长白山沿海拔梯度臭冷杉径向生长对气候变化的响应
2016-12-19覃鑫浩何友均许娜子
覃鑫浩,何友均,许娜子
(1.国家林业局调查规划设计院,北京 100714;2.中国林业科学研究院林业科技信息研究所,北京 100091)
长白山沿海拔梯度臭冷杉径向生长对气候变化的响应
覃鑫浩1,何友均2,许娜子2
(1.国家林业局调查规划设计院,北京 100714;2.中国林业科学研究院林业科技信息研究所,北京 100091)
为了研究树木生长对气候变化的响应,选取长白山北坡3个海拔高度建立了臭冷杉Abies nephrolepis的年轮差值年表。通过年表特征及响应函数分析,探讨不同海拔臭冷杉径向生长对气候变化的响应。结果表明,温度是影响海拔分布上限(1 760 m)臭冷杉生长的主要影响因子,年轮宽度指数与上年9月和当年5月的平均温呈显著正相关,与降水没有显著关系;在中海拔(1 250 m),年轮宽度指数与温度和降水都没有呈现出显著相关关系;在海拔分布下限(780 m),降水是主要的影响因子,年轮宽度指数与上年9月和10月的降水量呈显著正相关,与温度没有达到显著相关。未来气候变化有利于臭冷杉的径向生长,有利于长白山暗针叶林生产力的增加。
树木年轮;气候变化;分布极限;径向生长
20世纪以来,全球的温度和降水发生了变化。其中,全球增温显著,1901~2012年平均气温上了0.72 [0.49 to 0.89] ℃[1],而21世纪后半期的增温更加明显,1951到2012年的增温速率为0.12[0.08 to 0.14] ℃/10年。接下来的20年(2016~2035),温度预计还会增加0.3~0.7 ℃,而到了2081~2100年,温度增加会在0.3~4.8 ℃[1]。相对于温度,1901年以来的平均降水量只是略有增加[2]。在中国东北部长白山地区,1900年以来温度上升了2 ℃,其中1950年以来温度上升更为显著(1 ℃以上)。到21世纪末,该地区温度将上升 3.8℃[3-4]。
气候变化将显著影响树木生长,进而影响到种群发展乃至森林生态系统,尤其在物种分布的生理极限区,这种影响更加明显[5]。沿海拔变化,影响树木生长的气候因子会随着改变。在海拔分布上限,低温往往是限制树木生长的主要因子[6]。在海拔分布下限,温度对树木生长的限制作用减弱,而降水变得更为重要[7]。
长白山臭冷杉Abies nephrolepis的海拔分布广泛(750~1 800 m),是该地区,特别是暗针叶林(1 100~1 800 m)的主要树种,在维持森林生态系统稳定方面发挥着重要的作用。在长白山地区,许多学者利用年轮资料研究了不同海拔红松Pinus koraiensis[8]、鱼鳞云杉Picea jezoensis[9]和岳桦Betula ermanii[10]径向生长对气候变化的响应,发现了树木生长与气候因子的显著关系,但是关于臭冷杉生长在海拔梯度上对气候变化响应的研究还处于空白。本文在3个海拔高度研究臭冷杉径向生长与气候因子的关系,完善了该地区的年轮资料。同时,由于不同树种对气候变化的响应存在差异,本文结果也是对研究该地区影响树木生长主要气候因子的有利补充。
1 材料与方法
1.1 研究区概况
长白山位于中国东北部(41°31′~ 42 °28′N,127°9′~ 128°55′E)。气候特征为冬季寒冷漫长,夏季凉爽湿润。年均温3.6 ℃,其中1月温度最低,年均温-15.4 ℃,7月温度最高,年均温19.6 ℃。平均年降水量为707 mm,主要集中在5~9月(75%)(见图1)。森林类型随海拔升高依此变化为阔叶红松林(750~1 100 m),主要树种有红松,紫椴Tilia amurensis、水曲柳Fraxinus manshurica、蒙古栎Quercus mongolica、白桦Betula platyphylla和五角枫Acer mono等;暗针叶林(1 100~1 800 m),主要树种有鱼鳞云杉、臭冷杉和落叶松Larix olgensis等;岳桦林(1 800~2100 m),主要树种有岳桦和落叶松[11]。土壤类型变化为深棕色森林土、山地棕色针叶林土和暗棕色森林土。
