13]。CCA分析表明,橡胶人工林土壤微生物群落对胺类、羧酸类、氨基酸类和糖及衍生物类碳源利用能力较强,而对酚酸类和多聚物类碳源的利用能力较差(图7)。
2.3.2 橡胶林土壤微生物利用碳源变化 通过主成分分析,计算橡胶林土壤微生物群落利用碳能力的综合得分,可以看出割胶前橡胶林土壤微生物群落利用碳的能力要高于割胶后。不同林龄橡胶林土壤微生物群落在割胶前后利用碳源的变化也不同,割胶前为林龄30年>10年>25年>13年>18年>5年,而割胶后为林龄5年>18年>25年>30年>13年>10年(图8)。
3 结论与讨论
3.1 橡胶林土壤微生物活性及利用碳源类型
在培养0~168 h内,橡胶林土壤微生物AWCD值随着培养时间延长而逐渐升高。这与其他学者的研究结果一致,即 AWCD 值随着培养时间的延续呈现出微生物常规生长曲线[14-15]。
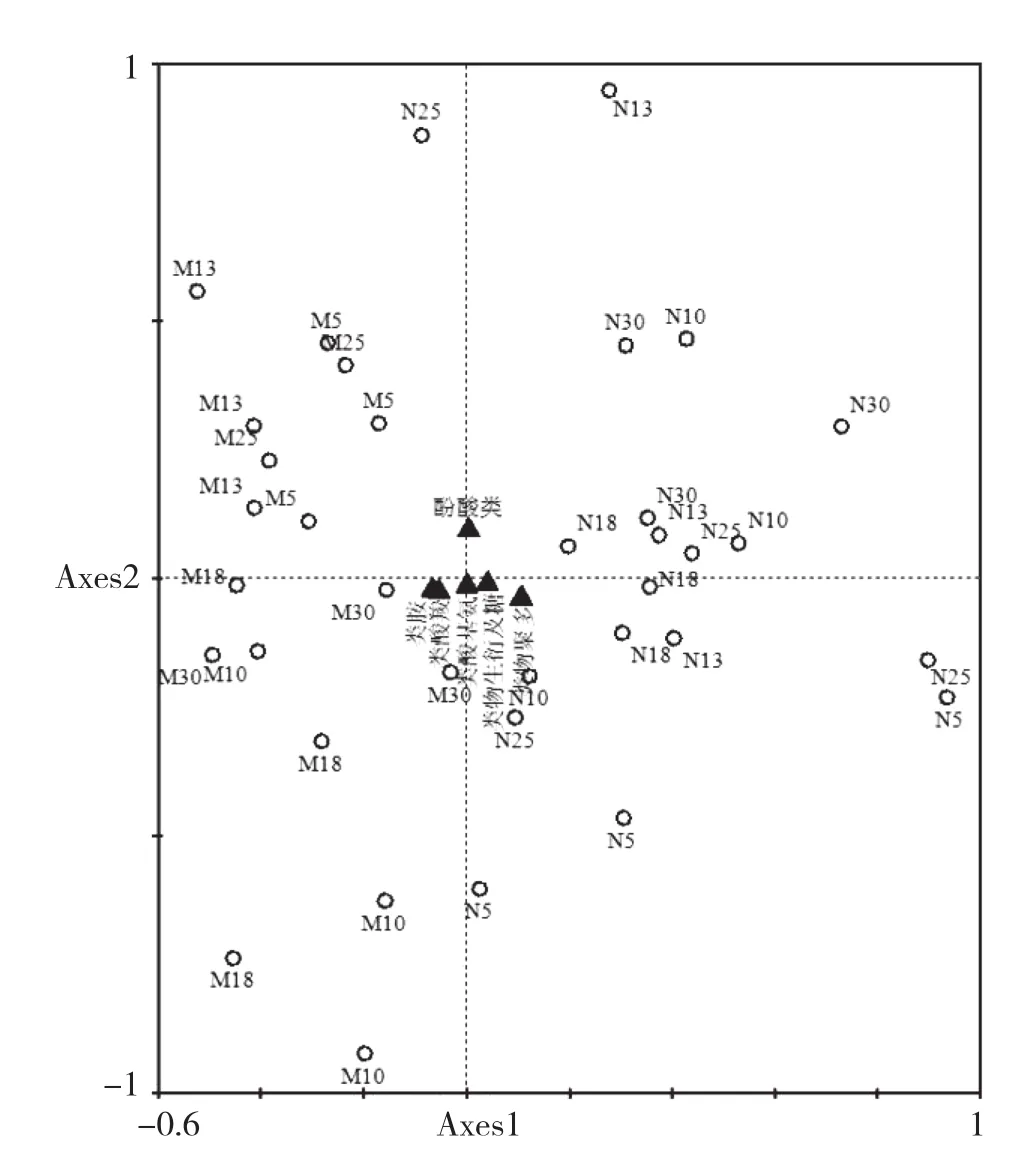
图7 橡胶林土壤微生物利用碳源类型CCA排序
橡胶林土壤微生物群落利用D-甘露醇、腐胺、苯乙胺、R-羟丁酸及衣康酸的能力较强,而利用环式糊精、吐温80、甘氨酰-L-谷氨酸、α-D乳糖、肝糖、D-木糖和赤藓糖醇的能力较差。总的来看,橡胶林土壤微生物群落对羧酸类、氨基酸类、胺类和糖及衍生物类利用能力较强,而对酚酸类和多聚物类的利用能力较差。这与其他天然次生林和人工林利用碳的类型基本一致,以往研究即得出土壤微生物利用率最高的是糖类、氨基酸类和羧酸[16-17],并认为糖类、氨基酸类和羧酸类是区分土壤微生物群落功能多样性最主要和最敏感的碳源[18]。

图8 橡胶林土壤微生物利用碳源的综合得分
3.2 割胶对橡胶林土壤微生物碳代谢功能多样性的影响
