闽楠不同种源苗期生长规律研究
2017-03-10谭文婧林建勇刘雄盛梁瑞龙
李 娟,谭文婧,林建勇,刘雄盛,梁瑞龙
(1.广西壮族自治区林业科学研究院/广西优良用材林资源培育重点实验室/国家林业局中南速生材繁育实验室,广西 南宁 530002;2.广西壮族自治区融水县国营贝江河林场,广西 柳州 545300)
闽楠(Phoebe bournei)为樟科常绿大乔木,是我国特有的珍贵用材树种和优良观赏树种[1-2],被列为国家Ⅱ级珍稀濒危植物。闽楠材质结构细密、耐腐性能好、经久耐用,是高档家具、精致器具、上等建筑及棺木良材,为商品材“金丝楠”最主要产材树种。该树种自然分布于福建、江西、湖南、广西、广东、贵州、浙江、湖北、四川等地[1],在广西境内主要产于北部的桂林、贺州、柳州,南宁市的大明山亦有自然生长。
目前对闽楠的相关研究主要涉及野外资源调查[3-4]、群落生态[5-7]、生理特性[8-13]、育苗和造林[14-19]、遗传多样性[20-22]等方面。有关闽楠遗传改良方面的研究,以苗期生长规律为主,部分学者也对闽楠幼龄林生长性状的遗传变异规律做了相关的研究分析[23-25]。但是这些研究的参试种源仅来自于福建、江西两个省份,种源选择范围较小。我们选择闽楠天然分布较多的6个省份11个代表种源进行苗期比较试验,揭示不同种源闽楠一年生苗木在我国闽楠分布区南缘广西的生长规律,以期为科学培育闽楠苗木和选育适合广西栽培的优良种源提供参考。
1 材料与方法
1.1 试验地概况
试验在广西柳州市融水县国营贝江河林场苗圃基地进行,试验地位于109°14′E、25°04′N,属中亚热带季风气候,由于海拔较高、山地较多,故山区气候特征比较明显。年平均温度19.6℃,年均降雨量1 757.5 mm,降水主要集中在4~8月;全年平均总日照时数1 699.0 h。
1.2 试验材料
供试11个闽楠种源采种点的地理气候信息见表1。2014年11月下旬至12月上旬,每个种源选择10~15株健壮母树采种,母树间距50 m以上。果实采回后除去果肉,洗净阴干后,等量混合,充分混匀后作为该种源的供试材料。于2016年2月17日播种于沙床催芽,4月22~23日将露白的种子点入直径7.5 cm、高12 cm的无纺布育苗袋,育苗基质为97%经过发酵的菌渣+3%黄心土混合基质。苗期管理与常规容器育苗管理相同,夏季搭上遮光率为80%的遮阳网遮阳。

表1 闽楠各种源采种点的地理位置与气候因子
1.3 指标测定
2016年5月23 日,每个种源随机选择100株挂牌并测定苗高,由于5月闽楠幼苗茎幼嫩,不适合用游标卡尺测量,因此本次试验并未测量地径。此后每个月23日测定一次苗高、地径,至12月下旬苗木基本停止生长为止。
1.4 数据处理和统计分析
1.4.1 生长模型的建立 采用Logistic方程对苗高和地径进行拟合,其方程为:
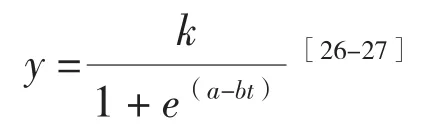
式中,y为苗木生长量,t为生长时间,a和b为待定系数,k为既定条件下苗木生长极限值。
1.4.2 有序聚类分析 由于苗木的生长过程是连续的,因此在不打乱样本顺序的前提下采用有序样本聚类分析法[26]对苗高和地径不同生长阶段进行划分,主要步骤如下:
(1)按照直径计算公式:

(2)计算最小误差函数值:
E[p0 (n,k)] = min {e[p(j-1 )(k-1)]+D(j,n)}
试验数据采用DPS 9.0进行处理,采用Excel 2007进行统计,采用SPSS 18.0软件进行相关分析和聚类分析。
2 结果与分析
2.1 闽楠不同种源苗高年生长节律比较

