高脂高糖饮食诱导的非酒精性脂肪性肝炎大鼠肝组织TLR4和MyD88 mRNA的表达
2017-03-02向晶陈海君施军平
向晶 陈海君 施军平

[摘要] 目的 探讨Toll样受体4(TLR4)、髓样分化因子88(MyD88)在高脂高糖饮食诱导的非酒精性脂肪性肝病(NAFLD)大鼠肝组织中的表达及其意义。 方法 选取SD大鼠20只,随机分为两组,模型组SD大鼠给予高脂高糖饲料喂养,正常组给予普通饲料喂养,10周末分别进行如下检测:①计算肝指数;②HE染色,光镜下观察各组大鼠肝组织的病理改变;③应用RT-PCR法检测大鼠肝组织TLR4、MyD88 mRNA的表达。 结果 模型组的肝指数较正常组明显增加(P<0.01);模型组的NAFLD活动度计分与正常组比较差异有统计学意义(P<0.01);模型组大鼠肝组织TLR4和MyD88的mRNA表达水平与正常组比较差异有统计学意义(P<0.01)。 结论 持续10周的高脂高糖饮食喂养可以诱导NAFLD大鼠模型,模型组大鼠肝组织TLR4 mRNA和MyD88 mRNA表达升高,在肝细胞炎症改变中起重要作用。
[关键词] 非酒精性脂肪肝;大鼠;Toll样受体4;髓样分化因子88
[中图分类号] R575.1 [文献标识码] A [文章编号] 1673-9701(2016)30-0024-05
近年来非酒精性脂肪性肝病(nonalcoholic fatty liver disease,NAFLD)的发病率逐渐升高,且呈年轻化趋势。胡卫等[1]对武汉大学医学院教职工体检的调查研究发现,NAFLD总患病率达到20.7%,其具体发病机制尚未完全明确,但大量研究表明胰岛素抵抗、炎症反应、脂质代谢异常在该病的发生发展中起着重要作用[2]。目前研究表明,Toll样受体(Toll like receptor,TLR)与感染信号的识别及转导和机体的损伤密切相关,将感染和炎症、组织损伤联系起来[3]。Toll样受体4(Toll like receptor 4,TLR4)是内毒素识别受体,目前已知TLR4介导的信号转导包括髓样分化因子88(myeloid differentiation factor 88,MyD88)依赖性和非依赖性途径。MyD88依赖性途径主要介导核因子-κB(nuclear fator-κB,NF-κB)活化、细胞因子的产生[4]。本文通过研究TLR4、MyD88在高脂高糖饮食诱导的非酒精性脂肪性肝炎(non-alcoholic steatohepatitis,NASH)大鼠肝组织中的表达,旨在探讨TLR4、MyD88在NASH炎症反应中的作用。
1 材料与方法
1.1 非酒精性脂肪性肝炎动物模型的建立
健康雄性Spraque-Dawley大鼠20只,体重180~225 g,清洁级,购买于上海斯莱克实验动物有限公司[许可证号:SCXK(沪) 2008-0016],饲养于浙江中医药大学动物实验中心。20只SD大鼠随机分为正常组(予普通饲料喂养)10只及模型组10只(予以高脂高糖饲料喂养);于第10周处死正常组10只大鼠及模型组10只大鼠,HE染色,光镜下观察正常组和模型组大鼠肝组织的病理改变,以证实大鼠非酒精性脂肪性肝炎(non-alcoholic steatohepatitis,NASH)模型成功建立。实验动物每笼5只,分笼饲养,均自由饮水和进食,(20±2)℃明暗各12 h,均连续10周。