1.2 样本采集及年表建立
2008年,我们在长白山北坡选取3个海拔高度(780 m、1250 m和1760 m),每个海拔点选择20~25株干形通直、无损伤和病虫害的臭冷杉,每株树在胸高(1.3 m)处从东西两个方向上采集两颗年轮样芯,放入塑料管中并编号。样点概况如表1所示。

图1 长白山1960~2008年逐月温度和降水Fig.1 Monthly mean temperature and precipitation in Changbai Mountain during 1960~2008

表1 采样点基本情况Table 1 Characteristics of study sites
将样芯带回实验室,按照树木年轮学的标准工作流程,先将样芯进行黏贴固定在凹槽内,在阴凉通风处凉干后,用不同目数的砂纸进行打磨直至样芯表面光滑、年轮清晰可见。年轮宽度利用Lintab5树轮测量仪在0.001的精度下进行测量。利用COFECHA[12]程序检验定年的准确性,剔除不能交叉定年的年轮序列。
利用ARSTAN[13]程序建立各海拔的年表。由于树木生长趋势受年龄以及外界干扰等因素影响,我们运用采用步长为2/3序列长度的样条函数对所有年表进行去趋势,之后再利用自回归模型方法得到包含较多高频气候信息的差值年表,用以分析年轮宽度指数与气候因子的关系。
1.3 气候数据
本研究选用二道白河气象站(42°43′N,128°12′E,海 拔 700 m)的 器 测 数 据(1960~2008年),利用Kendall检验[14]和double-mass检验[15]对气候数据进行了均一性检验,检验结果表明温度和降水资料稳定,变化均一。我们选取月平均温和月降水量这两个气候因子进行年轮宽度指数与气候因子相关性分析。
1.4 数据分析
为了分析不同海拔间树木生长特征,将1950~2007的年轮宽度转化为胸高断面积生长量,用单因素方差分析比较3个海拔间臭冷杉平均胸高断面积的差异,从而比较树木生长速率。未选择1950年以前的数据,是为了去除年龄对生长量的影响。
气候对树木生长具有滞后效应,因此选取上年7月到当年9月的逐月平均温和月总降水量与差值年表进行相关性分析。利用树轮软件Dendroclim2002中的响应函数来分析年轮指数与气候因子的关系[16]。
2 结果与分析
2.1 树木生长特征
单因素方差分析表明,在1950~2007时间段内,中海拔(1 250 m)的平均胸高断面积(1.9 cm2)显著高于(p<0.001)高海拔(0.58 cm2)和低海拔(0.59 cm2)的平均胸高断面积,说明中海拔地区的水热条件最好,适合于臭冷杉的生长。另外,所有海拔臭冷杉近50年来的胸高断面积呈增加趋势(图2)。
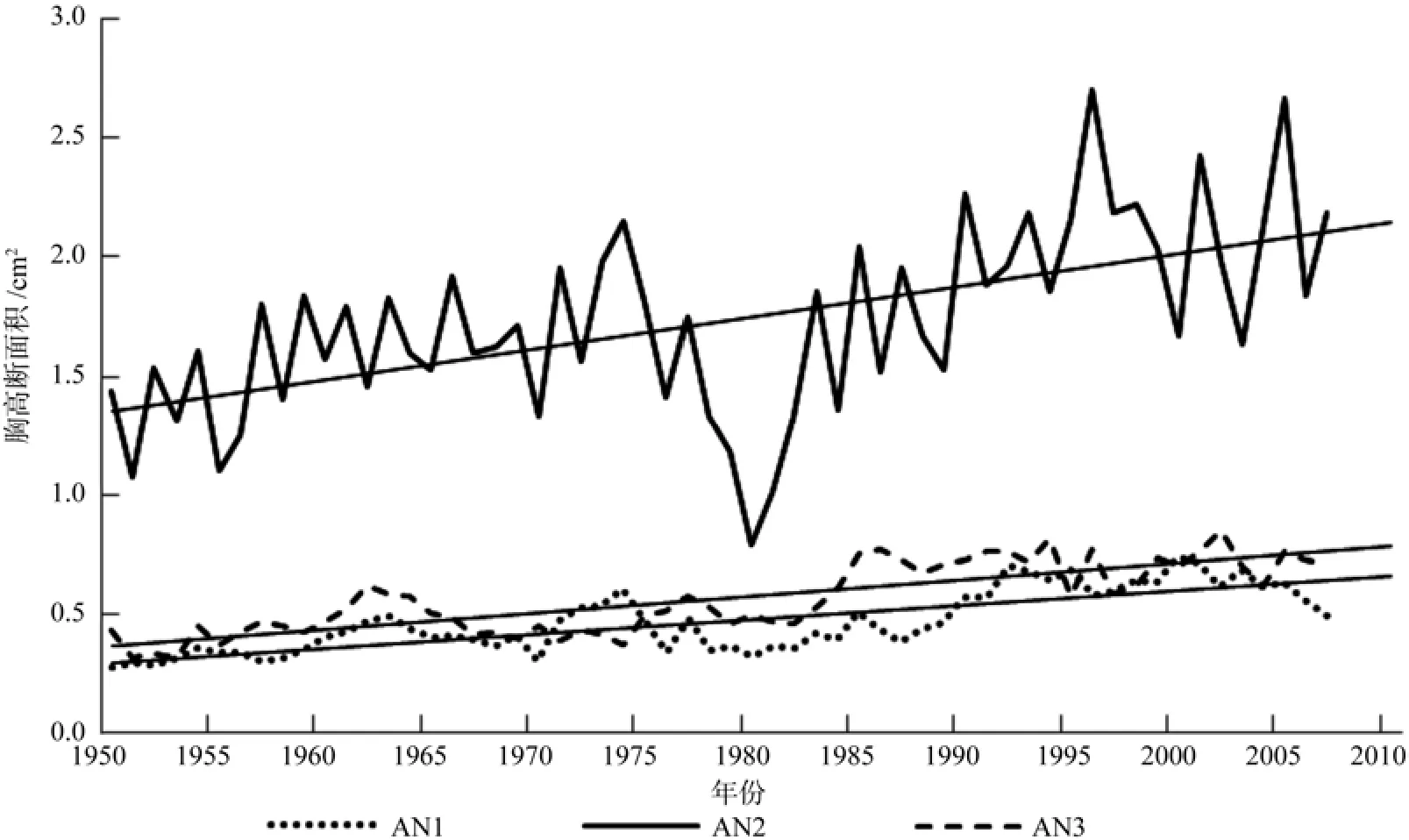
图2 3个海拔高度臭冷杉胸高断面积的变化(1950~2007)Fig. 2 Changes in basal area increment of A. nephrolepis at three altitudes during the period 1950~2007
2.2 年表统计特征
差值年表的统计量如表2所示。海拔上限(1 760 m)年表的平均敏感度、第一特征向量、信噪比和样本总体代表性均高于其它两个海拔(780 和1 250 m)的年表,说明海拔上限的树木包含更多的环境信息,对气候变化更为敏感。3个样点树与树之间的相关系数均高于0.3,说明树木之间的相关性较好,能够反映环境变化的一致性。3个样点的样本总体代表性均超过了85%,说明臭冷杉适合于年轮气候学的研究。