图1 闽楠不同种源苗高累积生长量曲线
从图1可以看出,闽楠11个种源幼苗苗高从4月下旬播种至6月下旬生长较为缓慢,7月下旬至10月下旬苗高生长速度较快,10月之后苗高增长缓慢,苗高快速增长的时间持续约3个月。闽楠苗高年生长过程呈现出“慢-快-慢”的“S”型生长趋势,且苗高在整个生长季节都持续增长,属于全期生长类型。对不同种源闽楠进行比较发现,6月下旬以前各个种源的苗高生长差异很小;从7月初开始到10月下旬,不同种源间高生长差异逐渐拉大,生长最快的广西富川种源和生长最慢的湖南沅陵种源相差37.47 cm,前者约为后者的3倍。此后,虽然广西资源、贵州思南、江西井冈山种源高增长略快于其他种源,但是各种源的生长差异趋于稳定。

图2 闽楠不同种源苗高净生长量曲线
从图2可以看出,闽楠各种源苗高生长呈现两次高峰,第1次生长高峰均在5月23日,之后生长暂时放缓,大部分种源从7月下旬开始进入快速生长期,苗高增长量逐渐大于5月峰值,直至9月23日达到第2次生长高峰,湖南永顺和湖南沅陵种源则在10月23日达到生长高峰。此外,湖南沅陵种源不同峰值之间的差别不大,尤其是9月23日和10月23日的苗高净增生长量只相差0.4 cm,这可能与种源遗传特性和生态适应性有关。总之,闽楠幼苗高生长具有明显的间歇性。
2.2 闽楠不同种源苗高生长周期比较
采用有序样本聚类分析法,结合苗木生长特性,闽楠11个种源苗高生长均可划分为出苗期、生长初期、速生期和生长后期4个阶段。各个阶段的起止时间和生长量占生长总量的比例见表2。从图2和表2可以看出,各种源闽楠的生长周期基本一致,江西井冈山种源进入速生期时间是8月24日、持续92 d,其余10个种源速生期从7月24日至10月23日共91 d、占观察时间的37.30%。各种源的速生期苗高净增长量明显高于其他阶段,均在55%以上。速生期净生长量占总生长量比例最高的为湖北来凤种源,达71.70%;其次是广西富川、贵州思南,分别为69.89%、67.42%;最低的是湖南永顺(55.73%)。除江西井冈山种源外的10个种源速生期持续时间约为生长前期的1.5倍,而净生长量则为生长前期的4.2~6.6倍。
江西井冈山种源的生长初期时间较其他种源多30 d,因而进入速生期的时间较其他种源晚,速生期持续时间和其他种源基本相同,生长后期的时间则较其他种源少30 d。江西井冈山种源生长前期持续时间与速生期相同,但是速生期的净生长量约为生长前期的3倍,这可能与其自身的遗传特性有关。
2.3 闽楠不同种源地径生长节律比较
从闽楠各种源地径生长过程(图3)可知,各种源地径生长趋势比较一致,均未明显表现出“慢-快-慢”的趋势。各种源间地径的生长差异在6月23日之后逐渐拉大,至生长停止时湖南沅陵种源地径显著小于其他种源,比最粗的贵州思南种源小1.95 mm,前者仅为后者的66.26%。除广西富川、贵州思南、湖南沅陵种源外,其余8个种源间地径差异较小。图4表明,闽楠各种源幼苗在年生长期内有1~2次生长高峰,除广西富川和福建南平种源外,其余种源的第1次地径生长高峰均出现在8月23日。各种源间第2次生长高峰出现的时间则有所差别,有的种源出现在10月23日,有的则在11月23日。广西富川和福建南平种源在生长期间仅于11月23日出现1次明显的生长高峰,在此之前地径增长较平缓。