1.2 标本采集
第10周末当晚两组大鼠禁食不禁水,次日晨空腹腹腔注射麻醉,心脏采血用于测定生化指标和细胞因子;分离整个肝脏称重,在肝右叶切取数块肝组织,装入冻存管,投入液氮中保存,进入实验室后移入-80℃冰箱中保存,用于提取总RNA,以检测肝组织中TLR4、MyD88的mRNA表達。
1.3 病理组织学检查
常规HE染色,光镜下观察大鼠肝细胞脂肪变性程度、气球样变性、小叶内炎症程度以评定NAFLD活动度计分(NAFLD activity score,NAS),脂肪肝病理组织学诊断参考中华医学会肝脏病学分会脂肪肝和酒精性肝病学组“非酒精性脂肪肝诊断标准”[5]。根据肝小叶内脂肪变的肝细胞数将脂变程度量化:0分(肝小叶内<5%肝细胞脂肪变)、1分(肝小叶内5%~33%肝细胞脂肪变)、2分(肝小叶内34%~66%肝细胞脂肪变)、3分(肝小叶内>66%肝细胞脂肪变)。肝小叶内炎症(20倍镜计数坏死灶)计分:0分(无坏死灶)、1分(<2个坏死灶)、2分(2~4个坏死灶)、3分(>4个坏死灶)。肝细胞气球样变程度计分:0分(无气球样变)、1分(少见气球样变)、2分(多见气球样变)。NAFLD活动度计分(NAFLD activity score,NAS)为肝细胞脂肪变+小叶内炎症+肝细胞气球样变,NAS小于3分则不考虑NASH,NAS大于4分则可诊断NASH,介于两者之间为NASH可能。
1.4 RT-PCR检测TLR4、MyD88 mRNA在NASH大鼠肝组织中的表达
1.4.1 总RNA提取 从超低温冰箱中取出冰冻的组织,约为100 mg,迅速转移至用液氮预冷的研钵中,用研杵研磨组织,直至研磨成粉末状,向研钵中加入2 mL RNAiso Plus,完全覆盖研磨成粉末状的样品,静置室温中,待样品完全融化,再用研杵研磨至裂解液呈透明状,将匀浆液转移至2个1.5 mL无RNA酶和无DNA酶的离心管中。离心之后,吸取上清液,移入新的1.5 mL无RNA酶和无DNA酶的离心管中,向新的匀浆裂解液中加入0.2 mL氯仿,用手剧烈震荡15 s,12 000 r 4℃离心15 min,吸取上清液转移至另一新的离心管中,加入等体积的异丙醇,上下颠倒离心管充分混匀后,在室温下静置10 min,再次12 000 r 4℃离心10 min,弃去上清,加入75%的乙醇1 mL洗涤并干燥。最后用40~50 μL的DEPC处理水溶解RNA。提取的总RNA经紫外分光光度计测定提取物的浓度,OD260/280比值在1.8~2.0之间。
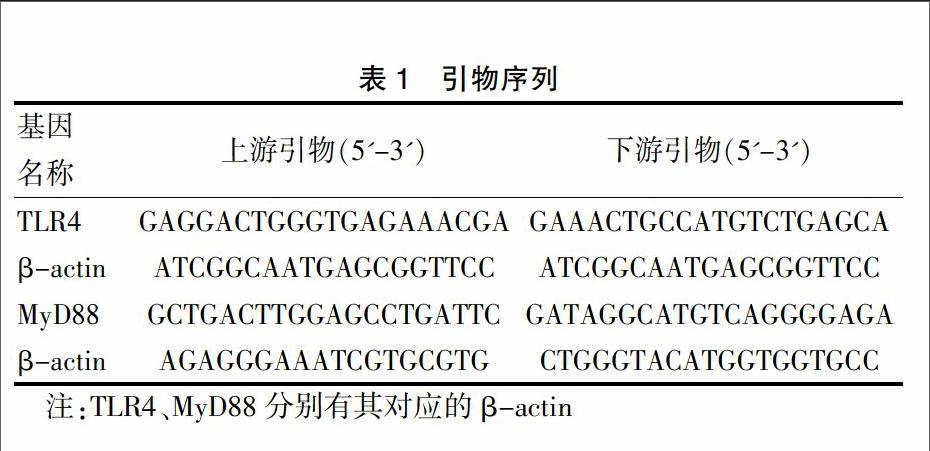
1.4.2 合成相关引物 从有关资料获得所需引物,并在基因库(Genebank)中进行核对。引物序列见表1,由上海Invitrogen 公司合成。