2.3 树木生长与气候因子的关系
臭冷杉差值年表与平均气温和降水量的相关分析表明,温度是影响臭冷杉海拔上限径向生长的主要影响因子,降水则是其海拔下限生长的限制因子(图3)。在海拔分布上限,臭冷杉年轮宽度指数与上年9月和当年5月的温度呈显著正相关,而与降水没有显著相关性;在中海拔,臭冷杉生长没有与温度和降水表现出显著相关关系;在海拔分布下限,上年10月和当年9月的降水积极影响臭冷杉的径向生长。

表2 臭冷杉差值年表统计特征及公共区间(1960~2008)分析Table 2 Statistics of residual chronologies of A. nephrolepis and common interval analysis

图3 树轮宽度差值年表与逐月气候因子的相关分析Fig.3 Response function analysis between the residual chronology and the monthly climatic factors
3 结论与讨论
臭冷杉径向生长对气候变化的反应随海拔变化而变化,从海拔下限的降水为主要影响因子,到中海拔的没有相关性,再到海拔上限的温度为主要影响因子,说明在海拔分布的极限区,树木生长对气候变化更为敏感。本研究的结果也与传统的年轮学观点相符[17],温度和降水分别是海拔上、下限树木生长的限制因子。
在海拔分布下限,臭冷杉径向生长主要受上年10月和当年9月降水的影响,即生长季末期降水的增加有利于树木生长。上年10月降水的增加,可以促进生长季末期碳水化合物的增加和积累,从而有利于来年的生长[18],这也和以往该地区臭冷杉低海拔树轮研究的结果一致[7]。当年9月的降水能够提高树木中的水分,从而增强光合作用形成宽轮[19]。在中海拔,雨热条件最有利于树木的生长,温度和降水均不成为树木生长的限制因子,因而没有呈现出显著相关关系。在海拔分布上限,上年9月和当年5月的温度增加有利于宽轮的形成。上年9月的增温可以提高光合作用的效率,在生长季末期促进有机物的积累,从而有利于来年树木的生长[20]。5月温度的提高在3个方面促进树木的生长[21-22],(1)温度升高,可以促进地面冰雪的融化,为早期树木的生长提供充足的水分;(2)温度升高,可以使得土壤早日解冻,促进根系活动,并同时促进开花和展叶,从而延长了生长季;(3)温度升高,可以提高叶面积和光合作用效率,从而促进生长初期的生长。这也与长白山相邻的日本亚高山冷杉的年轮研究结果一致[23],说明在东北亚地区生长季初期的温度对冷杉树种的径向生长起着关键作用。
本文利用树木年轮学的方法研究了臭冷杉沿长白山海拔变化径向生长对气候变化的响应,阐明了不同海拔影响臭冷杉径向生长的主要气候因子,为该地区的气候重建提供了理论依据。但受气候因子获取手段的限制,只对温度和降水与树木生长关系进行了分析,但干旱指数等对树木生长也有显著影响,因而将来应获取更多的气象资料与年轮指数进行相关性分析,从而更加全面的阐述影响该区域树木生长的气候因子。
[1] IPCC, 2013: Summary for Policy makers. In: Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change [Stocker, T.F., D. Qin, G.-K. Plattner,M. Tignor, S.K. Allen, J. Boschung, A. Nauels, Y. Xia, V.Bex and P.M. Midgley (eds.)]. Cambridge University Press,Cambridge, United Kingdom and New York, NY, USA.