图3 闽楠不同种源地径累积生长量曲线
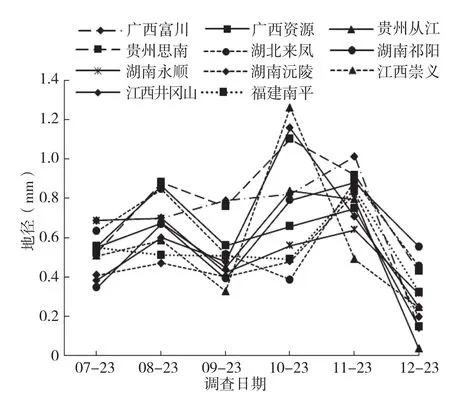
图4 闽楠不同种源地径净生长量曲线
2.4 闽楠不同种源地径生长周期比较
由表3可知,11个种源闽楠地径生长均可划分为出苗期、生长初期、速生期和生长后期4个阶段。全部种源的速生期均介于8月24日至11月23日,共92 d,占整个观察期的37.70%。除江西井冈山种源地径与苗高同时进入速生期外,其余种源地径进入速生期的时间较苗高晚31 d。在苗高生长放缓后,地径仍保持较高
的增长水平。从表3可以看出,各个种源的地径净生长量在出苗期和生长初期这两个阶段所占比例均在20%~30%,生长后期所占比例除湖南祁阳种源外均在10%以下;各种源地径在速生期持续时间约为生长初期的1.5倍,净生长量是生长初期的1.2~2.4倍,占总生长量的34.76%~48.61%,高于其他几个生长阶段。然而,地径在速生期的增长量与前两个生长阶段的差距相对苗高的要小。

表2 闽楠不同种源苗高生长阶段划分

表3 闽楠不同种源地径生长阶段划分
2.5 闽楠不同种源苗期生长模型的建立和拟合
用Logistic模型拟合不同种源闽楠幼苗苗高和地径所测得数据,建立闽楠苗高和地径生长模型。方程拟合结果(表4)显示,各种源闽楠苗高拟合方程的决定系数R2在0.967~0.994,各种源地径的决定系数R2在0.981~0.996,表明各种源闽楠获得的苗高和地径拟合曲线拟合度较好,Logistic曲线能较好地描述不同种源闽楠苗高和地径的生长趋势。

表4 闽楠不同种源苗高、地径年生长动态Logistic方程拟合结果
2.6 闽楠不同种源苗期生长量比较
不同种源间苗高和地径存在显著差异,多重比较分析结果(表5)表明,一年生苗高生长以广西富川种源表现最优,显著大于其他种源;其次为广西资源;再次为贵州思南和湖北来凤;生长最差的是湖南沅陵种源。1年生苗高累积生长量最大的广西富川种源是最小的湖南沅陵种源的2.46倍。地径生长表现最佳的种源是贵州思南和广西富川,其次为广西资源、湖北来凤和湖南祁阳;生长最差的是湖南沅陵种源。地径累积生长量最大的贵州思南种源是最小的湖南沅陵种源的1.41倍。
2.7 聚类分析
由于闽楠各种源1年生苗的苗高和地径存在显著差异,以苗高和地径为指标,对11个种源进行聚类分析,结果(图5)表明可将11个种源划分为3类:第1类为速生种源,包括广西富川和广西资源,这类种源生长长势最好,表现最优;第2类为中生种源,包括贵州从江、贵州思南、湖北来凤、湖南祁阳、湖南永顺、江西崇义、江西井冈山、福建南平,这类种源长势稍差于第一类;第3类为慢生种源,包括湖南沅陵,这类种源的苗高和地径长势最差。

