拟南芥amiR-RUS4 ms35雄性不育双突变体的构建、鉴定及表型分析
2017-01-05董晶晶赵淑清
董晶晶,赵淑清
(山西大学生物技术研究所,化学生物学与分子工程教育部重点实验室,山西太原030006)
拟南芥amiR-RUS4 ms35雄性不育双突变体的构建、鉴定及表型分析
董晶晶,赵淑清
(山西大学生物技术研究所,化学生物学与分子工程教育部重点实验室,山西太原030006)
控制雄性育性是研究植物生殖和选择育种的一个重要目标。对雄性不育分子机制的深入理解将为控制雄性育性和杂种繁育提供有效途径。利用人工microRNA技术对DUF647蛋白家族成员RUS4基因进行特异沉默,结果发现,amiR-RUS4植株败育,药室内壁次生加厚异常,花药不开裂,表明RUS4与药室内壁的木质化有关。MS35/MYB26是木质素生物合成途径上游一个重要的激活因子,为了进一步了解RUS4和MYB26在调控花药药室内壁次生加厚过程中的作用,以ms35为母本、amiR-RUS4为父本进行人工杂交,得到杂交子2代(F2);并对杂交子2代(F2)的基因型、表型和基因表达进行了鉴定,结果获得一个纯合的amiR-RUS4 ms35双突变体,这为进一步分析RUS4和MS35在调控药室内壁次生加厚中的作用提供了宝贵的试验材料。
拟南芥;雄性不育;RUS4;MYB26
雄性不育是高等植物中较为普遍存在的一种遗传现象,是指植物雄性生殖器官不能产生正常功能的雄配子——花粉的现象,如植物花药中无花粉、花粉败育和花药不开裂等。雄性不育在农作物的杂种优势利用上具有重要的价值。近年来,人们对花药和花粉发育基本过程的认识有了很大的提高,其中,许多信息来自于对拟南芥雄性不育突变体的详细研究[1-4]。
花粉在植物雄性生殖器官花药中发育成熟,早期花药原基中造孢细胞通过一系列的细胞分裂、分化过程,形成小孢子母细胞和围绕在外面的绒毡层、中层和药室内壁3层营养组织,这3层营养组织和最外层表皮对小孢子母细胞起着重要的保护作用,同时为其发育成熟提供必要的物质和营养保障。当花药内部表皮、药室内壁、中层和绒毡层已经分化、减数分裂已经完成时,随即进入花粉成熟以及花药开裂进程。拟南芥花药开裂发生主要包括中间层和绒毡层的退化,内壁细胞的膨胀,在内壁细胞和连接细胞间的纤维层累积即次生壁加厚,隔膜退化形成二室花药,花药脱水,花粉粒膨大,最终导致花药开裂,释放花粉粒。其中,药室内壁次生加厚可为花药开裂提供一种机械力量[5]。对花药发育和花粉释放机制的深入研究将为控制作物雄性育性和杂种繁育提供有效的途径。
RUS4(ROOT UV-B SENSITIVE 4)是拟南芥DUF647(Domain of unknown function647)蛋白家族的一个成员[6]。通过人工microRNAs(artificial microRNAs,amiRNAs)技术获得了特异沉默RUS4(At2g23470)基因的突变体[7]。amiR-RUS4突变体由于自花授粉失败而产生短小的角果,但是通过人工授予有活力的花粉可以挽救其不育表型。研究结果显示,突变体的花粉活力降低,同时药室内壁次生加厚出现异常,导致花药开裂失败。
MYB26/MALE STERILE 35为R2R3型MYB转录因子,在花药开裂过程中具有重要作用[8]。MYB26功能异常导致植株败育,花药不开裂,但花粉具有活力[9-10]。研究发现,MYB26定位于细胞核,在花药药室内壁早期发育时开始表达,花粉有丝分裂和双核时期表达量达到最大,通过细胞特异的方式对花药药室内壁木质化和开裂起作用。MYB26过表达导致植株的异位次生加厚,在表皮组织表现尤为明显。MYB26可调控多种次生壁加厚相关基因的表达,包括IRREGULAR XYLEM1(IRX1),IRX3,IRX8和IRX12,且myb26突变体中NAC SECONDARY WALL THICKENING PROMOTING FACTOR1(NST1)和NST2的表达发生变化,说明MYB26可调控NST1和NST2的表达,可能位于木质素合成途径的上游,从而控制次生壁的木质化过程,推测MYB26通过NST1和NST2来调控IRX1,IRX3,IRX8,IRX12的表达[11]。