1.4.3 逆转录反应 用上述细胞中提取的2 μL总RNA加入0.2 mL离心管中,于70℃温育10 min,短暂离心后置于冰上。依据如下顺序加入以下试剂以建立一个20 μL的反应体系:MgCl2(25 mM)4 μL ,反转录10×缓冲液2 μL,dNTP混合物(10 mM)2 μL,重组的RNasin核糖核酸酶抑制剂0.5 μL,AMV反转录酶0.6 μL,随机引物1 μL,无核酸酶的水7.9 μL,总RNA 2 μL。将反应体系于室温温育10 min,于42℃温育60 min,于95℃加热5 min,再于4℃放置5 min。即为cDNA,放入-30℃存放。
1.4.4 PCR反应 取上述逆转录反应体系于18 μL反应体系中进行PCR反应。反应体系组成如下:2×Taq PCR Master Mix 9 μL,TLR4、MyD88上游引物 2 μL,TLR4、MyD88下游引物 2 μL,β-actin上游引物 2 μL,β-actin下游引物 2 μL,去离子水1 μL。变性94℃ 30 sec,退火58℃ 30 sec,延伸72℃ 30 sec,4℃冷却5 min结束反应。按上述条件循环25次,最后2%琼脂糖凝胶电泳20 min。应用BIO-RAD图像分析系统进行吸光度扫描,计算TLR4/β-actin及MyD88/β-actin的比值。
1.5 统计学处理
采用SPSS17.0统计软件包进行统计,计量资料以均数±标准差(x±s)表示,采用单因素方差分析进行统计分析,P<0.05表示差异有统计学意义,P<0.01表示差异有高度统计学意义。
2 结果
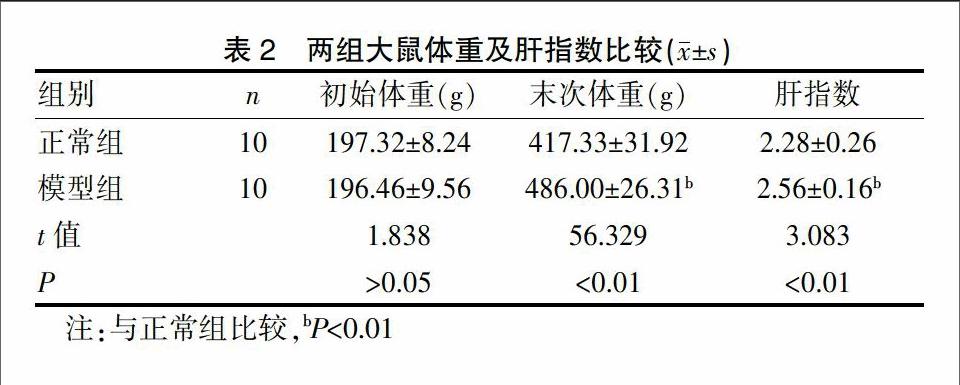
2.1 两组大鼠体重及肝指数比较
实验期间正常组大鼠灵活,精力充沛,皮肤有光泽,体重逐渐增加;模型组大鼠造模时体重较正常组逐渐增加,粪便粘臭,毛发失去光泽,渐油腻,实验初期活动与正常组无异,第5周后大鼠活动明显减少,体态笨重。由表2可见模型组的肝指数较正常组明显增加(P<0.01)。
2.2 两组大鼠病理组织学变化及NAFLD活动度计分比较
通过光镜下观察发现:正常组大鼠肝脏组织肝小叶结构完整;肝细胞呈多边形围绕中央静脉呈放射状分布,大小较一致,核结构清晰可见,肝窦清晰可见。模型组大鼠肝细胞脂肪变性明显,主要以小泡性为主,偶尔可见重度脂肪变性,小叶内可见淋巴细胞浸润形成的点灶状坏死,见图1。模型组的NAFLD活动度计分(NAFLD activity scores,NAS)与正常组比较有显著差异(P<0.01),证实NASH模型成功建立,见表3。
2.3 两组大鼠肝组织TLR4和MyD88 mRNA的表达比较
用RT-PCR法分别检测NASH大鼠肝组织TLR4和MyD88 mRNA的表达,TLR4和β-actin的RT-PCR产物电泳结果显示扩增片段为266 bp和149 bp,与预期设计的目的片段大小相同;MyD88和β-actin的RT-PCR产物电泳结果显示扩增片段为191 bp和309 bp,与预期设计的目的片段大小相同,见图2。模型组大鼠肝组织TLR4和MyD88 mRNA的表达水平与正常组比较有显著差异(P<0.01),见表4。