表5 闽楠不同种源苗高、地径LSD多重比较分析结果
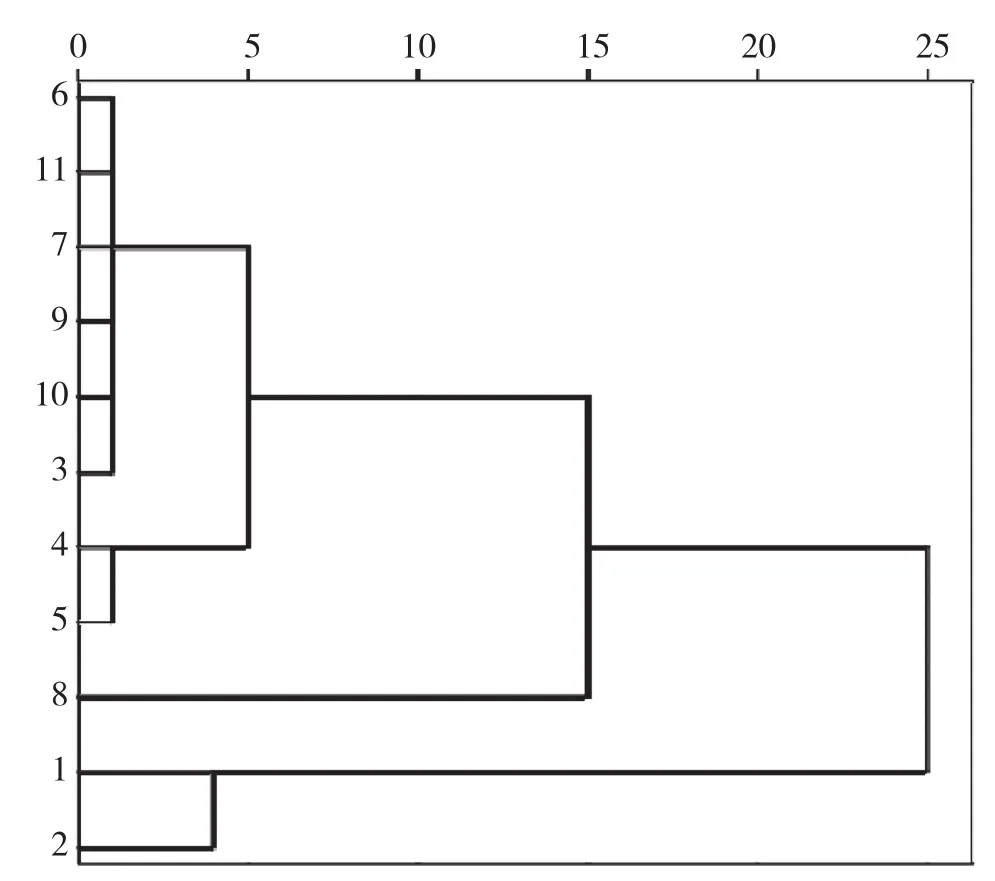
图5 11个闽楠种源聚类分析
3 结论与讨论
在本试验中,我们利用Logistic方程拟合11个闽楠种源的苗高、地径,结果表明决定系数在0.967~0.996之间,拟合效果良好,能够用于描述闽楠苗高、地径的生长规律,有助于预测最佳的出圃时间。11个种源的苗高生长节律基本一致,呈现出“慢-快-慢”的“S”型趋势。闽楠苗高生长高峰期出现在5月和9月,个别种源的第2次生长高峰期出现在10月,这与江香梅等[25]的研究结果有所不同。地径生长则并未表现出典型的“S”型生长规律,地径的两次生长高峰期均比苗高晚1个月。闽楠幼苗这种异速生长现象可能是苗木为了能够获得更多阳光和占据更有利的生存空间,在生长初期将更多的营养物质投入高生长。
采用有序聚类分析法,结合苗木的生长特性,闽楠11个种源苗高和地径生长均可划分为出苗期、生长初期、速生期和生长后期4个阶段,各个种源在各生长阶段都表现出相似的生长规律。地径的出苗期较苗高长1个月,因此进入后面3个生长阶段的时间也较苗高晚1个月。苗高速生期约占整个观测期的37%,但却完成了年生长量55%以上的生长量。地径速生期持续时间和苗高的时间一致,速生期生长量占年生长量的34%~48%。这说明闽楠速生期的生长量决定着整个生长期的生长量,因此在速生期要加强水肥管理,合理调节水肥光热条件,促进苗木快速生长并争取延长幼苗的速生期。
方差分析结果表明,11个闽楠种源的苗高和地径存在显著差异,说明种源选择对闽楠遗传改良有重要意义。苗高表现最好的是广西富川种源,其次为广西资源种源,生长最差的是湖南沅陵种源。地径生长表现最佳的是贵州思南和广西富川种源,其次为广西资源、湖北来凤和湖南祁阳种源,生长最差的是湖南沅陵种源。对11个闽楠种源的苗高和地径进行聚类分析,可将它们分成3类:第1类为速生种源,包括广西富川和广西资源;第2类为中生种源,包括贵州从江、贵州思南、湖北来凤、湖南祁阳、湖南永顺、江西崇义、江西井冈山、福建南平;第3类为慢生种源,包括湖南沅陵种源。广西的两个种源表现较佳,可能是因为广西种源能较好地适应本地气候特点。本研究主要针对幼苗阶段开展试验,闽楠苗期的苗高和地径的生长速率还不稳定,因此研究结果只能作为优良种源早期选择的参考,而各种源间的生长差异还有待进一步的深入研究。
[1]梁盛业. 广西树木志[M]. 北京:中国林业出版社,2012:242-243.
[2]中国植物志编辑委员会. 中国植物志[M]. 北京:科学出版社,1982:112.
[3]王通,吴大荣,徐建明,等. 车八岭闽楠种群的现状及保护对策[J]. 华南农业大学学报,2000,21(1):72-74.
[4]邹秀红. 福建永春闽楠天然林植物区系和物种多样性研究[J]. 亚热带植物科学,2002,31(3):23-26.
[5]刘宝,陈存及,陈世品,等. 闽楠群落优势种群结构与空间分布格局[J]. 福建林学院学报,2006,26(3):210-213.
[6]吴大荣,朱政德. 福建省罗卜岩自然保护区闽楠种群结构和空间分布格局初步研究[J]. 林业科学,2003,39(1):23-30.
[7]吴大荣. 罗卜岩保护区闽楠等优势植物种群竞争研究初步[J]. 南京林业大学学报,1998,22(3):38-41.
