大白菜玻利玛细胞质雄性不育系92—09A的形态特征及相关基因的克隆与表达模式分析
2017-01-21王红霞李洪雷郝建梅丁谦张一卉王
王红霞+李洪雷+郝建梅+丁谦+张一卉+王凤德+邓永林+王均邦+何启伟+李景娟+高建伟

摘 要:玻利玛细胞质雄性不育(Pol CMS)是目前很有利用价值的胞质雄性不育源。在前期的研究中,我们利用柯桂兰提供的大白菜Pol CMS不育源为母本,以大白菜骨干亲本92-09为轮回亲本,培育出大白菜Pol CMS不育系92-09A。本研究对大白菜Pol CMS不育系92-09A及其保持系92-09、恢复系早28的形态特征进行观察,并对相应的不育基因Braraorf224及其恢复基因BraraPPR进行了克隆及生物信息学分析。结果表明,大白菜Pol CMS不育系92-09A的花瓣、萼片和雌雄蕊都比保持系92-09小,不能形成有活力的成熟花粉粒;其保持系和杂种F1(不育系92-09A×恢复系早28)均可以产生有活力的花粉粒。大白菜Pol CMS不育基因Braraorf224及恢复基因BraraPPR与甘蓝型油菜的Pol CMS不育基因及恢复基因同源性很高。qRT-PCR结果显示,BraraPPR在恢复系早28和杂种F1中的表达量均明显高于雄性不育系92-09A。本研究可为进一步揭示大白菜Pol CMS雄性不育机理奠定基础。
关键词:大白菜;Pol CMS;不育基因;恢复基因
中图分类号:S634.1+Q78文献标识号:A文章编号:1001-4942(2016)12-0006-10
Abstract Polima cytoplasmic male sterile (Pol CMS) is a very important CMS source in plant breeding. In the previous study, we used Pol CMS line KG-Pol of Chinese cabbage provided by Professor Ke Guilan as female parent and core line 92-09 as recurrent parent to breed Pol CMS line 92-09A of Chinese cabbage. Here, the morphological characteristics of Chinese cabbage Pol CMS line 92-09A, its parental maintainer line 92-09 and restorer line Zao 28 were investigated, and the Pol CMS gene Braraorf224 and its restorer gene BraraPPR were cloned and analyzed. The results showed that the petals, sepals, stamens and pistils of Chinese cabbage Pol CMS line 92-09A were all smaller than those of its maintainer line 92-09, and it could not form mature pollen with viability. While the maintainer line 92-09 and the F1 hybrid of Pol CMS line 92-09A×restorer line Zao 28 both had pollen grains with viability. Bioinformatics analysis indicated that the nucleic acids and amino acids of Chinese cabbage Pol CMS gene Braraorf224 and restorer gene BraraPPR were very close to those of Brassica napus.The results of qRT-PCR showed that the expression level of BraraPPR in restorer line Zao 28 and F1 hybrid were both higher than that in Pol CMS line 92-09A. The research would be helpful to understand the molecular mechanism of Pol CMS of Chinese cabbage in the future.
