运动训练生物学基础模型的演变
——从超量恢复学说到运动适应理论
2017-02-13陈小平
陈小平

运动训练生物学基础模型的演变
——从超量恢复学说到运动适应理论
陈小平1,2
人体的运动能力为什么可以通过运动得到提高?这是一个人们长期努力要回答的问题,也是自运动科学化训练以来体育科学研究的重点和热点。“超量恢复”学说和“运动适应”理论是目前世界运动训练领域解释人体运动能力在训练中变化的经典。20世纪30年代,人们开始关注人体运动的能量代谢问题,60年代发现肌糖原储备在力竭性运动后出现超量恢复的现象。20世纪70年代中期,雅克夫列夫将肌糖原超量恢复扩展到整个竞技训练的范围,提出了运动训练的超量恢复学说。在20世纪50年代塞利提出的“应激-适应”理论影响下,“适应”从60年代开始逐渐成为运动训练的重要基础理论。之后,相继出现了“疲劳-适应模型”“机能储备模型”“改变-适应的时间动态模型”和“能力潜能元模型”等一系列试图揭示运动训练与运动能力增长之间关系、对运动能力变化进行预测和评估的学说和理论。但是,鉴于运动能力构成和发展的复杂性和当前人类对自身了解的局限性,迄今已有的各种模型还未能达到上述目标,还需要更加深入的研究。
超量恢复;适应;运动训练;演变Abstract:Why the body's sport ability can be improved through exercise? This is a question that people have been trying to answer for a long time and it is also the focus and hotspot of sports science research since sports scientific training."Supercompensation" and "adaptation" are two classic theory of the world in the field of sport training explaining human exercise ability changes in training.In 1930s,researchers began to focus on energy metabolism of human activity.In 1960s,the supercompensation’s phenomenon of muscle glycogen reserves was found in exhaustive exercise.In 1970s,Yakovlev extend muscle glycogen super-compensation to the area of the athletic training and super-compensation theory for sports training was proposed.Under the influence of the theory of “Stress Adaptation”by Selye in 1950s,“Adaptation”has become an important basic theory in sports training since the 1960s .After this,a series of doctrine and theory to attempt to reveal the relations between athletic ability growth and sports training and to predict and assess changes in exercise capacity,such as the " Fitness-Fatigue Model",the " Functional Reserves Model",the" Change-Adaptation Time Dynamic Model "and the “Performance Potential Metamodel” have appeared.However,given the complexity in composition and development of athletic ability and limitations in current human understanding of their own,so far a variety of models have not been able to achieve these objectives,and also require more in-depth to study.
运用“模型”揭示和解释运动能力在训练中的发展和变化是运动训练科学研究的一个重要领域,也是当前和未来发展的一个重要趋势。从“模型”的功能来看,它始终从两个方面影响着运动训练,首先是从源头探索运动训练对人体机能的影响,试图解释机能能力在运动训练刺激下产生变化的机制问题,被视为运动训练的生物学基础;其次是研究“负荷-疲劳-适应”之间的关系,试图对运动训练的过程和结果进行诊断、预测和控制,使运动训练成为一个可操作的科学化过程,也被看作是实现训练科学化的关键。
自20世纪60年代以来,运动训练领域相继出现了多种不同的解释运动能力在训练中变化的理论模型,其中最具影响的为超量恢复学说和适应理论。这些学说和理论不仅促进了竞技运动训练的发展,而且也引起各种学术观点的争论。本文拟对超量恢复和适应理论的起源、发展和演变进行梳理,对从20世纪20年代就已经提出来的肌糖原超量恢复现象,到之后出现的运动训练超量恢复学说,再到目前以“适应”理论为基础和背景的各种“模型”进行分析,以期推进和提升我国运动训练领域对该问题的认识。
1 超量恢复学说的起源与发展
1.1 肌糖原的超量恢复
人的运动能力为什么可以通过运动得到提高?人体的运动器官和系统为什么可以在运动训练中得到壮大?这样的问题是运动训练(包括竞技运动训练和大众健身训练)的基础和依据,是一个自运动训练进入科学化时代以来人们就开始关注的热点,也是迄今为止运动科学研究的一个重要领域。
早在19世纪末,一些生理学家就开始注意到运动对人体生理功能的影响。1890—1900年期间,德国科学家苏恩茨(N.Zuntz)就运用氧耗和二氧化碳代谢的方法测试人在安静和运动时的呼吸交换率(RER),他们的研究发现,人体运动的能量主要取决于膳食[59],不同的膳食,脂肪、蛋白质或碳水化合物导致肌肉工作时能量代谢的不同。在这一时期,法国科学家萨尔考维斯基(E.Salkowski)第一个发现了肌肉收缩过程中无机磷酸盐从有机化合物中的释放现象[59],为之后人体运动能量代谢的研究奠定了基础。20世纪初,德国人洛克斯(Roux)在《机体发展机制的报告和论文》[44]一书中,分析和总结了大量有关人体机能能力在运动过程中变化的研究成果,认为导致机体能力增加或降低的主要原因很可能与机体器官细胞的蛋白质质量的增长(肥大)或下降(萎缩)有关。