3讨论
NAFLD是指除大量饮酒、遗传疾病、成脂药物影响等导致的肝脂肪病,影像或者病理证实存在肝脏脂肪变的一种代谢性疾病[6,7],包括非酒精性单纯性脂肪肝(non-alcoholic simple fatty liver,NAFL)、非酒精性脂肪性肝炎(NASH)、脂肪肝相关肝硬化。随着肥胖及其相关代谢综合征全球化的流行趋势,NAFLD现已成为欧美等发达国家和我国富裕地区慢性肝病的重要病因。研究发现,在NAFLD漫长的病程中,NAFLD患者肝脏一般呈静止状态,随访10~20年肝硬化发生率仅0.6%~3%,而NASH患者10年内肝硬化发生率为15%~25%[8],NASH为NAFLD发生肝硬化的限速步骤,因此,NAFLD引起了临床医师极大关注,其防治研究成为医学研究的热门课题之一。
目前其发病机制尚不十分清楚,以1998年Day CP[9]提出的“二次打击学说”最为著名,即各种原因如肥胖、2型糖尿病、脂代谢紊乱等导致的胰岛素抵抗(insulin resistance,IR),游离脂肪酸增加,肝脏脂肪代谢障碍,从而使肝细胞内合成三酰甘油增加而输出减少,导致过量的肝脏脂质聚积,并使肝脏易受第二次打击是“二次打击学说”的第一步;线粒体脂肪酸氧化引起的氧化应激,核因子-κB(nuclear fator-κB,NF-κB)依赖的炎症细胞因子表达和脂肪细胞因子是所有的被认为是导致第二次打击产生肝细胞损伤、炎症和纤维化的潜在因素[10,11]。
本研究組成员蒋艳明等[12]利用高脂高糖饮食喂养建立NAFLD模型,已证明其可靠性,本实验亦采用高脂高糖喂养诱导NAFLD大鼠模型,大鼠肝脏HE染色显示肝脏脂肪变性程度加深、脂滴沉积,并可见炎症细胞浸润,具有典型NAFLD病理表现,并检测模型组大鼠肝组织中TLR4和MyD88的表达。Toll样受体(Toll-like receptors,TLRs)是一类最重要的模式识别受体,可识别内源性的损伤信号相关分子模式和微生物的病原学相关分子模式,是近年来发现的在免疫应答中起重要作用的细胞表面受体分子,其不仅激活先天性免疫应答,还引起细胞因子的释放,是连接天然免疫和获得性免疫的桥梁,对获得性免疫具有重要的影响[13,14]。此类受体的基因最早在果蝇体内发现,介导天然免疫,能感受入侵的病原体。TLR家族至少已包括13个成员,不同的TLR配体也有所不同。Medzhitov R等[15]首次在哺乳动物体内发现与果蝇中Toll受体结构同源的受体蛋白,后来命名为Toll样受体,主要表达于单核巨噬细胞膜上,树突状细胞、B细胞以及表皮细胞、脂肪细胞、成纤维细胞也有表达。其属于Ⅰ型跨膜蛋白(type Ⅰ transmembrance protein)受体,由胞外区、跨膜区、胞内区组成。胞外区为一段富含亮氨酸的重复序列(leucine-rich repeat,LRR),可与CD14 结合,参与各种病原体的识别;跨膜区是富含半胱氨酸的结构域;胞内区是一段约有200个氨基酸的序列,该序列与白细胞介素1(interleukin-1,IL-1)受体胞内区具有同源性,所以又称为Toll/IL-1 受体同源区(Toll/IL-1 receptor homologous region,TIR),是TLRs向下游进行信号传递的核心元件。当TLR4与相应配体结合后,信号转导到TIR区域,然后进一步激活核因子κB(nuclear factor-κB)、丝裂原蛋白激酶信号通路,诱导促炎细胞因子如肿瘤坏死因子和白介素等基因的表达[16]。