Keywords Chinese cabbage (Brassica rapa L. ssp. pekinensis); Polima cytoplasmic male sterile (Pol CMS); Male-sterile gene; Restorer gene
大白菜起源于中国,是我国的特产蔬菜。山东是大白菜的重要起源地之一,有丰富的种质资源,纯系间杂交往往表现出极强的杂种优势。大白菜杂种优势利用是靠一代杂种实现的。利用雄性不育系生产一代杂种是最理想的制种技术之一[1]。
大白菜雄性不育主要包括核基因雄性不育和核质互作雄性不育。由于使用核不育系配制的商品种子实际上是三交种,所以核不育系在生产中应用的难度较大。目前报道的大白菜核质互作雄性不育源主要来自于甘蓝型油菜玻利玛(Polima)细胞质雄性不育(Pol CMS)和萝卜Ogura细胞质雄性不育(Ogura CMS)材料。甘蓝型油菜Polima雄性不育源的发现大大促进了大白菜优势育种的发展[1,2]。
1972年,傅廷栋等[3]发现油菜Pol CMS雄性不育株,随后利用Pol CMS实现了三系配套,培育出优良油菜品种华杂2号、湘杂8号及华杂3号。此后,Pol CMS被传至欧美等国,培育出多个油菜杂交种。1985-1994 年, 中、加、澳、印等国共注册(审定)22 个油菜杂交种, 17个注明不育系来源中, 13个利用玻利玛雄性不育材料。Pol CMS成为目前很有利用价值的胞质雄性不育源[4]。Polima不育性的利用受到了其它植物育种学家的重视。任成伟等(1992)[5]将油菜Pol CMS 不育材料与小白菜进行种间杂交和回交, 获得小白菜不育系;向长萍等(2000)[6]用红菜薹大股子作转育亲本, 转育Polima雄性不育系, 经4 年6 代选择, 育成原始不育系。在大白菜育种方面,柯桂兰等(1992)[7]选育出大白菜Pol CMS不育系,其它大白菜育种工作者也相继选育出多个Pol CMS不育亲本材料[8]。
1994年,我们利用柯桂兰提供的Pol CMS雄性不育源,选育出大白菜Pol CMS不育系92-09A。本研究拟通过大白菜Pol CMS不育系92-09A的选育、形态特征观察,并对其相应的不育基因、恢复基因进行克隆及生物信息学、表达模式分析,为进一步揭示大白菜Pol CMS雄性不育机理奠定基础。
1 材料与方法
1.1 材料选育与培养
1.1.1 大白菜Pol CMS雄性不育系92-09A的选育 以柯桂兰馈赠的雄性不育KG-Pol植株为母本,以大白菜骨干亲本纯系92-09为轮回亲本,连续回交6代,选育获得大白菜Pol CMS雄性不育纯系92-09A。92-09即为92-09A的保持系。不育系92-09A与早28产生的一代杂交种(“鲁白13号”,又称“西白1号”)花粉可育,早28即为92-09A的恢复系。
1.1.2 材料培养 取92-09A、保持系、恢复系及92-09A×早28 F1种子,萌发后4℃低温春化4周,于3月15日前后移栽至山东省农业科学院蔬菜花卉研究所实验田内,保证其正常开花,于盛花期分别取花和花蕾进行形态学观察、花粉活力鉴定及RNA提取。