本研究以amiR-RUS4和ms35突变体为材料,进行人工杂交,以创制amiR-RUS4 ms35双突变体,对杂交子2代(F2)进行了基因型、表型分析,并对双突变体进行了表达分析,目的是得到纯合的amiR-RUS4 ms35双突变体,旨在为进一步分析RUS4和MYB26基因在调控药室内壁次生加厚过程中的关系奠定基础。
1 材料和方法
1.1 材料与生长条件
植物材料为野生型拟南芥 Col(Arabidopsis thaliana,Columbia ecotype),amiR-RUS4和ms35。其生长条件为:光周期16 h光照/8 h黑暗;相对湿度50%~70%;温度(22±2)℃。
1.2 方法
1.2.1 人工授粉杂交 选择ms35植株花瓣刚刚露白的花蕾,小心剥去花萼、花瓣和雄蕊,只留雌蕊,做好标记,作为母本。选择完全盛开的amiR-RUS4的花作为父本。由于amiR-RUS4突变体花药不开裂,需要人工释放花粉粒,并将其授到ms35植株的雌蕊柱头上,为保证成功,一般授粉2~3个花药。授粉3 d后观察,若杂交成功,可看到雌蕊伸长;若不成功,则萎蔫。15 d左右后收取单个角果,F1自交产生F2。
1.2.2 amiR-RUS4 ms35双突变体的鉴定 待F2植株生长15 d左右,取叶片于已配制的蔗糖缓冲液[12](50 mmol/L Tris-HCl,pH值 7.5,300 mmol/L NaCl,300 mmol/L蔗糖溶液)中充分研磨,99℃加热10 min,6 000 r/min短暂离心,单株提取基因组DNA,利用引物MIR319a-F/R(表1)进行基因组水平的PCR检测,以确定amiRNA前体的存在。纯合的ms35株系则通过表皮毛来鉴定,由于ms35突变体中MS35和GL基因均发生突变,而GL基因的缺失导致叶片缺少表皮毛,所以,可以通过叶片表皮毛的有无来鉴定纯合的ms35突变体。
1.2.3 表达分析 取经基因型和表型筛选出F2株系3,36,38号的花蕾,用TaKaRa MiniBEST Plant RNA Extraction Kit提取RNA,对RUS4和MYB26的表达进行RT-PCR检测(引物如表1所示),从而进一步鉴定纯合的amiR-RUS4 ms35双突变体。
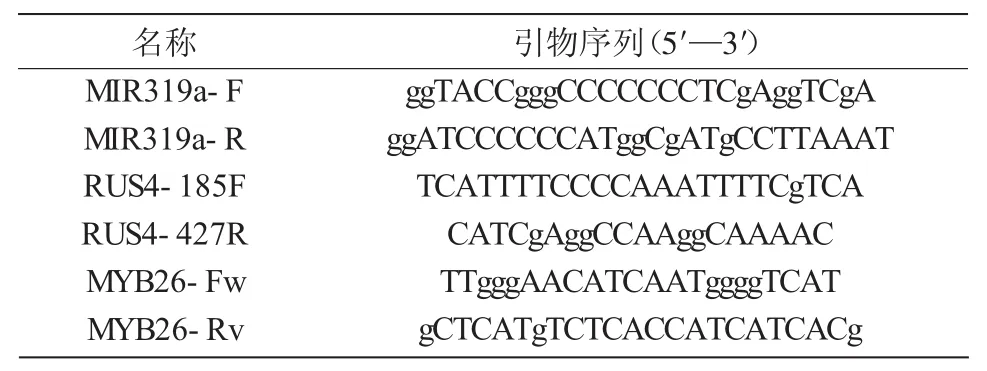
表1 amiR-RUS4 ms35植株鉴定的引物序列
1.2.4 表型分析 待F2植株生长50 d左右,观察植株生长状况、育性、叶片表皮毛、角果生长状况。
2 结果与分析
2.1 amiR-RUS4 ms35植株的基因型分析
amiR-RUS4突变体由人工microRNA敲减获得,其含有MIR319a前体核苷酸序列。种植amiRRUS4与ms35杂交F2共66株,用引物MIR319a-F和MIR319a-R在基因组水平进行PCR鉴定(以野生型Col作为对照),结果显示,除6,9,17,23,35,47,48,54,66号外,其余57株植株均有目的条带,且片段大小正确(图1),表明此57株植株中含有人工microRNA背景。
2.2 amiR-RUS4 ms35植株的表型分析