除林龄5年橡胶林外,林龄10、13、18、25、30年橡胶林割胶前的AWCD值均高于割胶后;割胶前Shannon-Wiener指数和McIntosh指数均以林龄30年橡胶林最高;割胶后Shannon-Wiener指数和McIntosh指数均以林龄5年橡胶林最高;不同林龄橡胶林土壤微生物群落利用碳能力割胶前为林龄30年>10年>25年>13年>18年>5年,而割胶后为林龄5年>18年>25年>30年>13年>10年。说明割胶前橡胶林土壤微生物AWCD值、多样性指数和碳源利用能力要高于割胶后,割胶导致处于胶乳生产期的林龄10、13、18、25、30年橡胶林土壤微生物碳代谢功能多样性下降,而林龄5年橡胶林由于尚未割胶,其土壤微生物功能多样性未受到割胶的影响,因此在割胶后林龄5年橡胶林的土壤微生物功能多样性要高于其他林龄橡胶林。割胶降低了土壤微生物群落的活性和碳代谢功能多样性,其可能与割胶致使土壤养分大量流失有关,以往研究即认为土壤微生物群落功能多样性与土壤养分密切相关[19-21]。
土壤微生物的活性和功能多样性降低是反映土壤生态系统质量和人类活动胁迫效应的重要生态指标[22]。健康土壤是橡胶树正常生长的保证,而土壤微生物多样性则是维持土壤健康和土壤质量的关键。因此,提高土壤微生物群落的活性和功能多样性,可以提高土壤肥力和结构,从而提高橡胶林的经济效益。建议根据不同林龄橡胶林的特点进行胶林土壤培肥改良,为土壤微生物创造良好的生长环境,以改善橡胶林土壤微生物群落的结构和功能,增强橡胶林的可持续生产能力;同时重视橡胶林土壤微生物功能多样性的研究,揭示橡胶人工林土壤生态系统“植物—土壤—微生物”三者之间的关系,为橡胶林生态系统健康以及土壤生态系统碳循环机制研究提供理论支撑。
[1]张其水,俞新妥. 杉木连栽林地营造混交林后土壤微生物的季节性动态研究[J]. 生态学报,1990,10(2):121-126.