在同一时期,一些科学家开始从另一个角度对人体运动进行观察和研究。1919年,诺贝尔奖获得者、丹麦科学家科汝(J.Krogh)和林德哈德(J.Lindhard)发现,人的摄氧量在运动后5~10 min仍高于基础代谢水平的现象[31],表明肌肉收缩的停止并不意味着机体运动功能和能量代谢活动的终止,参与运动的器官和系统在运动停止后仍需要一定的时间使其机能恢复到安静状态。该研究不仅为之后希尔(A.V.Hill)等人发现“氧债”的形成与消除现象奠定了基础,更为重要的是拓宽了对运动训练的认识,将运动结束后的机体活动纳入到运动训练的范畴,开启了“运动后恢复”(Post-exercise recovery)这一新的研究领域。在此之后,人们开始关注运动对人体能量代谢的影响以及运动后人体能量代谢的变化等问题,这一研究领域的开启及其后续研究无疑为“超量恢复”学说的形成建立了研究基础。
进入20世纪20年代之后,世界上一批运动生理和生化学者发现,肌肉中的磷酸肌酸(phosphocreatine)随着肌肉的收缩出现下降,而在收缩后的恢复期又出现合成,这是最早对能量物质在运动中的消耗和再合成的研究。具有代表性的为德国的埃博登(G.Embden)和阿德勒(E.Adler)[18],美国的费斯克(C.H.Fiske)和苏巴罗夫(L.Subbarow)[19]、英国的P.艾格莱通(P.Eggleton)和G.P.艾格莱通(G.P.Eggleton)[17]等人的研究。他们的研究都证明,运动训练可以导致骨骼肌的糖原、肌氨酸和磷酸肌酸储备的增加,这是从能量代谢角度对“超量恢复”的早期发现。
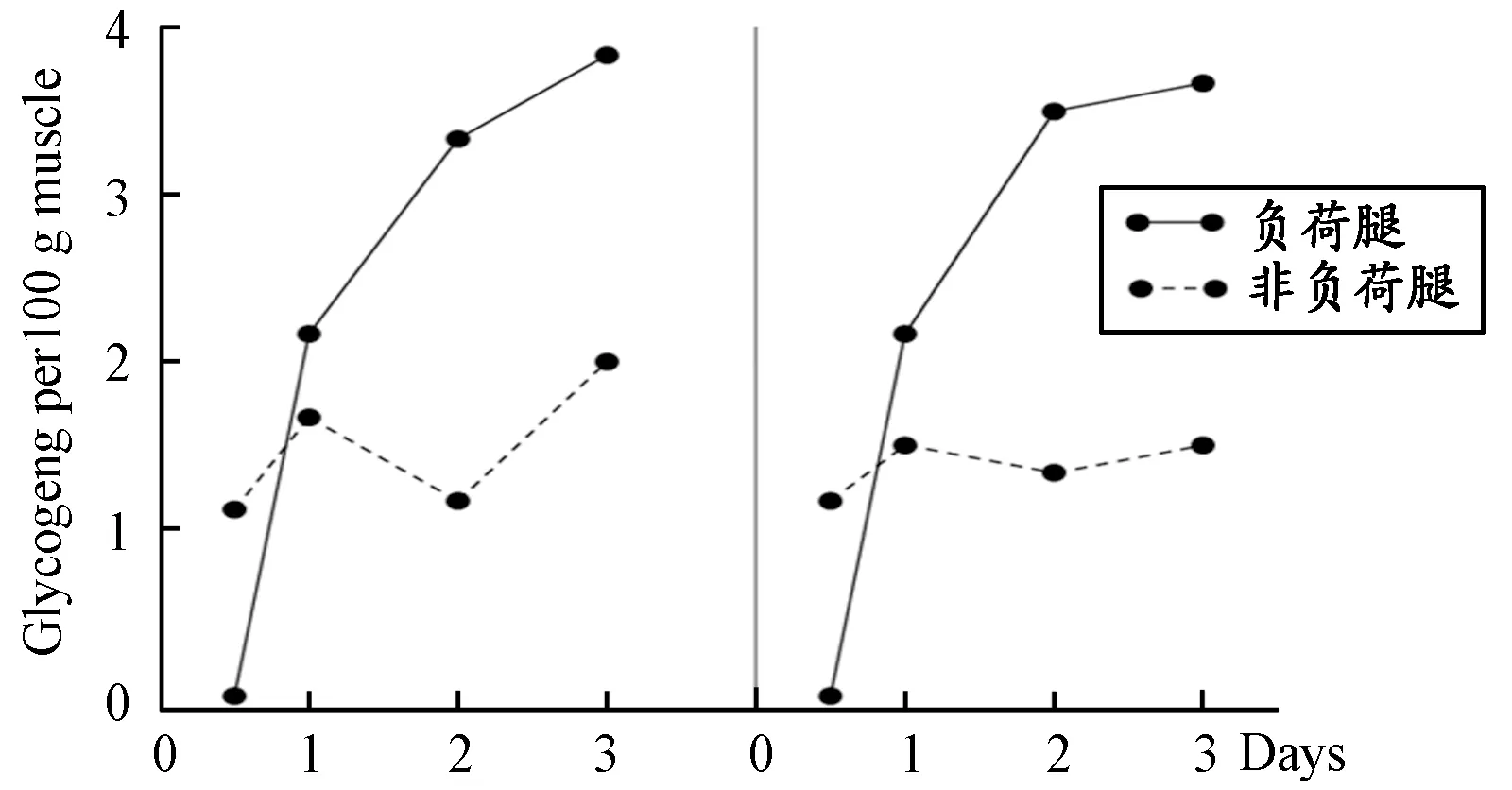
图 1 力竭负荷刺激后肌糖原的超量恢复Figure 1. Supercompensation of Muscle Glycogen after Exhaustive Load Stimulation (Bergstrom/Hultman,1966)
对人体运动时出现“超量恢复”现象的重要发现来自于瑞典的博格斯通(J.Bergström)和胡尔特曼(E.Hultman)等人的研究。需要强调的是,他们在研究中首次使用了人体肌肉生物活检针刺技术,该技术翻开了运动生理学和生物化学研究新的一页,对其后的研究产生了重大影响和积极推动作用。从营养的角度探索不同能量物质对肌肉力竭性持续收缩时间的影响,是他们最初的研究设想[7],其结果发现,人体力竭性运动的时间与参与运动肌肉的肌糖原储备量高度相关,以蛋白质和脂肪类为主的食物会降低肌糖原的储备并进而减少力竭性运动的时间,而补充碳水化合物为主的食物可以明显增加肌糖原恢复水平并延长运动时间。1966年,他们在世界权威刊物《自然》(Nature)上发表了题为“Muscleglycogensynthesisafterexercise:anenhancingfactorlocalizedtothemusclecellsinman”的论文[9],该研究后来成为“肌糖原超量恢复”(Glycogen supercompensation)和“碳水化合物负荷”(Carbohydrate loading)两个著名经典学说提出的重要基础。他们的研究发现,以1 200 kpm/min的负荷单腿骑功率车的极限运动后,运动腿的肌糖原含量迅速下降接近枯竭,但在补充高碳水化合物的条件下,训练停止后的第1天肌糖原开始快速恢复并达到未运动腿的水平,第2和第3天肌糖原储备继续增加并达到初始量2倍的水平(图1)。1967年,该项目的主要参与者胡尔特曼在其发表的综述文章中指出,“一系列的研究结果已经证明,参与工作肌肉中的肌糖原储备是限制肌肉长时间从事大负荷运动的主要因素”[21]。这一结论已经超出研究者原实验的“营养”设计初衷,从能量代谢的角度为日后肌糖原“超量恢复”的提出奠定了基础。
1.2 运动训练“超量恢复”学说的提出与核心
20世纪40~50年代,一些原苏联科学家开始对运动对肌肉的影响这一命题产生兴趣,最突出的是著名生理学家帕拉丁(A.V.Palladin)和他所率领的研究团队。他们从20世纪30年代就开始进行肌肉运动的能量代谢研究,相继发现运动训练可以提高磷酸原和肌糖原的水平和肌肉黄素的含量[39]。在他们的研究基础上,原苏联列宁格勒体育学院生物化学部主任,后被誉为苏联运动生物化学奠基人的雅克夫列夫(N.N.Jakowlev)为首的一些苏联学者,注意到肌糖原储备和营养补充对运动能力影响的问题,开始从运动生物化学的角度进行这方面的研究。1948年,与雅克夫列夫在同一研究所工作的雅姆珀里斯卡娅(L.I.Yampolyskaya)博士,运用电刺激的方法对冬蛙运动后肌糖原储备的变化进行了测试。她的实验结果表明,冬蛙肌肉在104次/min 收缩频率下肌糖原出现持续的下降,在刺激停止后的休息期肌糖原出现明显和稳定的超量恢复现象,肌肉运动后肌糖原的超量恢复水平及其稳定性程度取决于肌肉收缩的强度和运动的持续时间[57]。在该研究中,作者根据人体负荷后肌糖原储备出现的“下降、恢复和超量恢复”的特性,提出运用“超量恢复”解释运动训练对人体机能能力影响作用的设想,即将运动训练对人体机能能力产生影响作用的机制归结为“超量恢复”。从时间上看,雅姆珀里斯卡娅在其研究论文中明确提出了“超量恢复”的概念,并对运动后肌糖原超量恢复的水平及其稳定程度做了初步的定性分析,尽管在她的文章中也提到包括雅克夫列夫在内的其他的一些原苏联学者的类似研究成果,但就目前所能看到的原苏联研究文献中,该研究是最早提出肌糖原“超量恢复”名词的文章。在该研究成果发表之后,雅克夫列夫也相继通过实验得到了运动后肌糖原储备出现先下降,后恢复到初始水平,再后出现显著性提高的类似结果,进一步证实了的确存在肌糖原超量恢复这一事实[25]。
雅克夫列夫和他的团队不仅一直致力于运动训练的生化基础研究,而且,其研究密切结合训练实践,对训练负荷、运动疲劳和超量恢复等问题非常关注,并成为他们长期的研究重点。1972年,他首次将“超量恢复”作为人体运动能力提高的生理机制,认为对运动员施加训练负荷的主要目的就是打破机体已经形成的“内环境平衡”(Homoeostasis),运动能力的提高就是基于“超量恢复”机理的在更高的层次上建立新的“平衡”[26]。1975年,他又撰文对训练过程中机体内环境的破坏与重建机理进行了深入分析,指出训练负荷量和强度的增加是造成机体(特别是肌肉系统)出现“超量恢复”的主要途径[27]。1977年,雅克夫列夫在专著《运动生物化学》中将“超量恢复” 作为解释运动训练效果的生物学机制[28],将运动负荷刺激下肌糖原的“下降-恢复-超量恢复”过程扩展到解释整个人体机能能力的增长。