[2] IPCC, 2007: Climate Change 2007: Impacts, Adaptation, and Vulnerability. Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change [Parry, M. L., Canziani, O. F., Palutikof, J. P., van der Linden, P. J. and Hanson, C. E. 7–22. UK: Cambridge University Press. Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change.
[3] Zhai P M, Zhang X B., Wan H, et al. Trends in total precipitation and frequency of daily precipitation extremes over China[J].Journal of Climate, 2005, 18: 1096-1108.
[4] Xu Y L., Huang X Y., Zhang Y, et al. Statistical analyses of climate change scenarios over China in the 21st Century[J].Advances in Climate Change Research, 2006, 2: 50-53.
[5] Peñuelas J, Boada M. A global change-induced biome shift in the Montseny mountains (NE Spain)[J]. Global Change Biology,2003, 9: 131-140.
[6] 彭剑锋,勾晓华,陈发虎,等.阿尼玛卿山地不同海拔青海云杉树轮生长特征及其对气候的响应[J].生态学报,2007,27(8):3268-3276.
[7] Zhang Y, Bergeron Y, Gao L S, et al. Tree growth and regeneration dynamics at a mountain ecotone on Changbai Mountain, Northeastern China: which factors control species distributions?[J]. Ecoscience, 2014, 21: 1-18.
[8] 陈 力,尹云鹤,赵东升,等.长白山不同海拔树木生长对气候变化的响应差异[J]. 生态学报, 2014, 34(6): 1568-1574.
[9] Yu D P, Wang Q L, Wang G G, et al. Dendroclimatic response of Picea jezoensis along an altitudinal gradient in Changbai Mountains[J]. Science in China Series E, 2006, 49: 150-159.
[10] 王晓明,赵秀海,高露双,等.长白山北坡沿海拔梯度岳桦径向生长对气候的响应[J]. 应用与环境生物学报,2013,19(6):929-934.
[11] 张 赟,张春雨,赵秀海,等.长白山次生林乔木树种空间分布格局[J].生态学杂志,2008, 27(10): 1639-1646.