目前经TLR4介导的信号转导包括髓样分化因子88(myeloid differentiation factor 88,MyD88)依赖性和非依赖性途径。MyD88依赖通路通过MyD88和IL-1受体相关激酶(interleukin-1 receptor-assciated kinase,IRAK)相互作用,募集TNF-受体相关因子6(tumor necrosis fator receptor-assciated fator 6,TRAF6),激活的TRAF6导致IκB激酶复合物活化,最终导致NF-κB、P38、c-Jun氨基末端激酶(c-Jun N-terminal kinase,JNK)激活,进一步形成AP-1转录因子,从而诱导产生炎症因子。徐正婕等[17]通过建立大鼠NASH模型,观察肝脏内毒素受体表达的动态变化,结果发现实验8周时肝脏CD14和TLR4表达较正常组增高,12周进行性增高,并于16周达到高峰。Gao Y等[18]发现NASH患者体内TLR4、LBP、TNF-α的水平明显高于对照组,推测NASH的发病可能与TLR4及LPS的表达增高有关。Liu J等[19]对TLR4野生型和突变型小鼠给予高脂高糖饮食,发现分别在第4、8、16周出现野生型小鼠肝脏脂肪沉积、脂肪沉积合并炎症以及纤维化,TLR4、MyD88表达在第4周增加,第8周达到高峰,突变型小鼠肝脏脂质沉积则相对减轻,说明TLR4参与NAFLD的炎症和纤维化进程。刘涛等[20]采用高脂饲料喂养建立大鼠NAFLD模型,检测NAFLD组和正常组TLR4、MyD88肝组织mRNA和蛋白的表达,发现NAFLD组大鼠肝组织中TLR4、MyD88表达均增高,差异有统计学意义,研究认为TLR4信号在NAFLD大鼠模型中高表达,说明其参与NAFLD的炎症和纤维化进程。且Jagavelu K等[21]研究发现必须依赖MyD88的TLR4信号途径,内毒素才能促进炎症的产生。
本研究中,模型组的肝指数较正常组明显增加,并且模型组的NAS与正常组比较有显著差异,在分子水平模型组TLR4 mRNA表达水平明显高于正常组,同时作为TLR4下游蛋白的MyD88的mRNA结果也明显增高。综上所述,TRL4 介导的炎性反应可能是导致非酒精性脂肪性大鼠肝脏炎症发生发展的原因之一。
[参考文献]
[1] 胡卫,张苗旋,谭金. 非酒精性脂肪性肝病与代谢综合征相关性研究[J]. 中华流行病学杂志,2014,35(9):1062-1064.
[2] 邓银芝,周中银. 血清PTX3 脂肪因子在非酒精性脂肪肝病中临床意义[J]. 临床和实验医学杂志,2015,14(1):23-26.