此后,“超量恢复”迅速成为运动训练生物学“基础”的一个重要组成部分[3-5],并被延伸作为“超量负荷”(overload)训练原则的依据[11,54](图2)。“超量恢复”学说认为,机体在负荷的刺激下,其能量储备、物质代谢以及神经调节系统的机能水平出现下降(疲劳),在负荷后这些机能能力不仅可以恢复到负荷前的初始水平,而且能够在短期内超过初始水平,达到“超量恢复”的效果。如果在“超量恢复”阶段适时给予新的负荷刺激,“负荷-疲劳-恢复-超量恢复”的过程则可以不断地在更高的水平层次上周而复始的进行,由此使机体的能力得到不断的持续提高[45]。
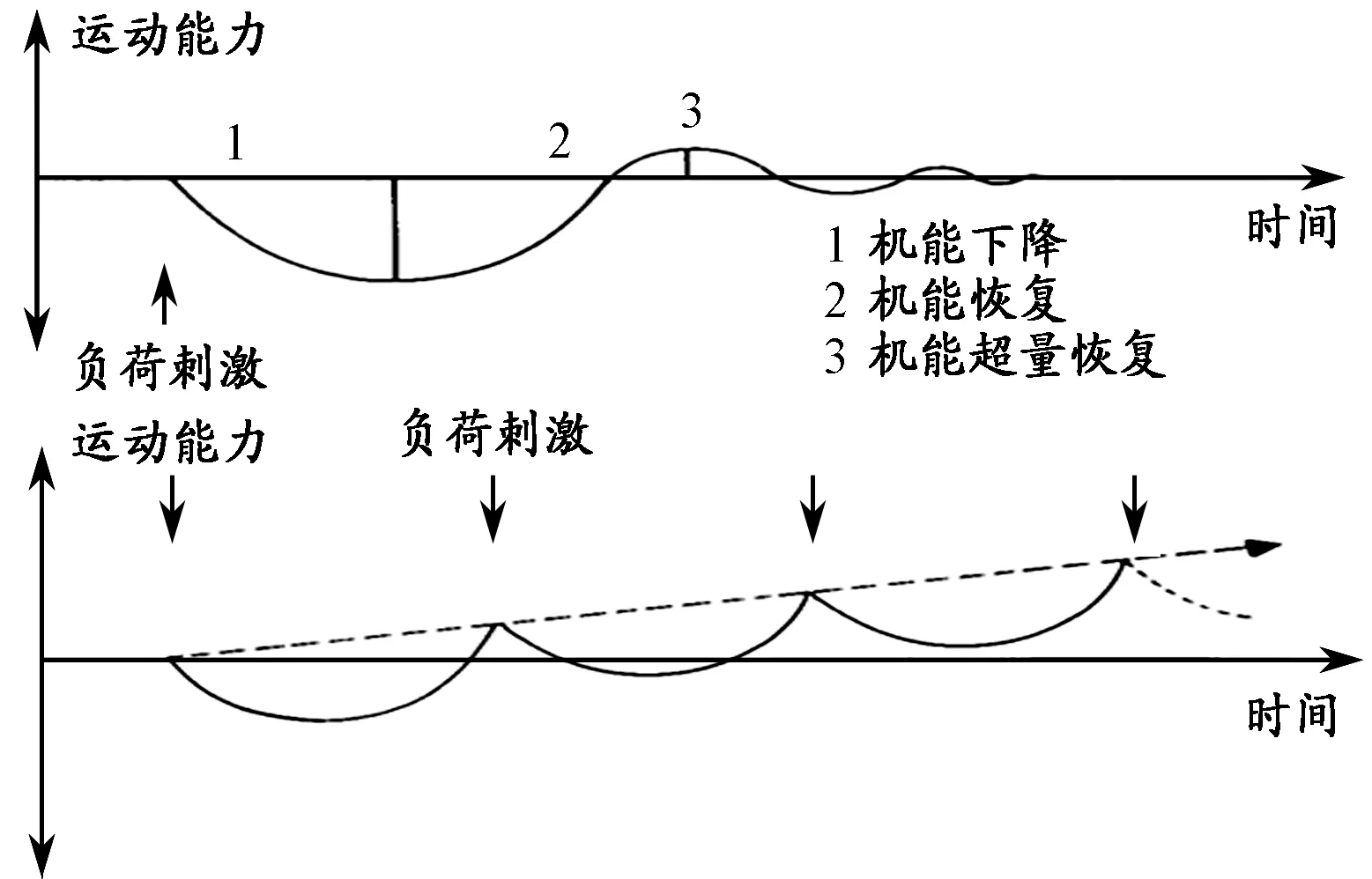
图 2 超量恢复示意图Figure 2. Schematic Diagram of Supercompensation (Weineck,1994)
1.3 “超量恢复”学说的发展和意义
总体上,“超量恢复”学说的形成可以分为两个阶段,肌糖原超量恢复阶段和运动训练超量恢复阶段。从20世纪20年代到70年代初,超量恢复的概念主要还是被限定在基于肌糖原、磷酸原、线粒体以及一些相关酶类运动后出现“下降-恢复-超量恢复”的一种能量代谢现象。在此期间,包括原苏联在内的学者们基本都在进行着以负荷、疲劳、恢复和能量代谢为关键词的生理生化研究。自20世纪70年代初,原苏联的雅克夫列夫将肌糖原超量恢复的概念扩展为运动训练的超量恢复之后,该名词及其理论就不再被限定在生理和生化领域,而被广泛运用到运动训练理论和实践当中,成为解释运动训练效果变化以及训练负荷与能力增长之间关系的基础依据。
在能量代谢语境下的“超量恢复”至今仍然是运动生物学领域的研究重点和热点。在瑞典博格斯通和胡尔特曼等人对肌糖原超量恢复的经典研究之后,人们开始将研究的视角转向运动后的能量消耗和补充问题,并逐渐将其发展成一个相对独立的研究领域。研究人员对运动后的能量恢复进行了更为细致的划分,将从运动后即刻能量消耗的最低点到恢复到运动开始时的水平称为“能量储备的重新注入”(Repletion of energy stores),将超出能量储备原有水平的部分称为“超量恢复”(Supercompensation)。其后的一些研究基本围绕着这2个阶段进行,重点集中在不同能量物质运动后的补充性恢复和超量恢复的时间、幅度以及影响因素等问题。
在运动后能量物质的恢复(重新注入)方面,胡尔特曼等人[21]对肌肉的高能磷酸化合物(ATP)、磷酸肌酸(CP)和糖原等能量物质的恢复速度进行了研究,其结果表明,ATP-CP(磷酸原系统)的恢复明显快于糖原,在运动终止后60 s ATP-CP就可以恢复到运动开始水平的70 %左右,ATP的完全恢复大约需要2 min,而CP的完全恢复则至少需要3 min以上的时间。特尔君格(R.L.Terjung)等人[50]对不同类型肌纤维力竭训练后糖原的恢复速度进行了研究,肌糖原运动后合成率最快的发生在氧化快肌(FOG),慢肌(SO)的合成速度比氧化快肌慢2倍左右,糖原合成速度最慢的是无氧快肌(FG)。他们的研究结果还发现,在力竭性运动后肌糖原的恢复在时间上并不是线性的,30~60 min区间的糖原合成率最高,即运动后第1个30 min和60 min后的合成速度均低于第2个30 min。另外,还有研究发现,运动后糖原的恢复分为快速和慢速2个阶段,糖原的快速恢复期一般都发生在运动结束后的4~6 h,在快速合成阶段糖原的恢复速度主要取决于运动中糖原的消耗量,而在慢速阶段则取决于胰岛素的分泌水平[20,40]。
在运动后能量物质的超量恢复方面,以雅克夫列夫为代表的原苏联学者在20世纪60~70年代进行了大量研究,其研究发现,运动后肌糖原和磷酸原出现超量恢复的时间不同,肌糖原的超量恢复大约发生在运动结束后的1~24 h,而磷酸原的超量恢复则早于肌糖原[15]。他们的研究还发现,糖原和磷酸原超量恢复发生的时间和程度与训练强度密切相关,在短距离高强度游泳后,糖原和磷酸原均出现快速合成,而在长距离低强度游泳后,两种能量物质的超量恢复速度和幅度均出现下降[6]。
雅克夫列夫和他的研究团队在20世纪70年代中期将肌糖原超量恢复拓展为对整个运动训练进行解释的运动能力超量恢复学说之后,仍然继续从运动生物学的角度为该学说的建立寻找更多的依据。1978年,他们在动物实验中发现,骨骼肌线粒体蛋白在运动后即刻出现下降,而在1 h恢复后实验组的线粒体蛋白明显高于对照组,从而认为肌肉线粒体也具有运动后的超量恢复现象[26]。在此之后的研究中,他们又发现磷酸元和肌糖原的超量恢复在慢肌的发生早于快肌的现象[30]。
无论是早期的磷酸原和肌糖原的超量恢复,还是之后由雅克夫列夫提出的作为解释人体运动能力提高机制的超量恢复,都对竞技运动训练产生了重要和深远的影响。在理论上,超量恢复学说突出了运动训练的能量代谢基础,认为能量代谢水平及其在训练中的变化既是运动训练的目标也是运动训练的依据,使运动训练过程的控制和效果的评价成为可能。同时,进一步强调了运动训练对身体能力的“延时”作用,引发并促进了运动后能量代谢的研究,使之形成了一个相对独立的研究领域,且至今仍是运动生物科学研究的重点和热点,其相关研究结果不断成为支撑和指导运动训练的重要基础和依据。在实践上,人们通过“超量恢复”进一步认识到训练负荷和机体恢复是2个对运动训练效果具有同样重要作用的因素,它们之间存在某种的关系,这种关系在很大程度上决定了运动训练的质量和效率,在高度重视训练负荷投入的同时,加强了对训练后各器官和系统恢复的关注。
1.4 超量恢复学说的不同观点和争论
自从将“超量恢复”作为解释整个运动训练的机制以来,该理论一方面在运动训练领域得到迅速传播和认可,成为运动训练的重要基础;另一方面,也受到运动科学界,尤其是运动生物学领域学者的广泛质疑。
在20世纪60年代初,原苏联运动训练学者马特维也夫(Matwejew,L.P.)就将超量恢复作为运动训练的重要基础之一,认为人体竞技能力的提高是一个“刺激-疲劳-恢复-超量恢复”的过程[32]。德国运动训练学者温奈克(Weineck J.)在其专著《最佳训练》(OptimalesTraining)中将“超量恢复”作为负荷与恢复最佳关系原则的生物学基础,提出了能力下降、能力恢复和能力超量恢复的3个阶段,同时也指出,如果训练不当也会导致运动能力下降的风险[54]。加拿大运动训练学者邦帕(Bompa T.O.)和哈弗(Haff G.G.)在其对世界运动训练极具影响的《周期——运动训练理论与方法》一书中,将超量恢复和适应理论相结合,提出运动训练将对运动员造成3种不同的结果,正常训练、过度训练和超量恢复。他们认为,“超量恢复是运动和能量再生之间的一种关系,它能够在比赛前使运动员产生超量的机体适应,即生理的代谢和神经系统的唤醒”[11]。从这些有关论述来看,“超量恢复”理论的提出从能量代谢的角度为运动员机能能力的长期发展奠定了基础,促使教练员在制定训练计划时高度关注负荷和恢复的关系,将不同能量物质的储备、消耗和恢复作为影响运动能力发展的重要因素。