1.2 花器官形态观察及花粉活力鉴定
采用Nikon Eclipse TE2000-S显微镜对雄性不育系92-09A、保持系92-09及92-09A×早28 F1的花器官形态进行观察,采用亚历山大染液对花粉活力进行鉴定,方法参考Alexander(1969)[9]。
1.3 基因克隆、生物信息学及表达模式分析
1.3.1 RNA提取和反转录 取当天和第二天花瓣展开的Pol CMS不育系92-09A、恢复系早28及92-09A×早28 F1的花和花蕾分别提取总RNA。采用TRNzol-A+试剂盒(天根)进行RNA提取,方法参考说明书。反转录按照TaKaRa PrimeScript RT reagent Kit with gDNA Eraser (Perfect Real Time)产品说明书操作。
1.3.2 大白菜Pol CMS雄性不育基因Braraorf224的克隆 以大白菜Pol CMS不育系92-09A总RNA的反转录产物为模板,用引物(正向引物:5′-ATGCCTCAACTGGATAAATTCACTT-3′;反向引物5′-TCATCGAAATAGATCGAGGATTT-3′)对Pol CMS雄性不育基因Braraorf224进行全序列PCR扩增。PCR反应体系(20 μL)为:2×高保真PCR Mix 10 μL,上游引物(10 mmol/L)0.5 μL,下游引物(10 mmol/L)0.5 μL,模板cDNA 1 μL,ddH2O 8 μL。PCR反应程序为:94℃ 3 min;94℃ 30 s,58℃ 30 s,72℃ 1 min,36个循环;72℃ 10 min。
1.3.3 大白菜Pol CMS 雄性不育系恢复基因BraraPPR的克隆及表达模式分析 以恢复系早28总RNA的反转录产物为模板,用引物(正向引物:5′- ACACGAAATTATCACCAA-3′;反向引物5′- CTGAAACTTTAAGACGGA -3′)对Pol CMS 雄性不育系的恢复基因BraraPPR进行全序列PCR扩增。PCR反应体系(20 μL)为:2×高保真PCR Mix(天根) 10 μL,上游引物(10 mmol/L)0.5 μL,下游引物(10 mmol/L)0.5 μL,模板cDNA 1 μL,ddH2O 8 μL。PCR反应程序为:94℃ 3 min; 94℃ 30 s,58℃ 30 s,72℃ 3 min,36个循环;72℃ 10 min。
Real-time PCR使用TaKaRa公司的SYBR green PCR master mix,仪器为IQ5 Real-Time PCR system (BIO-RAD, Hercules,CA,USA),八联管为AXYGEN公司生产,内参基因为Actin2。扩增Actin2的正向引物为5′-AGGAAGGATCTGTACGGTAAC-3′,反向引物为5′-TTCTGTGAACGATTCCTGGAC-3′。扩增BraraPPR的正向引物为5′-CCCCACAGAGGTTTAGTCCC-3′,反向引物为5′-TCACCAACCCTTCCTACCTTACA-3′。荧光定量PCR反应采用两步法进行: 95℃ 2 min; 95℃ 15 s,60℃ 30 s, 45 个循环,并在反应结束后制作熔解曲线,检测是否有引物二聚体生成。