ms35来自Ler生态型,且该突变体中MS35和GL基因突变。GL基因编码一种类似MYB的蛋白质,该蛋白对于诱导表皮毛的发育是必需的,故ms35为表皮毛缺陷突变体,可作为表型观察的一个特征。观察66株中含有人工microRNA前体背景的F2植株表型发现,F2中3号植株无表皮毛,不结角果,其可能为纯合的双突变体,而36,38号植株育性降低且有表皮毛,其可能为杂合体(图2)。

2.3 amiR-RUS4 ms35植株的表达分析
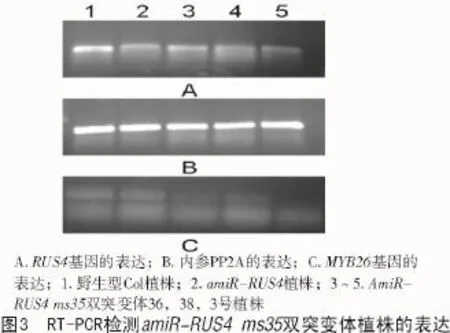
本研究对3,36,38号植株进行了RNA水平的半定量RT-PCR鉴定,结果显示,3号植株花蕾中RUS4,MYB26基因的表达明显下调,36,38号花蕾中RUS4表达降低不太明显,MYB26表达量减少(图3)。因此,杂交F2中3号植株为纯合双突变体。
3 讨论
拟南芥作为模式植物,对其雄性育性相关基因的功能分析有助于促进作物杂种优势的应用。由基因控制的可遗传的雄性不育突变体中花药和花粉发育出现异常的过程是研究的重点,它将雄性不育个体调控育性变化的研究和分子水平研究联系起来,为了解不育基因调控雄性不育的过程奠定了基础,对全面认识植物雄性育性的分子调控具有重要意义。
DUF647蛋白家族是广泛存在于真核生物体内高度保守的蛋白质家族,其在植物的生长发育过程中有着重要作用。拟南芥DUF647蛋白家族有6个成员:RUS1,RUS2,RUS3,RUS4,RUS5,RUS6。其中,RUS1和RUS2通过彼此相互作用,在根UVB感受途径中发挥作用[6,13]。且有研究表明,WRX3/ RUS1在生长素极性运输中有重要的调控作用[14]。RUS4作为DUF647蛋白家族成员之一,其是否与RUS1基因相类似在生长素运输途径中发挥功能,还有待于进一步分析。
本研究在实验室运用人工microRNA技术对RUS4进行了特异沉默,获得了amiR-RUS4基因敲减株,发现其花粉活力降低,药室内壁次生加厚异常,花药不开裂,为雄性不育株。ms35突变体中的MS35突变,使得花药无药室内壁木质化、花药开裂缺陷,但花粉具有活力;同时该突变体中GL基因突变,GL基因可调控表皮毛的发育,其突变会导致植株叶片缺乏表皮毛。本试验对amiR-RUS4和ms35杂交产生的F2植株进行了基因型、表型、表达分析,鉴定出了纯合的amiR-RUS4 ms35双突变体。突变体叶片缺乏表皮毛,含有microRNA前体序列,花蕾中RUS4和MYB26的表达明显下调,观察发现,药室内壁次生加厚异常,这为进一步分析RUS4和MYB26在调控花药药室内壁次生加厚中的作用提供了有价值的试验材料。
本研究通过吖啶橙/溴化乙锭染色发现,amiR-RUS4突变体药室内壁木质化减少,而ms35/ myb26突变体花药中药室内壁完全没有次生加厚,说明RUS4与MYB26均在药室内壁次生加厚和花药开裂过程中起作用。对amiR-RUS4 ms35纯合双突变体的鉴定发现,其植株表型和myb26相似,药室内壁次生加厚均有缺陷。通过表达分析发现,在amiR-RUS4突变体中,MYB26的表达与野生型相比无明显变化,说明RUS4功能缺失不会影响到MYB26的表达,推测MYB26可能位于RUS4基因的上游起作用或二者通过不同的途径发挥作用。MYB26在药室内壁木质化过程中,通过调控NST1和NST2来调节木质素合成相关基因IRX的表达,因此,通过对amiR-RUS4突变体中NST1/2的表达进行分析,也可推测RUS4与MYB26是否作用于同一途径。
花药药室内壁次生加厚在花药开裂过程中起关键性作用[8,15],此过程需植物激素(如生长素[16-17])和多种基因(如MYB26,SAF1[18],CBSX2[19],RPK2[20],RUS4)参与调控。尽管目前已经确定了药室内壁次生加厚过程的一些重要的调控因子,但是对于其具体调控机制仍不清楚,纯合amiR-RUS4 myb26双突变体的鉴定可为进一步研究RUS4基因调控花药药室内壁次生加厚的分子机制奠定一定的基础。