对雅克夫列夫“超量恢复”理论的质疑主要来自运动生理、生化领域。至今,欧美出版的运动生理学、运动医学和运动生物化学著作中基本都不出现“超量恢复”理论的内容和表述[23,33,55],说明这些具有指导运动训练功能的书籍并没有将“超量恢复”作为运动训练的生理基础。蒂-玛瑞斯(de Marées)在其专著《运动生理学》中甚至明确指出:“所谓的‘超量恢复’至今仍然在许多方面缺乏科学的证据。”[16]霍尔曼(Hollmann)和海廷格尔(Hettinger)认为,将肌糖原等能量物质在运动训练极限负荷条件下出现的短暂性超量恢复现象扩展到对整个运动训练机制的程度,是该理论存在的主要问题,不能将肌糖原这一单一指标的超量恢复现象延伸扩展到解释整个机体对运动训练的适应[30]。齐默尔曼(Zimmermann)也强调,运动训练的适应是一个长期的、带有强烈个体特征的过程,而不像超量恢复模型描述的那样简单[1]。
2 适应理论的起源与发展
2.1 适应理论的提出及其在运动训练领域的应用
20世纪30年代,先后在加拿大麦基尔大学和蒙特利尔大学执教,被誉为适应理论之父的塞利(H.Selye)从病理学的角度提出了生命的“应激”(Stress)和“适应”(adaptation)问题[46]。他认为,“应激”和“适应”是各种生物体的基本特征,任何一种作用于人体器官的侵害性刺激都是由专门病源性和非专门病源性2个部分组成,机体对非专门病源刺激的反应被称作“一般适应综合症”(general adaptation syndrome,GAS)。同时,“适应”还可以被分为不同的形式,如生理、心理、社会等不同的适应[47]。1956年,塞利出版了著名的《生命的应激》(TheStressofLife)一书[48],标志着适应理论的问世。塞利将人体的“一般适应综合症”或“应激反应”视为一个过程,并且将其分为报警(Alarm)、抵抗(Resistance)和疲惫(Exhaustion)3个阶段。报警阶段是机体对刺激的一种生理唤醒和动员期,又分为冲击阶段(Shock)和抵抗冲击阶段(counter shock);抵抗阶段是机体对刺激的防御和忍耐期;疲惫阶段是机体对刺激抵抗力的丧失期。1976年,塞利又提出“局部适应”的问题,一些器官或功能可以通过蛋白质结构的改变而发生局部性的适应[49]。这一点对运动训练尤其重要,通过专门有针对性的训练可以优先发展某一器官或系统的机能。
从运动训练的角度来看,塞利的适应理论为运动训练构建了一个宏观的生物学基础,即人体对外在环境的自然适应过程。在该语境下,运动训练以其各种不同的方式形成了多种不同的“应激源”(Stressors),训练负荷就是外界对机体的一种刺激,而“适应”则是机体对训练负荷的“代偿性”变化,这种变化具有大小、快慢和优劣的特性,它的走向在很大程度上决定了运动能力的发展水平。在微观上,适应理论的核心观点“一般适应综合症”及其所包含的3个发展阶段,极其形象地描述了人体与刺激的动态变换关系以及在该关系下运动能力的各种变化。当运动员受到训练负荷的刺激时,机体首先进入 “报警”反应,参与运动的各器官和系统的工作强度显著提高,如神经-肌肉、心血管和能量代谢等系统的工作能力明显增强。随着负荷刺激的持续或增强,机体进入第2个应对级别,即抵抗阶段,在此阶段机体各器官与系统不断与外来的负荷刺激进行对抗,刺激与抵抗形成胶着态势。运动能力的增长主要发生在这个阶段,机体在与训练刺激对抗的过程中逐渐提升自身的能力,在功能和结构上逐渐产生适应性改变。然而,此时尽管生物体一直在抵抗外来刺激的压力,但其自身也变得较为“脆弱”,也更易于受到其他外来刺激的冲击,如运动员在持续疲劳状态下更易于受到疾病和损伤等其他因素的干扰和侵害。在训练负荷的持续压力下,连续不断的疲劳积累导致机体进入疲惫阶段,在该阶段虽然机体已经对训练负荷产生了一定程度的适应,但在训练负荷仍然保持高位水平甚至持续增加的情况下,其抵抗力就无法继续与之抗衡,一个额外的高负荷或长时间高负荷的维持都会对机体造成破坏性的打击,具体表现为竞技能力的下降甚至过度训练的发生。
2.2 运动适应理论的发展和意义
塞利的应激-适应理论一经问世就受到竞技训练领域的关注。1960年,美国《径赛技术》(TrackTechnique)杂志主编威尔特(F.Wilt)就认识到塞利的研究成果对运动训练的重要指导意义,发表了题为“应激与训练”(StressandTraining)的文章[56]。在文章中,他认为塞利提出的人体具有一个有限度的适应能力的观点,为人们探索和解释运动能力在训练中变化的机制提供了一个崭新的视角。随后,澳大利亚游泳国家队教练,悉尼大学讲师卡尔利勒(F.Carlile)也发表了“运动员与应激适应”(TheAthleteandAdaptationtoStress)的文章[13],认为运动员的训练是一个机体对训练负荷的适应过程,适宜的训练负荷可以提高运动员的运动能力,而不足或过大的训练负荷都会降低运动员的运动能力。同时,他还从适应理论的角度指出了当时运动训练中存在的大量不科学的训练方法和内容,提出应该从生物学、教育学和哲学等基础学科的角度和层面加强竞技训练的研究。
早期接受适应理论并将其思想运用到运动训练实践的典型代表人物是美国著名游泳教练员康希尔曼(James Counsilman)博士。20世纪50年代,在美国印第安纳大学担任游泳教练的康希尔曼,在塞利适应理论刚一问世就将这一思想迅速引入他的训练,以该理论作为指导思想建立了被称为“印第安纳系统”(The Indiana System)的游泳训练体系。1968年,在他撰写的被誉为当代游泳训练“圣经”的《游泳的科学》(TheScienceofSwimming)一书中[14],就将最大适应应激、循序渐进和动机作为制定训练计划的三大原则,并将运动员训练后的机体最大适应进一步分为不适应、疲劳、适应和超适应4个等级。他以1周的训练为例(图3),描述了运动员的训练负荷和机能表现可能出现3种不同的结果:如果运动员的训练负荷从周一到周五始终保持在“中等”水平的话,他的机体能力则处于“疲劳区域”,经过周六和周日的恢复其能力可以恢复到“适应区域”(A曲线);如果运动员的训练负荷较高,机体疲劳程度处于疲劳区域的下限(但没有达到不适应区域),经过周末的恢复机能能力则可能出现“超适应”的水平(B曲线);如果运动员的训练负荷过大,机能能力下降到不适应区域,尽管他也经过了周末的休息,但他的机能能力水平仍然无法达到“适应区域”或“超适应区域”(C曲线)。显然,康希尔曼的训练引入了塞利适应理论的核心思想,基于适应理论的“预警、抵抗和衰弱”三段论将运动员的训练结果分为提高(超适应)、保持(适应)和下降(不适应)3种不同的结果,而这些不同的训练结果取决于训练负荷的适时和适量的投入。