1.3.4 生物信息学分析 利用克隆得到的基因序列在GenBank中进行BLAST检索,查找Pol CMS雄性不育基因Braraorf224及恢复基因BraraPPR在其它物种中的同源序列。利用DNAMAN 6.0软件对来自不同物种的Pol CMS雄性不育基因及恢复基因的氨基酸序列进行比对并构建系统进化树。
2 结果与分析
2.1 大白菜Pol CMS 雄性不育系92-09A的选育及其形态特征
2.1.1 大白菜Pol CMS雄性不育系92-09A及保持系 自1972年被发现以来,Pol CMS雄性不育源已被广泛应用于甘蓝型油菜三系配套生产制种工作中[2]。1992年,柯桂兰等将甘蓝型油菜Pol CMS雄性不育源应用于大白菜育种中,育成了以Pol CMS为不育源的大白菜核质互作雄性不育系[7]。1994年,我们以柯桂兰馈赠的大白菜Pol CMS雄性不育KG-Pol植株为母本,以大白菜骨干亲本纯系92-09为轮回亲本进行连续回交,历经6年,选育获得了大白菜Pol CMS雄性不育纯系92-09A。大白菜骨干亲本92-09即为Pol CMS雄性不育系92-09A的保持系。保持系92-09是以亚蔬中心的夏阳为基础材料经多代自交选育的稳定大白菜自交不亲和系。
利用亲本92-09与另一个骨干亲本早28配置组合,获得了大白菜杂交新品种“鲁白13号”(又称“西白1号”)。由于亲本92-09是Pol CMS雄性不育系92-09A的保持系,所以我们利用不育系92-09A×早28大面积生产“鲁白13号”一代杂种。一代杂交种(92-09A×早28)长出的植株,花粉可育,因此大白菜亲本早28即为Pol CMS雄性不育系92-09A的恢复系。
大白菜Pol CMS雄性不育系92-09A、保持系92-09外叶深绿、无毛,叶柄绿色,球叶叠抱,叶球绿色,早熟,耐高温高湿。“鲁白13号”是山东省审定的第一个夏白菜品种,该品种外叶绿色、无毛,叶柄浅绿,球叶扣抱,叶球浅绿色,耐高温高湿,山东地区6月起垄播种,45~50天成熟,品质优,口感好,成熟后及时收获[10]。
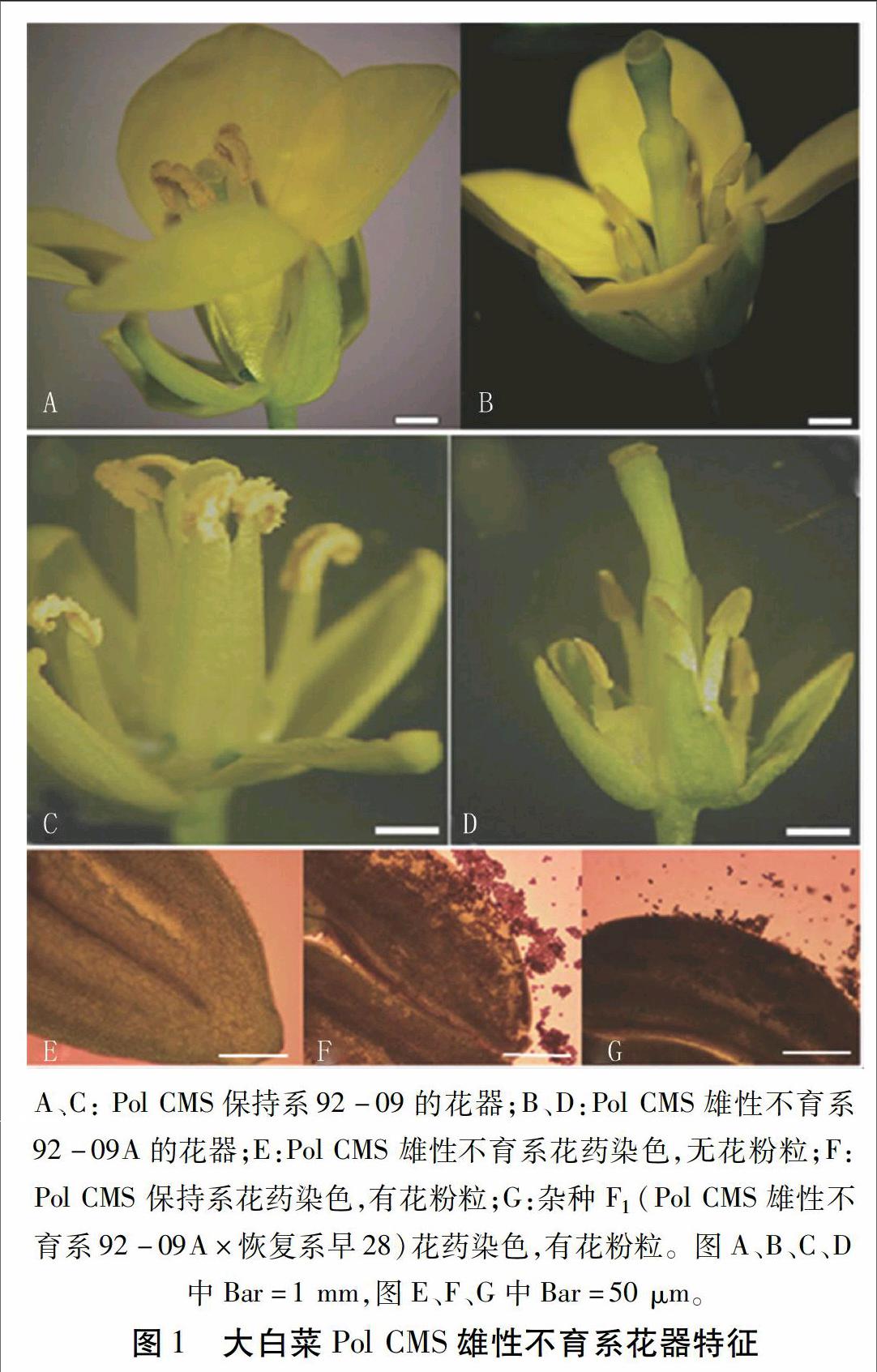
2.1.2 大白菜Pol CMS雄性不育系的花器结构特点 图1是大白菜Pol CMS雄性不育系92-09A、保持系92-09及杂种F1(92-09A×早28)花器结构、花粉粒形态的部分图片。大白菜Pol CMS雄性不育系92-09A的花蕾较保持系92-09小, 但外观看比较饱满,无干扁的症状。遇低温及生长弱等不利环境条件时,会出现败蕾现象,但是到后期这一现象通过规范管理可有效地消除。花器大小方面, 不育系92-09A植株的花瓣、萼片、雌雄蕊都比保持系92-09小(图1A~D)。且保持系92-09的花药饱满,花粉充足,具有明显的散粉现象(图1A、C);不育系92-09A雄蕊短小瘦弱,花丝短缩,紧箍在雌蕊基部,花药锥形,内无花粉(图1B、D)。保持系92-09、杂种F1(92-09A×早28)可以产生圆形饱满的花粉粒(图1F、G);而不育系92-09A不能形成圆形饱满有活力的成熟花粉粒(图1E)。不育系92-09A除花药不育、花器略微瘦小外,其它器官如子房、蜜腺等功能正常,这为利用胞质雄性不育系从事大规模制种提供了可能。