[2]郑华,欧阳志云,方志国,等. Biolog-ECO在土壤微生物群落功能多样性研究中的应用[J].土壤学报,2004,41(3):456-461.
[3]宋风雅,梁鹏,何其光,等. 应用PCR-RFLP技术分析橡胶林土壤真菌种群动态变化[J]. 西南农业学报,2014,27(6):2469-2477.
[4]谭宏伟,杨尚东,吴俊,等. 红壤区桉树人工林与不同林分土壤微生物活性及细菌多样性的比较[J]. 土壤学报,2015,51(3):575-584.
[5]Garland J L,Mills A L. Classification and characterization of heterotrophic microbial communities on the basis of patterns of community-level sole-carbon source utilization[J]. Applied and environmental microbiology,1991,57(8):2351-2359.
[6]Zak J C,Willing M R,Moorhead D L,et al.Functional diversity of microbial communities:a quantitative approach[J]. Soil Biology and Biochemistry,1994,26(9):1101-1108.
[7]胡可,王利宾. BIOLOG微平板技术在土壤微生态研究中的应用[J]. 土壤通报,2007,38(4):819-821.
[8]董立国,蒋齐,蔡进军,等. 基于Biolog-ECO技术不同退耕年限苜蓿地土壤微生物功能多样性分析[J]. 干旱区研究,2011,28(4):630-637.
[9]邓晓,李勤奋,侯宪文,等. 香蕉枯萎病患病与健康蕉园土壤微生物群落功能多样性的比较研究[J]. 土壤通报,2013,44 (2):355-362.
[10]贾夏,董岁明,周春娟. 微生物生态研究中Biolog Eco微平板培养时间对分析结果的影响[J]. 应用基础与工程科学学报,2013,21(1):10-19.
[11]Choi K H.Comparison of two kinds of Biolog microplates(GN and ECO)in their ability to distinguish among aquatic microbial communities[J]. Journal of Microbiological Methods,1999,36(3):203-213.
[12]Preston-Mafham J,Boddy L,Randerson P F. Analysis of microbial community functional diversity using sole-carbon-source utilization profiles-a critique[J]. Fems Microbiology Ecology,2002,42(1):1-14.
[13]孔滨,杨秀娟. Biolog生态板的应用原理及碳源构成[J]. 绿色科技,2011(7):231-234.
[14]王洪媛,管华诗,江晓路. 微生物生态学中分子生物学方法及T-RFLP技术研究[J]. 中国生物工程杂志,2004,24(8):42-47.
[15]范瑞英,杨小燕,王恩姮,等.黑土区不同林龄落叶松人工林土壤微生物群落功能多样性的对比研究[J]. 北京林业大学学报,2013,35(2):63-68.
[16]胡婵娟,傅伯杰,刘国华,等. 黄土丘陵沟壑区典型人工林下土壤微生物功能多样性[J]. 生态学报,2009,29(2):727-733.
[17]李小容,韦金玉,陈云,等. 海南岛不同林龄的木麻黄林地土壤微生物的功能多样性[J]. 植物生态学报,2014,38(6):608-618.
[18]郑华,陈法霖,欧阳志云,等. 不同森林土壤微生物群落对Biolog-GN板碳源的利用[J]. 环境科学,2007,28(5):1126-1130.
[19]韩世忠,高人,李爱萍,等. 中亚热带地区两种森林植被类型土壤微生物群落结构[J]. 应用生态学报,2015,26(7):2151-2158.
[20]徐文娴,赵从举,朱敏捷,等. 海南岛桉树人工林土壤微生物功能多样性特征[J]. 广东农业科学,2016,43(7):65-72.
[21]字洪标,刘敏,阿的鲁骥,等. 三江源区不同建植年限对人工草地土壤微生物功能多样性的影响. 生态学杂志,2017,36( 4) :978-987.
[22]李智卫,王超,陈伟,等. 不同树龄苹果园土壤微生物生态特征研究[J]. 土壤通报,2011,42(2):302-306.