图 3 应激、疲劳、适应和超适应Figure 3. Schematic Diagram of Stress,Fatigue,Adaptation and Super Adaptation (Counsilman, 1968)
将塞利的适应理论系统运用于运动训练领域的学者是爱沙尼亚塔尔图大学的维禄(A.Viru)教授。20世纪80年代初,维禄开始运用适应理论指导运动训练[52],他把一般的非专门性反应,即应激反应,看作是人体专门适应形成的必备条件,一般应激水平的发展可以有力地支持特定系统适应能力的改善。他认为,训练负荷是“应激源”,它必须是一个足够大和有效的刺激,才会对机体产生作用。机体能力在负荷刺激的作用下出现障碍(疲劳),又随着应激水平的提高形成新的平衡状态(能力提高)。机体对刺激的适应是一个在中枢神经支配和调控下的过程,中枢神经协调并校正不同器官和系统在负荷下的协作机能,使机体出现一种“有效的训练结果”。外界的刺激如果达不到或超出了神经中枢协调和控制的范围,机体则可能对该刺激不做出反应或出现负面反应[1]。
2008年,维禄撰写了《竞技训练的适应》(AdaptationinSportsTraining)一书[53],从应激-适应的角度全面和深入地诠释了运动能力在训练过程中的增长、保持和下降,是迄今将运动训练放在机体适应背景下进行研究和讨论的重要著作。该书对与运动密切关联的神经-肌肉系统、内分泌系统和能量代谢系统在训练刺激作用下的变化进行了研究,不仅从传统的运动生理或运动生化层面对这些变化进行了阐述,而且,从运动适应的角度对这些变化的原因进行了分析。将“运动后恢复期”(Post-exercise Recovery Period)作为一个章节进行专门论述,是该书的一个主要特点。维禄认为,肌肉收缩的停止并非意味着机体运动的结束,运动后的恢复同样应纳入运动训练的范畴,应该关注各种重要能量物质运动后的恢复和超量恢复。在宏观上,他将运动后机体的恢复分为2个阶段和5个层面(图4),在运动后几分钟到几小时的快速恢复阶段,机体一些主要功能指标,如心率、血压、乳酸等,首先快速恢复到训练前的安静水平,之后机体内环境平衡也恢复到运动前的状态,一些重要的在运动中大量消耗的能量物质储备,如ATP-CP和糖原等,也在该阶段被重新补充甚至出现超量恢复。在被称为延迟恢复(Delayed recovery)的阶段,运动能力和机能将得到恢复和重建。关于延期恢复问题,维禄在书中强调,“关于身体机能和工作能力的延期恢复,一种错误的理解是该阶段开始于第1阶段的结束,然而该阶段实际上应开始于运动的结束”[53]。也就是说,对于一些能量物质或机体能力来说,它们的恢复也同样开始于运动结束的即刻,但它们的恢复和重建比那些在短时间内能够快速恢复的功能和能量物质需要更长的时间,因此而贯穿快速和延迟恢复2个阶段。

图 4 运动后机体恢复的阶段划分Figure 4. Stage of Recovery after Exercise (Viru,2008)
2.3 基于运动适应理论的若干“模型”
随着适应理论在运动训练领域应用的发展,出现了若干基于该理论的运动机能模型,引起学术界的关注和研究。
加拿大的班尼斯特(E.W.Banister)和他的同事们从20世纪70年代开始关注运动训练中刺激、疲劳和适应三者之间关系的问题,他们以大学生游泳运动员全年训练的数据为素材,建立了游泳运动员的疲劳-能力数学模型[7,51]。1982年,班尼斯特正式提出了具有一般训练意义的“疲劳-适应模型”(Fitness-Fatigue Model)[8],与雅克夫列夫提出的“超量恢复”这一运用疲劳单一因素解释运动训练效果的模型比较,该模型也被称为运用疲劳和适应2个指标描述机能能力发展的双因素模型(图5)。班尼斯特认为,人体在负荷的刺激下会同时出现机体疲劳和适应2种反应,这2种相互对抗(antagonistic)的反应会根据负荷的变化而变化,且二者之间存在某种关系。在高负荷训练的情况下,疲劳的上升快于适应,其曲线高于适应曲线,但随着负荷的下降疲劳曲线的下降速度加快并逐渐与适应曲线出现交汇,其交汇点被认为是运动员出现最佳竞技状态的临界点,重大比赛应该出现在该临界点之后,即最佳竞状态区域出现。疲劳-适应双因素模型不仅推导出竞技能力长期发展的基本规律,而且给出了在一定负荷刺激下疲劳与适应的关系,人们可以通过实验找到二者之间的关系,使运动训练负荷过程的定量诊断和控制成为可能。

图 5 疲劳-适应模型Figure 5. Fitness-Fatigue Model (Banister, 1982)
班尼斯特的疲劳-适应双因素模型对赛前减量训练具有重要指导意义,在某种意义上开启了赛前训练量化研究的先河。在其问世之后,人们运用该模型的思想或方法对多个运动项目进行了赛前训练的研究[36],同时也在其基础上对模型本身进行了研究,提出了诸如“非线性模型”(nonlinear model)[12]等改进型赛前训练模型。
尽管班尼斯特的双因素模型已经比超量恢复模型有了很大的进步,人们可以借助该模型寻找负荷(疲劳)与适应(能力)之间的量化对应关系,预见未来竞技能力(状态)的发展水平,但是,该模型仍然存在其局限性,人们无法通过该模型对所有运动项目进行预测和评估,而只能对那些力竭性体能项目具有一定的指导意义。同时,该模型也不能区别不同的训练刺激对机体造成的不同类型的疲劳,如神经疲劳、肌肉疲劳和心血管疲劳等,而只能从整体上构建刺激与疲劳和适应之间的关系,影响到模型的准确性。
20世纪80年代后期,人们开始意识到,人体运动能力的增长、保持和消退与人体运动生理学,尤其是能量代谢系统紧密相关,不能仅从投入(负荷刺激)和产出(能力改变)2个端口进行研究,而必须深入到训练过程中探求其各个器官、系统的变化。在这方面,较为典型的研究成果是德国的马德尔(Mader)的“机能储备模型”(Functional Reserves Model)[34]和诺依曼(Neumann)的“转变-适应模型”(Change-Adaptation Time Dynamic Model)[37]。
20世纪80年代末,德国运动生理学者马德尔教授从细胞代谢的层面研究了人体机能能力在负荷刺激下变化的问题。他认为,适应的个体因素藏匿在细胞遗传的机制之后,细胞蛋白质分解与合成之间的关系是人体运动机能能力变化的主要原因,蛋白质自身所具有的衰弱、凋亡和再生时间以及在运动刺激条件下的分解与合成在很大程度上决定了人体运动能力的增长、保持或下降。因此,他提出,人体除了具有一个正常状态下的细胞蛋白质标准代谢率之外,还存在一个在负荷作用下的“机能受限性”蛋白质代谢率的额外增长,如果该增长发生在那些对运动能力具有重要影响甚至决定性作用的蛋白质(线粒体等),则必然带来机体运动能力的提高。当然,如果外来刺激(负荷)过小或者超过了机体的承受能力,都会影响到蛋白质的分解和合成率,具体表现为运动能力的停滞或下降。