2.2 大白菜Pol CMS雄性不育基因Braraorf224的克隆及序列分析
2.2.1 大白菜Pol CMS雄性不育基因Braraorf224的克隆 以大白菜Pol CMS不育系92-09A花、花蕾总RNA反转录产物为模板,对大白菜Pol CMS雄性不育基因Braraorf224进行PCR扩增,获得669 bp的条带。测序后通过序列比对,初步认定该基因是甘蓝型油菜Pol CMS雄性不育基因Branorf224 (Accession No. EU254235)的同源基因,命名为Braraorf224。Braraorf224基因编码222个氨基酸,编码区内不含有内含子(图2)。
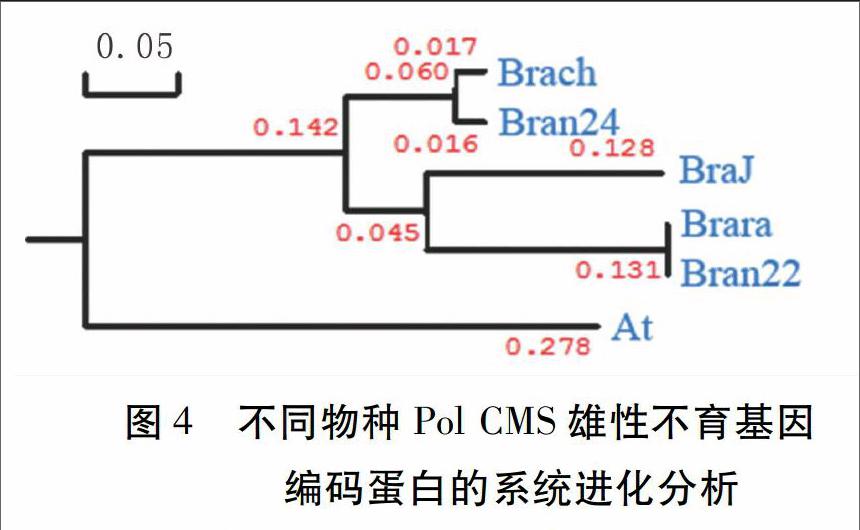
2.2.2 大白菜Pol CMS雄性不育基因Braraorf224的生物信息学分析 通过在GenBank中的BLAST检索,发现了两个甘蓝型油菜Pol CMS雄性不育基因,编码蛋白长度分别为222个氨基酸(Accession No. YP_717120)、224个氨基酸(Accession No. EU254235),分别命名为Bran22、Bran24。在青菜、芥菜中也发现了Pol CMS雄性不育基因,分别为青菜Brach (Accession No. AAT42256)、芥菜BraJ (Accession No. AAO48983)。拟南芥中的Pol CMS雄性不育基因蛋白为At(Accession No. NP_085529)。对来自不同物种的Pol CMS雄性不育基因编码的氨基酸序列进行生物信息学分析,发现大白菜Pol CMS雄性不育基因Braraorf224(Brara)编码的氨基酸序列与甘蓝型油菜Bran22的一致性为100%,与甘蓝型油菜Bran24的一致性为78%,与芥菜BraJ的一致性为73%,与青菜Brach的一致性为72%。但是,Brara与拟南芥At的一致性仅为46%,远远低于上述芸薹属物种(图3)。从进化树的结果来看,大白菜Brara 与油菜Bran22的亲缘关系最为接近,与拟南芥At亲缘关系最远(图4)。
2.3 大白菜Pol CMS 雄性不育系恢复基因BraraPPR的克隆及表达分析
2.3.1 大白菜Pol CMS 雄性不育系恢复基因BraraPPR的克隆 以大白菜Pol CMS 雄性不育恢复系早28花、花蕾总RNA的反转录产物为模板,对恢复基因BraraPPR进行PCR扩增,获得2 444 bp的条带,经测序为甘蓝型油菜Pol CMS恢复基因BrnPPR1 (Accession No. XM_013841641)的同源基因,命名为BraraPPR。BraraPPR基因编码区长1 953 bp,编码650个氨基酸,编码区内不含有内含子(图5)。