[1]Scott R J,Spielman M,Dickinson H G.Stamen structure and function[J].Plant Cell,2004,16(suppl):46-60.
[2]Ma H.Molecular genetic analyses of microsporogenesis and microgametogenesis in flowering plants[J].Annual Review Plant Biology,2005,56:393-434.
[3]Wilson Z A,Zhang D B.From Arabidopsis to rice:pathways in pollen development[J].Journal of Experimental Botany,2009,60(5):1479-1492.
[4]Shi J,Cui M,Yang L,et al.Genetic and biochemical mechanisms of pollen wall development[J].Trends in Plant Science,2015,20(11):741-753.
[5]Sanders P M,Bui A Q,Weterings K.Anther developmental defects in Arabidopsis thaliana male-sterile mutants[J].Sexual Plant Reproduction,1999,11(6):297-322.
[6]Leasure C D,Tong H,Yuen G,et al.ROOT UV-B SENSITIVE2 acts with ROOT UV-B SENSITIVE1 in a root ultraviolet B-sensing pathway[J].Plant Physiolgy,2009,150(4):1902-1915.
[7]李文超,赵淑清.人工microRNAs对拟南芥At1g13770和At2g23470基因的特异沉默[J].遗传,2012,34(3):348-355.
[8]Wilson Z A,Song J,Taylor B,et al.The final split:the regulation of anther dehiscence[J].Journal of Experimental Botany,2011,62(5):1633-1649.
[9]Dawson J,Sozen E,Vizir I,et al.Characterization and genetic mappingofa mutation(ms35)which prevents anther dehiscence in Arabidopsis thaliana by affecting secondary wall thickening in the endothecium[J].New Phytologist,1999,144(2):213-222.
[10]Steiner-Lange S,Unte U S,Eckstein L,et al.Disruption of Arabidopsis thaliana MYB26 results in male sterility due to non-dehiscent anthers[J].Plant Journal,2003,34(4):519-528.