在上述认识基础上,马德尔提出了“机能储备模型”(图6)。“最大机能容量”“适应储备”“现实机能容量”“机能储备”和“现实生理负荷”是机能储备模型的5个关键元素。“最大机能容量”是人体已经具有的各种能力和一个尚未开发的“适应储备”的总合,既反映了运动员当前已经具备的总体最高运动能力,也显示出运动员的最大潜在能力储备。“适应储备”是一个受遗传限制并可以枯竭的能力区域,它的空间越小表明运动员能力的可塑性越小,其下限代表着运动员目前所具备的运动水平,其上限是最大机能能力的顶端,也是运动能力的极限。“现时机能容量”是运动员各种子能力目前所具有的可供开发的潜能,它由“现时生理负荷”和“机能储备”两部分构成,前者是机体各种不同能力已具备的各自的现时状态,也是至今为止机体对训练的适应程度,后者是这些子能力可以支配但还未达到的机能储备。上述三者之间的关系形象地揭示了训练负荷的运用规律,在运动训练中,如果运动员的“现时生理负荷量”越逼近“现时机能容量”,也就是说,“机能储备”的余留量越少,说明训练负荷量是最佳的。如果训练负荷低于或超过了供支配的“现时机能”储备,则表明训练负荷的安排不正确。这两种情况均会影响训练的效果,过低的负荷不会引起机体的反应,机体也不会出现相应的适应,而过高的负荷同样不会给机体带来良性的适应,甚至造成过度训练和损伤。同时,“现时机能容量”也是人体不同子能力的代名词,各个“现时机能容量”之间以及其生理负荷与机能储备的大小也存在区别和差异,它们既是有限的,又在一定范围内是动态变化的。不同“现时机能容量”的总和构成了人体的“最大机能容量”,其中,不同的机能储备又构成了总体的适应储备,对不同机能储备的挖掘,即生理负荷的不断增长,也是对整个机体“适应储备”上限的逼近[2]。
诺依曼在1991年提出了人体的“转变-适应”模型[38]。该模型从时间的角度对机体细胞和组织在运动训练负荷作用下的适应性变化进行了解释,提出了机体各器官系统受到足够大的外来刺激时会产生“转变”,在反复施加负荷的条件下则出现“适应”,具体表现为机能能力的提高。同时,该理论还根据各器官系统的适应形成在时间上的不同,将适应的过程划分为4个阶段并为各个阶段的实现给予了明确的时间期限(图7)。

图 7 人体的“转变-适应”模型Figure 7. Change-Adaptation Time Dynamic Model (Neumann, 1991)
由图7可见,人体对运动训练产生适应性改变首先发生在神经-肌肉系统的层面,在7~10天的训练中,肌肉细胞中的底物和神经对肌肉的支配程序出现相应的变化。在之后的10~20天训练中,机体的能量储备出现显著性增加,肌肉蛋白质的结构也同时出现变化。随后,运动适应进入第3个阶段,具体表现为能量物质的动员和转换速度加快,肌肉的募集和收缩速度也得到相应的提高。当训练进行到6周左右的时候,机体适应进入到第4个阶段,这也是运动适应过程的最后阶段,参与运动的各个不同器官和系统之间的协调得到明显的改善,不仅运动时的动力出现明显增强,而且其经济性也得到显著优化。
该理论描述了在负荷情况下机体的平衡状态被破坏而产生“转变”,在连续适宜负荷的作用下又形成新的平衡,即一个从量变(积累作用)到质变的“适应”过程。我们注意到,诺依曼的理论中没有运用传统的“超量恢复”来解释机体器官系统对负荷的反应,而称其为“转变”。依据这一观点,机体在外来负荷的作用下其平衡状态被破坏,具体的反应就是疲劳,当负荷条件不存在时,机体会得到恢复,如果反复对机体施加负荷刺激,则机体会由于负荷的累积效果对该刺激产生适应,具体表现为运动器官系统在形态机能方面的改变。这种适应是以刺激的累积效应为基础,首先实现机体器官形态的“代偿”性改变,然后产生功能上的适应。例如,力量训练使神经对肌肉的支配作用得到改善和骨骼肌面积得到增加,从而增大了力量。
20世纪90年代之后,世界计算机技术的快速发展开启并推动了人体运动适应模型的智能化研究。2000年,德国美因茨大学计算机系的珀尔(J.Perl)和科隆体育大学运动与训练研究所的梅斯特(Mester J.)提出了“竞技潜能元模型”(Performance Potential Metamodel)[35,41],简称为PerPot的模型仍然秉承了“应激-适应”理论的核心思想,运用应力(Strain)输入、应答(Response)输出以及二者对竞技能力(Performance)的影响三者之间形成的若干交互作用,以各种“元”,即不同的系统和功能的形式模拟竞技能力的适应过程(图8)[42]。该模型认为,运动能力的改变是机体抵抗(antagonistic)外来刺激的结果,该结果具有滞后(delay)效应的特点,不仅表现为正面的能力提高,而且也会出现负面的能力下降[43]。
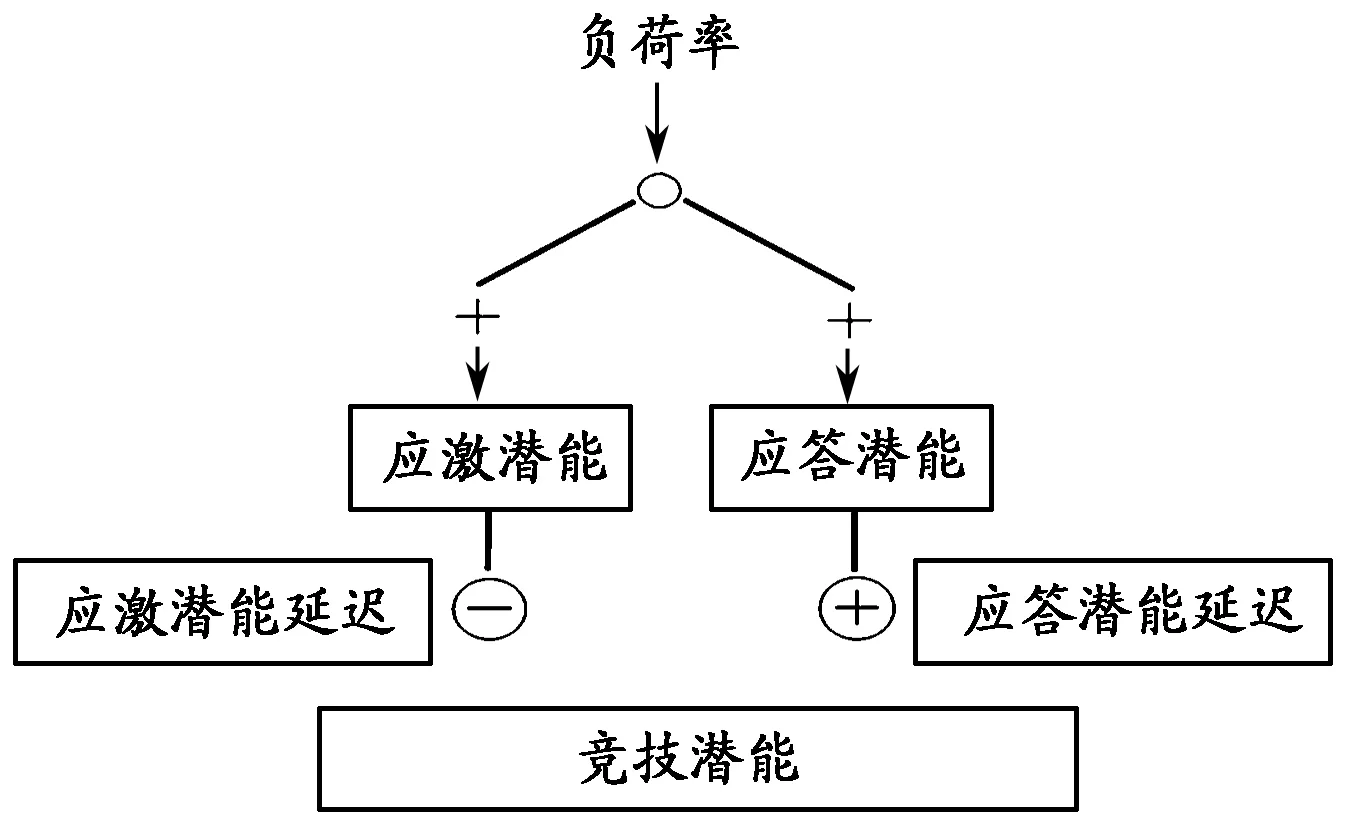
图 8 竞技潜能元(PerPot)模型Figure 8. Performance Potential Metamodel (Perl,2000)
与之前的竞技能力模型比较,PerPot模型的突出特点在于构建了一个基于计算机和相关数学建模技术的模拟平台。在该平台上,人们可以通过一些已经被研究证明的刺激(训练)与应答(适应)之间的关系,如跑步强度与心率之间的关系、血红蛋白增长与耐力训练时间之间的关系等,模拟或预测竞技能力的发展趋势。
3 运动机能模型发展展望
3.1 从历史发展的视角分析和评价运动机能模型
纵观运动训练机能模型的发展,可以将其分为3个阶段。
20世纪20年代,人们开始发现磷酸肌酸和肌糖原等能量物质存在运动后的超量恢复现象,标志性的成果为瑞典学者博格斯通和胡尔特曼等人1966年的研究,从方法和路径上为此后的研究奠定了基础,这一时期是运动机能模型研究的初始阶段。
1977年,原苏联著名学者雅克夫列夫在其专著《运动生物化学》中将肌糖原超量恢复现象扩展为运动训练的超量恢复,提出了“超量恢复”学说。雅克夫列夫“超量恢复”学说的问世标志着运动训练机能模型的研究进入到一个新的阶段,从运动训练的层面对大量能量代谢的基础研究从哲学的高度进行归纳和总结,在宏观上建立了机体在接受负荷刺激和作出应答过程中的机体疲劳-恢复-超量恢复3者之间的关系,构建了首个以解释运动训练效应为目标的理论模型。该模型首次超越了传统的运动生理、生化的生物学领域,没有将重点集中在磷酸肌酸、肌糖原等具体能量物质在运动训练前后产生的复杂变化方面,而是将运动训练中的能量代谢过程简化并提炼为下降、恢复和超量恢复3个阶段,高度概括了在训练刺激下机体机能的变化规律。该模型的优点主要表现在两个方面:首先,是将机体在训练条件的诸多复杂变化进行了高度简化和凝结,人们可以将其作为制定训练负荷和选择训练方法的依据,也可以将各种训练后机体的变化归因于“超量恢复”,便于教练员和运动员对训练负荷及其效果的理解;其次,是该模型具有运动生物学能量代谢基础的支撑,大量的实验研究已经证明肌糖原等能量物质超量恢复现象的存在,与之前的运动训练比较,超量恢复模型的应用无疑在一定程度上提高了运动训练的科学化水平。