2.3.2 大白菜Pol CMS 雄性不育系恢复基因BraraPPR的生物信息学分析 通过GenBank中的BLAST检索,在甘蓝型油菜、甘蓝、亚麻荠、山葵、拟南芥中均发现有Pol CMS不育系恢复基因的同源基因,它们编码的蛋白质分别为油菜BrnPPR1(Accession No. XM_013841641)、油菜BrnPPR2 (Accession No. XM_013853446)以及甘蓝BroPPR (Accession No. XM_013744538 )、亚麻荠CsPPR (Accession No. XM_010460416 )、山葵EsPPR (Accession No. XM_006417174)、拟南芥AtPPR(Accession No. NM_101102)。利用DNAMAN软件对上述蛋白质的氨基酸序列进行比较分析,发现大白菜BraraPPR的氨基酸序列与油菜BrnPPR1、甘蓝BroPPR、油菜BrnPPR2、山葵EsPPR、亚麻荠CsPPR、拟南芥AtPPR的一致性分别为98%、77%、73%、73%、71%、70%(图6)。从进化树的结果来看,大白菜BraraPPR与油菜BrnPPR1的亲缘关系最为接近(图7)。
2.3.3 大白菜Pol CMS 雄性不育系恢复基因BraraPPR的表达模式分析 利用不育系92-09A、恢复系早28及杂种F1(92-09A×早28)的花、花蕾提取总RNA,以反转录获得的cDNA为模板,对Pol CMS 雄性不育系恢复基因BraraPPR进行qRT-PCR反应(图8)。结果表明,恢复基因BraraPPR在杂种F1花器中的表达量最高,其次是恢复系,在不育系中的表达量最低;杂种F1、恢复系中的表达量分别是不育系的4.9倍和3.1倍。
3 讨论与结论
3.1 Pol CMS雄性不育的转育及其利用
大白菜与甘蓝型油菜同属十字花科芸薹属,可以杂交结实。人们可以利用有性杂交将甘蓝型油菜Pol CMS雄性不育基因导入大白菜中。目前大白菜中使用的Pol CMS均来自于甘蓝型油菜。在本研究中,我们以柯桂兰馈赠的大白菜Pol CMS雄性不育KG-Pol植株为母本,通过多代回交转育,获得了大白菜Pol CMS雄性不育纯系92-09A。92-09A的花瓣、萼片和雌雄蕊都比保持系92-09小,花药锥形干瘪,不能形成有活力的成熟花粉粒。通过对大白菜Pol CMS雄性不育系92-09A的不育基因Braraorf224和恢复基因BraraPPR的序列分析,发现它们与甘蓝型油菜Pol CMS雄性不育基因Bran22和恢复基因BrnPPR1的氨基酸序列一致性分别为100%、98%。系统进化分析表明,Braraorf224与油菜Bran22亲缘关系最近, BraraPPR与油菜BrnPPR1亲缘关系最近。说明我们获得的大白菜Pol CMS雄性不育系92-09A的不育基因及其恢复基因来自于甘蓝型油菜。