[8]颜珣,文仕知,郭文平,等. 施肥处理在高温季节对闽楠幼林叶绿素荧光特性的影响[J]. 中南林业科技大学学报,2015,35(8):73-76.
[9]葛永金,刘跃钧,高伟,等. 不同光照强度下楠木属3个树种苗木的形态响应与适应[J]. 江西农业大学学报,2014,36(1):109-114.
[10]安常蓉,韦小丽,叶嘉俊,等. 温湿度交互作用对闽楠幼苗形态和生理生化的影响[J]. 西北林学院学报,2015,30(5):20-27.
[11]刘宝,陈存及,林达定,等. 21个闽楠种源叶片光合色素含量及叶绿素荧光参数分析[J]. 江西农业大学学报,2014,36(1):115-121.
[12]盛杰,陈月华,吴际友,等. 闽楠家系苗期光合特性的研究[J]. 中南林业科技大学学报,2015,35(6):45-49.
[13]姜顺邦,张怡,韦小丽,等. 不同闽楠优树子代苗期生长及光合生理特性比较[J]. 浙江农林大学学报,2016,33(1):51-59.
[14]李娟,梁瑞龙,姜英. 闽楠优质苗培育技术[J].广西林业科学,2015,44(3):308-310.
[15]楚秀丽,刘青华,范辉华,等. 不同生境、造林模式闽楠人工林生长及林分分化[J]. 林业科学研究,2014,27(4):445-453.
[16]欧建德. 福建闽楠人工幼林氮磷钾施肥效应与施肥模式[J]. 浙江农林大学学报,2015(1):92-97.
[17]王东光,尹光天,杨锦昌,等. 磷肥对闽楠苗木生长及叶片氮磷钾浓度的影响[J]. 南京林业大学学报(自然科学版),2014,38(3):40-44.
[18]吴君,吴冬,楼雄珍. 不同基质配比及复合肥处理对3年生楠木容器苗生长的影响[J]. 西部林业科学,2015,44(1):109-113,120.
[19]邵明晓,文仕知,何功秀,等. 不同林龄闽楠人工林生物量结构特征[J]. 中南林业科技大学学报(自然科学版),2014,34(6):44-48.
[20]江香梅,温强,叶金山,等. 闽楠天然种群遗传多样性的RAPD分析[J]. 生态学报,2009,29(1):438-444.
[21]周生财,黄华宏,童再康,等. 4种楠木AFLP反应体系优化建立[J]. 浙江农林大学学报,2013,30(5):789-796.
[22]田晓俊,温强,汪信东,等. 闽楠、红楠AFLP反应体系建立[J]. 林业科技开发,2009(3):38-42.
[23]欧建德. 闽楠优良家系和单株的早期综合选择研究[J]. 西南林业大学学报,2015(1):33-37.
[24]罗宁. 闽楠优良地理种源选择及其子代林遗传增益分析[J]. 西部林业科学,2014,43(5):19-23,40.
[25]江香梅,肖复明,叶金山,等. 闽楠种源苗期生长性状地理变异及遗传参数估算[J]. 江西农业大学学报,2008,30(4):666-670.
[26]李秋荔,黄寿先,李志先,等. 中国马褂木不同种源苗期生长规律研究[J]. 广西植物,2012,32(3):355-361.
[27]杨志玲,杨旭,谭梓峰,等. 厚朴不同种源苗期生长模型的拟合[J]. 西北农林科技大学学报(自然科学版),2011,39(4):60-68.