第3个阶段实际上开始于20世纪50年代中期塞利的适应理论问世之后,美国的威尔特和康希尔曼分别在理论上和实践上率先将适应理论引入竞技运动训练,特别是康希尔曼在游泳训练上的卓越成功验证了适应理论对竞技运动训练具有重要指导作用。1980年,爱沙尼亚学者维禄开始深入推动适应理论在运动训练领域的发展,运用适应理论的思想解释机体在训练负荷下的变化,从此,“适应”逐渐取代“超量恢复”成为竞技运动训练的生物学基础。在20世纪80年代之后,运动训练机能模型的研究进入一个新的活跃期,相继出现了班尼斯特的“疲劳-适应双因素”模型、马德尔的“机能储备模型”、诺依曼的“转变-适应”模型和珀尔的“竞技潜能元”模型。这些模型尽管在影响力上难以超越超量恢复和适应理论,但它们融合了当代先进的科技思想和技术,在运用层面上更加贴近运动训练实践,表现出较好的实操性。
长期以来,尽管一些学者不断对雅克夫列夫提出的“超量恢复”模型提出质疑,认为该模型存在诸多问题。但是,我们应该注意到,在当时的体育科研水平下,该学说无论在生物学基础上,还是在运动训练实践上,都不失为是一个“最贴近”训练实际的模型。作为一名杰出的运动生化学者,雅克夫列夫和他的研究团队一直致力于从能量代谢层面探索人体机能能力在训练中变化的机制,而并非仅从哲学和方法论的角度解释这一问题。
塞利的适应理论在运动训练领域一直扮演着一种指导思想的角色,在某种程度上可以说是一种从宏观层面指导运动训练的思维方式或方法论。在超量恢复之后出现的相当部分模型是在适应理论思想的基础上或在其影响下构建和形成的,这些模型基本都以适应理论的“报警-抵抗-疲惫”机体对外来刺激反应的“三段论”为基础,强调人体机能能力的提高源于机体对外来刺激的适应,承认人体都具有个体的机能能力极限,重视不同大小和形式的刺激将引起机体不同的适应。
与“超量恢复”相比,“适应”理论在一些重要的条件上增加了“限制”,如提出了人体运动极限的存在,在一些具体的解释上更加符合运动的实际,如强调了不同运动员个体和不同机能能力对负荷的不同“适应”,突出了个体差异和区别对待。在对运动训练机制解释的客观性方面,“适应”理论显然具有较多的优势。但必须注意到,目前在“超量恢复”和“适应”理论问题上的争论并没有结束,尽管人们对“超量恢复”学说提出了质疑,但是依据“应激”理论将运动训练的生理、生化基础归因于机体的“适应”同样缺乏大量具体实验数据的支持,人体运动能力提高的机制在许多方面仍然是“黑箱”,以目前的科学理论和技术还无法做到精确的量化解释。因此,我们应该站在历史发展的立场审视和评价“超量恢复”和“适应”这2个对运动训练最具影响的理论,继续加强这方面的深入研究。
3.2 运动机能模型研究的发展
从当代科学技术发展的总趋势来看,人体运动机能模型研究的未来发展主要集中在以下3个方面。
首先,以超量恢复和适应理论为主干的运动机能模型的多年发展,仍然基本被局限在运动科学的范围内,即使是20世纪90年代基于一定计算机技术的“竞技潜能元”模型,与当前科技的发展水平仍然存在很大的差距。近年来,多学科和高科技在运动训练领域的运用已经明显改变了传统的训练,将互联网、大数据、人工智能、虚拟仿真等高科技的理念与技术运用于模型的开发和构建,将成为模型研究的一个未来发展趋势。
其次,进一步加强有关基础研究是运动机能模型研究未来发展的另一个主要趋势。人体机能模型涉及到遗传和后天训练两方面因素,涉及到训练负荷和机体对负荷刺激的各种生理和心理反应以及二者之间的关系,其中既有一般性规律也有个体化特点。因此,以模型为研究目标的若干基础研究,如基因、能量代谢和营养补充等问题,应成为模型研究的重要领域,这些领域的研究成果将成为未来模型构建的基础和依据。
第三,理论与实践的紧密结合是模型研究的一个重要发展方向。截至目前的各种运动机能模型在很大程度上大多停留在理论的层面,而并不能直接应用于对运动训练的指导,即其理论意义高于实践作用。究其原因,一是运动训练本身的复杂性和个体性,二是模型功能的客观性和可重复性,都还不能满足训练实践的需求。因此,在未来模型的研究中,应该将实际运用作为研究的主要目标,将训练负荷、机体疲劳和机体适应三者的关系作为研究的重点,运用模型指导运动员竞技能力的长期发展和竞技状态的短期调控。
[1]陈小平.运动训练的基石——“超量恢复”学说受到质疑[J].首都体育学院学报,2004,4(16):3-7.
[2]陈小平.当代运动训练热点问题研究:理论与实践亟待解决的问题[M].北京:北京体育大学出版社,2005:4-5.
[3]冯炜权.体育运动的生物学知识[M].北京:人民体育出版社,1980:74-79.
[4]过家兴,田麦久,等.运动训练学[M].北京:人民体育出版社,1990:80-85.
[5]田麦久,等.运动训练学[M].北京:人民体育出版社,2000:133.
[6]BATUNER L S.Restitution of energy potential of muscles after exercises close to maximal[J].Sechenov Physiol.J.USSR,1977,63:406.
[7]BANISTER E W,CALVERT T W,SAVAGE M V,etal.A systems model of training for athletic performance[J].Aust J.Sports Med,1975,7(5):61
[8]BANISTER E W.Modeling elite athletic performance[M]//MACDOUGALL J D,WENGER H W,GREEN H J.Physiological Testing of Elite Athletes.Human Kinetics,1982:403-425.
[9]BERGSTRöM J,HULTMAN E.Muscle glycogen synthesis after exercise:an enhancing factor localized to the muscle cells in man[J].Nature,1966,16(210):309-310.
[10]BERGSTRöM J,HERMANSEN L,HULTMAN E,etal.Diet,muscle glycogen and physical performance[J].Acta Physiol Scand,1967,71:140-150.
[11]BOMPA T O,HAFF G G.周期——运动训练理论与方法[M].李少丹,李艳翎,译.北京:北京体育大学出版社,2011:11-15.