在大白菜Pol CMS雄性不育转育过程中,我们发现不育株经常出现嵌合型、低温敏感型、高温敏感型不育。实践中常见同一株上可育/败育并存,低温表现不育, 温度升高后又转变为可育;高温下不育, 在通过低温影响后, 又变为可育。需大量筛选保持系, 才能育成稳定不育系。这与已有的报道结果相同[11,12]。因此,克服上述Pol CMS 雄性不育系的缺点是利用Pol CMS 雄性不育生产一代大白菜杂交种首先要面对的重要育种问题。为此,山东省农业科学院蔬菜花卉研究所分子育种研究室与山东登海种业股份有限公司西由种子分公司联合攻关,力争解决这一难题。目前,这方面的研究进展顺利,已获得了对高、低温不敏感的大白菜Pol CMS不育系。
虽然人们选育出了许多大白菜Pol CMS雄性不育系,但是真正利用Pol CMS雄性不育系大面积生产一代杂交种的例子尚不多见。造成这种情况的原因,除前面提到的Pol CMS的花粉败育易受温度影响不稳定外,还存在另外一个重要原因,就是目前人们主要利用大白菜的自交不亲和性和自交系内的迟配性来生产一代杂交种[2]。该方法制种可以获得正反交两个方面的一代杂交种子,产量高,是不育系制种法种子产量的1.3倍以上,同时纯度也可保持在95%以上。根据以上分析,我们认为综合利用大白菜Pol CMS雄性不育系、自交不亲合系、自交系(系内迟配性)选择适宜的亲本类型生产一代杂交种将是未来大白菜制种的发展方向。
3.2 大白菜Pol CMS雄性不育机理
随着人们对大白菜、油菜、萝卜等十字花科蔬菜线粒体、叶绿体基因组测序的完成[13-15],对各种雄性不育机理的研究也不断深入。到目前为止,人们对Pol CMS雄性不育机理研究结果主要有:(1)Pol CMS细胞质雄性不育直接与线粒体基因组(mtDNA)的差异、重排及其表达产物相关联,而与叶绿体基因组无关。mtDNA重排会导致新的嵌合基因产生或下游基因的表达模式和功能的改变,从而导致不育[16,17]。(2)Pol CMS雄性不育与线粒体中orf224/apt6转录结果有关。orf224/apt6有不同的转录起始位点,转录受恢复基因的影响[1, 18]。(3)蛋白质水平的作用与Pol CMS雄性不育有关,其中有能量蛋白假说和毒性蛋白假说[1,19]。上述研究结果仅能解释部分现象,要全面揭示Pol CMS不育机理还有更多的工作要做。比如,Pol CMS线粒体基因组全基因功能的研究及其与雄性不育性状的关联作用,Pol CMS线粒体基因突变及其基因工程技术研究等。
在本研究中,从大白菜Pol CMS雄性不育系92-09A中克隆得到的Braraorf224与甘蓝型油菜的雄性不育基因Bran22的氨基酸序列一致性为100%,说明大白菜Pol CMS雄性不育可能是由Braraorf224引起的。92-09A与早28杂交,获得的F1代杂交株是可育的,说明早28中携带有Pol CMS雄性不育的恢复基因。我们对恢复基因BraraPPR进行了克隆,并对其表达模式进行分析,结果表明,BraraPPR在恢复系早28及恢复系与雄性不育系杂交种F1中的表达量明显高于雄性不育系92-09A,说明大白菜Pol CMS雄性不育育性的恢复可能与BraraPPR有关,其分子机理仍需进一步深入研究。
参 考 文 献:
[1] Yamagishi H, Bhat S R. Cytoplasmic male sterility in Brassicaceae crops[J]. Breeding Science, 2014, 64: 38-47.