[12] Holmes R L. Computer assisted quality control in tree-ring dating and measurement[J].Tree-Ring Bulletin, 1983, 43: 69-75.
[13] Cook E R, Holmes R L. Users manual for ARSTAN [M]. Tucson:Laboratory of Tree-ring Research, University of Arizona, 1986.
[14] Kendall M G, Gibb7ons J D. Rank Correlation Methods [M].London: Edward Arnold, 1990.
[15] KornerC.,PaulsenJ.A world-wide study of high altitude tree line temperatures[J]. Journal of Biogeography, 2004, 31: 713-732.
[16] Biondi F, Waikul K. DENDROCLIM2002: a C++ program for statistical calibration of climate signals in tree-ring chronologies[J].Computers and Geosciences, 2004,30(3):303-311.
[17] Fritts H C, Smith D G, Cardis J W, et al. Tree-ring characteristics along a vegetation gradient in northern Arizona[J]. Ecology,1965, 46: 393-401.
[18] Liang E Y., Shao X M., Hu Y X, et al. Dendroclimatic evaluation of climate-growth relationships of Meyer spruce (Picea meyeri)on a sandy substrate in semi-arid grassland, north China[J]. Trees Structure and Function, 2001, 15: 230-235.
[19] Ettl G J., Peterson D L. Growth response of subalpine fi r (Abies lasiocarpa) to climate in the Olympic Mountains, Washington,USA[J]. Global Change Biology, 1995, 1: 213-230.
[20] 王 婷,于 丹,李江风,等.树木年轮宽度与气候变化关系研究进展[J].植物生态学报, 2003, 27(1): 23-33.
[21] Peterson D W, Peterson D L. Effects of climate on radial growth of subalpine conifers in the North Cascade Mountains[J].Canadian Journal of Forest Research, 1994, 24: 1921-1932.
[22] 曹受金,曹福祥,项文化.利用树木年轮研究湖南炎陵气温变化情况——1840年以来5~7月份气温变化情况重建[J].中南林业科技大学学报,2012,32(4):10-14.
[23] Takahashi K, Okuhara I, Tokumitsu Y, et al. Responses to climate by tree-ring widths and maximum latewood densities of two Abies species at upper and lower altitudinal distribution limits in central Japan. Trees Structure and Function, 2001, 25: 745-753.
Response of radial growth of Abies nephrolepis to climate change along an elevational gradient in Changbai Mountain
QIN Xin-hao1, HE You-jun2, XU Na-zi2
(1. State Forsetry Bureaue Survey Scheme Designing Institute, Beijing 100714, China; 2. Research Institute of Forsetry Policy and Information, Chinese Academy of Forsetry, Beijing 100091, China)
We established residual chronologies of Abies nephrolepis at three altitudes in Changbai Mountain to study responses of tree growth to climate change. According to analyses of chronology characteristics and response function, we detected responses of radial growth of A. nephrolepis to climate change along the elevational gradient. The results showed that temperature is the main factor affecting tree growth at the upper limits (1760 m), by presenting signi fi cant and positive correlations between ring-width index and mean temperature in previous September and current May. At the middle distribution (1250 m), neither temperature nor precipitation significantly affected radial growth of A. nephrolepis. At the lower distributional limits (780 m), precipitation mainly affected tree growth, ring-width index was signi fi cantly and positively correlated with precipitation in previous September and October. Based on the current prediction of future climate change in Changbai Mountain, the radial growth of A. nephrolepis would bene fi t from changes in temperature and precipitation, promoting productivity of spruce- fi r forest in the area.
dendrochronology; climate change; distributional limits; radial growth
S718.55+4
A
1673-923X(2016)11-0092-05
10.14067/j.cnki.1673-923x.2016.11.016
2016-03-07
国家自然科学基金(31570633&31170593)、国家林业局天然林保护评估项目(500103-1303)
覃鑫浩,工程师,硕士研究生 通讯作者:何友均,研究员,博士,硕士生导师;E-mail:hyjun163@163.com
覃鑫浩,何友均,许娜子.长白山沿海拔梯度臭冷杉径向生长对气候变化的响应[J].中南林业科技大学学报,2016,36(11): 92-96.
[本文编校:吴 毅]