[3] Piccinini AM,Midwood KS. DAMpening inflammation by madulating TLR signaling[J]. Mediators Inflamm,2010,2010:672395.
[4] Li YW,Zhang Y,Zhang L,et al. Protective effect of tea polyphenols on renal ischemiareperfusion injury via suppressing the activation of TLR4NF-κB p65 signal pathway[J].Gene,2014,542(1):46-51.
[5] 中華医学会肝病学会脂肪肝和酒精性肝病学组. 非酒精性脂肪性肝病诊疗指南(2010年修订版)[J]. 中华肝脏病杂志,2010,18(3):163-166.
[6] 马菁蔓,王景泉. 从虚论治非酒精性脂肪肝[J]. 长春中医药大学学报,2014,30(5):850-852.
[7] 黄甫竞坤,谢雯. 2012美国非酒精性脂肪性肝病诊断和管理指南[J/CD]. 中国肝脏病杂志(电子版),2014,6(4):91-98.
[8] Vuppalanchi R,Chalasani N. Nonalcoholic fatty liver disease and nonalcoholic steatohepatitis:Selected practical issues in their evaluation and management[J]. Hepatology,2009,49(1):306-317.
[9] Day CP. Pathogenesis of steatohepatitis[J]. Best Pract Res Clin Gastroenterol,2002,16(5):663-678.
[10] Hanley AJ,Williams K,Festa A,et al. Liver markers and development of the metabolic syndrome:The insulin resistance atherosclerosis study[J]. Diabetes,2005,54(11):3140-3147.
[11] 田培营,王炳芳. 非酒精性脂肪性肝病研究进展[J]. 同济大学学报(医学版),2006,27:46-48.
[12] 蒋艳明,施军平,等. 复方楂金颗粒剂对非酒精性脂肪性肝炎大鼠氧化应激的影响[J]. 中华中医药学刊,2012,30(9):2066-2068.
[13] 杨俊涛,张令强,贺福初. Toll样受体在肝脏疾病中的功能[J]. 中华肝脏病杂志,2006,14(8):632-634.
[14] Kawai T,Akira S. The role of pattern-recognition receptors in 1 innate immunity:Update on Toll-like receptors[J].Nat Immunol,2010,11(5):373-384.
[15] Medzhitov R,Preston-Hurlburt P. A human homologue of the Drosophila Toll protein signals activation of adaptive immunity[J]. Nature,1997,388(6640):394-397.
[16] ONeill LA. Signal transduction pathways activated by the IL-1 receptor/toll-like receptor superfamily[J]. Curr Top Microbiol Immunol,2002,270:47-61.
[17] 徐正婕,范建高,王兴鹏,等. 非酒精性脂肪性肝炎大鼠肝脏内毒素受体表达上调[J]. 中华肝脏病杂志,2006,14(1):49-52.
[18] Gao Y,Song LX,Jiang MN,et al. Effects of traditional Chinese medicine on endotoxin and its receptors in rats with non-alcoholic steatohepatitis[J]. Imflammation,2008, 31(2):121-132.
[19] Liu J,Zhuang ZJ,Bian DX,et al. Toll-like receptor-4 signalling in the progression of non-alcoholic fatty liver disease induced by high-fat and high-fructose diet in mice[J]. C|in Exp PharmacoI Physiol,2014,41(7):482-488.
[20] 劉涛,徐秋玲. 赵岩.ToII样受体4在高脂诱导非酒精性脂肪性肝病大鼠模型中的表达[J]. 临床肝胆病杂志,2015,31(12):2073-2076.
[21] Jagavelu K,Routray C,Shergill U,et al. Endothelial cell Toll-like receptor 4 regulates fibrosis-associated angiogenesis in the liver[J]. Hepatology,2010,52(2):590-601.
(收稿日期:2016-07-12)