[11]Yang C,Xu Z,Song J,et al.Arabidopsis MYB26/MALE STERILE35 regulates secondary thickening in the endothecium and is essential for anther dehiscence [J].Plant Cell,2007,19(2):534-548.
[12]Kenneth Berendzen,Iain Searle,Dean Ravenscroft,et al.A rapid and versatile combined DNA/RNA extraction protocol and its application to the analysis of a novel DNA marker set polymorphic between Arabidopsis thaliana ecotypes Col-0 and Landsbergerecta [J].Plant Methods,2005,1(1):4.
[13]TongH Y,Leasure CD,Hou XW,et al.Role ofroot UV-Bsensing in Arabidopsis early seedling development[J].PNAS,2008,105(52):21039-21044.
[14]Yu H,Karampelias M,Robert S,et al.ROOT ULTRAVIOLET BSENSITIVE1/WEAK AUXIN RESPONSE 3 is essential for polar auxin transport in Arabidopsis[J].Plant Physiolgy,2013,162(2):965-976.
[15]樊新平,牛西午.拟南芥花药和花粉发育的基因调控[J].山西农业科学,2013,41(5):506-510.
[16]Cecchetti V,Altamura M M,Costantino P,et al.Auxin controls Arabidopsis anther dehiscence by regulating endothecium lignification and jasmonic acid biosynthesis[J].The Plant Journal,2013,74(3):411-422.
[17]Cecchetti V,Altamura MM,Falasca G,et al.Auxin regulates Arabidopsis anther dehiscence,pollen maturation,and filament elongation[J].Plant Cell,2008,20(7):1760-1774.
[18]Kim Y Y,Jung K W,Jeung J U,et al.A novel F-box protein represses endothecial secondary wall thickening for anther dehiscence in Arabidopsis thaliana[J].Plant Physiology,2012,169(2):212-216.
[19]Jung K W,Kim Y Y,Yoo K S,et al.A Cystathionine-β-Synthase Domain-Containing Protein,CBSX2,regulates endothecial secondary cell wall thickening in anther development[J].Plant Cell Physiology,2013,54(2):195-208.
[20]Mizuno S,Osakabe Y,Maruyama K,et al.RPK2 is a novel factor controlling anther development in Arabidopsis thaliana[J].Plant Journal,2007,50(5):751-766.
Construction,Identification and Phenotypic Analysis of amiR-RUS4 ms35 Male Sterile Double Mutant in Arabidopsis thaliana
DONG Jing-jing,ZHAO Shu-qing
(Key Laboratory of Chemical Biology and Molecular Engineering of Ministry Education,Institute of Biotechnology,Shanxi University,Taiyuan 030006,China)
Controlling male fertility is an important goal for the research of plant reproduction and selective breeding.A better understanding of the male sterile molecular mechanisms will provide effective ways for the control of male fertility and for hybrid generation. Specific silencing of a DUF647-containing gene RUS4 by artificial microRNA led to a severe reduction of male fertility.The amiR-RUS4 anther displayed altered endothecium secondary thickening,which impacted on anther dehiscence.Transcription factor MYB26/MS35 played a regulatory role in endothecium lignification as wall thickening was not observed in the endothecial cells of the MYB26 mutant and displayed indehiscent anthers.This study generated an amiR-RUS4 ms35 double mutant where homozygous ms35 lines were used for crosses with amiR-RUS4 plants.The genotypic,phenotypic and gene expression analysis were performed in the F2generation.One homozygous amiR-RUS4 ms35 double mutant were obtained,which will provide a valuable material for further analysis of the relationship between RUS4 and MS35 in the regulation of secondary thickening in the endothecium.
Arabidopsis thaliana;male sterile;RUS4;MYB26
Q943
A
1002-2481(2016)05-0583-04
10.3969/j.issn.1002-2481.2016.05.04
2016-01-26
国家自然科学基金项目(31170273);太原市科技明星专项(11014902)
董晶晶(1990-),女,山西交城人,在读硕士,研究方向:植物分子生物学。赵淑清为通信作者。