[12]BUSSO T.Variable dose-response relationship between exercise training and performance[J].Med Sci Sports Exe,2003,35:1188-1195.
[13]CARLILE F.The Athlete and Adaptation to Stress[M].Track Technique,September,1961:56-158.
[14]COUNSILMAN E J.The Science of Swimming[M].Prentice-Hall,Inc.Englewood Cliffs,New Jersey,1968.
[15]CHAGOVETS N R.Biochemical changes in muscles in restitution period after physical work[J].Ukr Biochim Zh(Kiev),1957,29:450.
[16]DE MARÉES H.Sportphysiologie[M].Sport und Buch Strau,1996:168.
[17]EGGLETON P,EGGLETON G P.The physiological significance of “phosphagen” [J].J.Physiol,1927,63:155-161.
[18]EMBDEN G E.Adler Uber die physiologische Bedeutung des Wechsels des Permeabilitatszustandes von Musklfasergrenzschichten[J].Z.physiol.Chem.,1922,118:1-11.
[19]FISKE C H,SUBBAROW Y.Phosphorus compounds of muscle and liver[J].Science,1929,70:381-382,
[20]HERMANSEN L,VAAGE O.Lactate disappearance and glycogen synthesis in human muscle after maximal exercise[J].Am J Physiol,1977,233:E422.
[21]HULTMAN E.Physiological role of muscle glycogen in man,with special reference to exercise[J].Circ Res,1967,20-21:199-1114.
[22]HULTMAN E,BERGSTROM J,MCLENNAN-ANDERSON N.Breakdown and resynthesis of phosphocreatine and adenosingtriphosphate in connection with muscular work in man[J].Scand J Clin Lab Invest,1967,19:56.
[23]HOLLMANN W,STRUDERH K.Sportmedizin(5.Auflage)[M].Schattauer,2009.
[24]HOLLMANN W,HETTINGER T.Sportmedizin,Arbeits- und Trainingsgrundlagen[M].Stuttgart-New York,1980.
[25]JAKOVLEV N N.Biochemical foundations of the training of muscles[J].Usp Sovrem Biol(Moscow),1949,27:257.
[26]JAKOVLEV N N.Die bedeutung der homöostasestöruang für die effektivität des trainingsprozesse[J].Medizin Und Sport,12(1972),367.
[27]JAKOVLEV N N.Biochemistry of sport in the soviet union:Beginning,development,and present status[J].Med Sci Sports,1975,4(7):237-247.
[28]JAKOVLEV N N.Sportbiochemie[M].Barth,Leipzig,1977.
[29]JAKOVLEV N N,KRASNOVA A F,LENKOVA R L,etal.Effect of the adaptation to intensive muscle activity on the functional state of skeletal muscle mitochondria[J].Cytologia(Moscow),1978,14:197.
[30]JAKOVLEV N N.The expenditure and resynthesis of energy sources in muscles of various functional profile in regard to the character of load[J].Sechenov Physiol J USSR,1979,65:1796.
[31]KROGH J,LINDHARD J.The changes in respiration at the transition from work to rest[J].J Physiol,1919,53:431.
[32]MATWEJEW L P,KOLOKOLOWA W M.Allgemeine Grundlagen der Körpererziehung[M].Berlin(DDR),1962:20.
[33]MACLAREN D,MORTON J.Biochemistry for Sport and Exercise Metabolism[M].Wiley-Blackwell,2012.
[34]MADER A.Aktive belastungsadaptation und regulation der proteinsynthese auf zellulaerer ebene[J].Deut Z Sportmed,1990,41:40-58.
[35]MESTER J,PERL J.Grenzen der anpassungs- und leistung-sfähigkeit aus systemischer sicht-zeitreihenanalyse und ein informatisches metamodell zur untersuchung physiologischer adaptationsprozesse[J].Leistungssport,2000,30(1):43-51.
[36]MUJIKA I.The influence of training characteristics and tapering on the adaptation in highly trained individuals:A review[J].Int J Sports Med,1998,19:439-446.
[37]NEUMANN G,SCHUELE K P.Sportmedizinische Funktionsdiagnostik[M].Leipzig Berlin Heidelberg:Barth Verlagsgesellschaft,1994:13-17.
[38]NEUMANN G,BERBALK A.Umstellung und Anpassung des Organismus-grundlegende Voraussetzung der sportlichen Leistungsfaehigkeit[M]//BEMETT P,JESCHKE D.Sport und Medizin Pro und Contra.W.Zuckschwerdt Verl.,Munchen,1991:415-419.
[39]PALLADIN A V.The biochemistry of muscle training[J].Science,1945,102:576-578.
[40]PIEHLl K.Time course of refilling of glycogen-induced glycogen depletion[J].Acta Physiol.Scand,1974,90:297.[41]PERL J.Antagonistic adaptation systems:an example of how to improve understanding and simulating complex system behavior by use of meta-models and on-line simulation[C]//16th IMACS Congress.New Brunswick,NJ:Rutgers University,2000.[42]PERL J.Per Pot:A metamodel for simulation of load performance interaction[J].Eur J Sport Sci,2001,1(2):1-13.
[43]PERL J.Perpot- a meta-model and software tool for analysis and optimisation of load-performance-interaction[J].Int J Perform Anal Sport,2004,4(2):61-73.
[44]ROUX W.Vorträge und Aufsätze über Entwicklungsmechanik der Organismen[M].Leipzig,1905.
[45]RÖTHIG P U A.Sportwissenschaftliches Lexikon[M].Verlag Hofmann Schorndorf,1992:490-491.
[46] SELYE H.A syndrome produced by diverse nocuous agents[J].Nature,1936,138(2):230-231.
[47]SELYE H.Einführung in die Lehre vom Adaptationssyndrom[M].G.Thieme,Stuttgart,1953.
[48]SELYE H.The Stress of Life[M].New York:McGraw-Hill,1956.
[49]SELYE H.Stress in Health and Disease[M].Butterworth,Boston,1976.
[50]TERJUNG R L,BALDWIN K M,WINDER W W,etal.Glycogen repletion in different types of muscle and in liver after exhausting exercise[J].Am J Physiol,1974,226:1387.
[51]CALVERT T W,BANISTER E W,SAVAGEN M V,etal.A Systems Model of the Effects of Training on Physical Performance[J].IEEE Trarsactions Systems Man Cybemetics,1976,SMC-6(2):94-102.
[52]VIRU A.Das Problem der Vergröβerung der plastischen reserven des organismus im trainingsproze?[J].Leistungssport ,1980,10:280-282.
[53]VIRU A.Adaptation in SportsTraining[M].Informa Healthcare,New York,2008.
[54]WEINECK J.Optimales Training[M].Spitta Verlag GmbH,2000:32-35.
[55]WILMORE J H,COSTILL D L,KENNEY W L.运动生理学[M].王瑞元,汪军,译.北京:北京体育大学出版社,2011.
[56]WILT F.Stress and Training[J].Track Technique,1960,(1):6.
[57]YAMPOLYSKAYA L I.Muscle glycogen supercompensation in restitution period after exercises of various character and duration[J].Byull.Eksp.Biol.Med.(Moscow),1948,11:358.
[58]ZIMMERMANN E.Trainingsprinzipien[M].Sportwissenscha-ftliches Lexikon,Schorndorf,1983:423-424.
[59]ZUNTZ V.N.UMSATZ der NAHRSTOFFE.XI.Betrachtungen uber die beziehungen zwischen Nahrstoffen und Leistungen des Korpers[M]//Handbuch der Biochemie des Menschen und der Tiere,edited by K.Oppenheimer.Jena,FRG:Fischer,1911:826-855.
Evolution of the Biological Basis Model in Sports Training—From Supercompensation to Adaptation Theory
CHEN Xiao-ping1,2
supercompensation;adaptation;sportstraining;evolution
1000-677X(2017)01-0003-11
10.16469/j.css.201701001
体育科学
2016-07-26;
2016-12-30
国家社会科学基金资助项目(13BTY049)。
陈小平 ,男,教授,博士,博士生研究生导师,主要研究方向为运动训练,Tel:(010)87182557,E-mail:chenxiaoping@ciss.cn。
1.宁波大学 体育学院,浙江 宁波 315211;2.国家体育总局体育科学研究所,北京 100061 1.Ningbo University,Ningbo 315211,China;2.China Institute of Sport Science,Beijing 100061,China.
G808.1
A