[2] 柯桂兰. 中国大白菜育种学[M]. 北京:中国农业出版社, 2010: 104-111.
[3] 傅廷栋, 杨光圣, 杨小牛,等. 甘蓝型油菜波里马细胞质雄性不育的发现、研究与利用[J]. 自然科学进展,1995,5(3):287-293.
[4] 徐文玲,王淑芬,王翠花,等. 大白菜Pol CMS育性恢复基因的表达分析[J]. 园艺学报,2013,40(8):1475-1486.
[5] 任成伟, 曹寿椿. 不结球白菜雄性不育材料选育及应用潜力研究[C]//江苏省首届青年学术会执行委员会主编. 江苏省首届青年学术年会论文集(农业分册). 北京:中国农业科学技术出版社, 1992: 144-149.
[6] 向长萍, 晏儒来, 李锡香. 紫菜薹雄性不育系的选育和应用[J]. 中国蔬菜, 2000(5):28.
[7] 柯桂兰,赵稚雅,宋胭脂,等.大白菜异源胞质雄性不育系CMS3411-7的选育及应用[J].园艺学报,1992,19(4):333-340.
[8] 杨晓云,张淑霞,张清霞.大白菜波里马胞质雄性不育系A7的选育及应用[J]. 山东农业科学,2006(5):11-13.
[9] Alexander M P. Differential staining of aborted and nonaborted pollen [J]. Stain Technology,1969, 44(3): 117-122.
[10]孙加顺. 我国大白菜育种取得突破性进展 极早熟大白菜新品种鲁白13号通过审定[J]. 现代农业,1997(7): 24.
[11]林超, 孙萍, 李兴福, 等. 利用细胞质雄性不育系生产萝卜一代杂交种[J]. 山东农业科学,2007(7):5-8.
[12]张德双, 曹明庆, 徐家炳. 大白菜转育新型甘蓝型油菜细胞质雄性不育系的研究[J]. 华北农学报, 2002,17(1):60-63.
[13]Park J Y, Kwon S J, Choi B S, et al. In silico-selection of Brassica rapa organelle genome-derived BACs using their end sequences and sequence level comparative analysis of the 124 kb mitochondrial genome sequences in the family Brassicaceae[J]. J. Crop Sci. Biotech.,2009,12(4):207-215.
[14]Hatje K, Kollmar M. A phylogenetic analysis of the Brassicales clade based on an alignment-free sequence comparison method [J]. Frontiers in Plant Science, 2012, 3(4): 192.
[15]Chen J, Guan R, Chang S, et al. Substoichiometrically different mitotypes coexist in mitochondrial genomes of Brassica napus L.[J]. PLoS ONE, 2011, 6(3): e17662.
[16]Erickson L, Grant I, Beversdorf W. Cytoplasmic male sterility in rapeseed (Brassica napus L.) [J]. Theoretical and Applied Genetics,1986, 72(2): 151-157.
[17]Singh M, Brown G G. Suppression of cytoplasmic male sterility by nuclear genes alters expression of a novel mitochondrial gene region [J]. Plant Cell, 1992, 3(12):1349-1362.
[18]袁美,杨光圣,傅廷栋,等.甘蓝型油菜pol CMS育性恢复基因对orf224/atp6的转录调控 [J].遗传学报,2003,30(5):469-473.
[19]夏广清,何启伟. 大白菜雄性不育及其机理的研究进展[J]. 山东农业科学,2004(6):73-76.
