贵州西部光皮桦天然次生林生物循环特征
2016-12-19赵文君丁访军周凤娇刘延惠崔迎春
赵文君,丁访军,周凤娇,刘延惠,吴 鹏,崔迎春
(贵州省林业科学研究院,贵州 贵阳 550005)
贵州西部光皮桦天然次生林生物循环特征
赵文君,丁访军,周凤娇,刘延惠,吴 鹏,崔迎春
(贵州省林业科学研究院,贵州 贵阳 550005)
以贵州西部林龄10 a的光皮桦天然次生林为研究对象,对林分养分元素积累与分配及生物循环特征进行了研究,结果表明:光皮桦天然次生林各组分营养元素含量总的序列是草本层>树叶>凋落物层>灌木层>枝>根>干,乔木层中各元素含量为N>K>P;营养元素(N、P、K)总积累量为512.04 kg/hm2,乔木层、林下层植被、凋落物层分别占46.12%、32.66% 和21.22%,乔木层养分积累量较低;养分年吸收量、归还量、留存量分别为94.52、74.80、19.72 kg/(hm2·a),循环系数、利用系数和周转时间分别为0.79、0.40和3.16 a,循环速率依次为N>K>P。该林分循环速率快,周转期短,利于土壤肥力维持,利于林地生产力的发展,在生产实践中合理经营和管理光皮桦林分,对森林生态系统的营养循环及维持和改善林地土壤肥力有着积极的意义。
生物循环;林分养分;光皮桦;天然次生林
光皮桦Betula luminifera,系桦木科Betulaceae桦木属Betula阔叶落叶大乔木,材质优良,用途广泛,生长较快,适应性强,是中国特有的优良速生用材树种,是中亚热带荒山造林先锋树种和珍贵的生态树种[1]。贵州是光皮桦较为集中的分布区[2],光皮桦对喀斯特地区保持水土、涵养水源有重要的作用,对长江、珠江上游生态屏障有重要的意义。
养分元素生物循环是森林生态系统中重要的功能过程之一[3],研究生物循环规律有助于认识生态系统物质循环机制,不仅对林分生产力及生态系统连续性与稳定性的提高有重要的意义,而且有利于揭示森林经营对林地土壤肥力的影响[4],进而指导生产实践,调节和改善各种限制因素以实现加速养分的循环利用速率和最大限度地提高森林生产力的目的[5-6]。目前不同学者对不同森林生态系统生物循环进行了较全面的研究,如田大伦、项文化、肖兴翠等[7-9]对比研究了马尾松林和湿地松林养分循环特征及不同年龄阶段、不同密度调控与养分循环的关系,文仕知等[10]研究了桤木人工林营养元素的季节动态与生物循环规律,张希彪等[11]研究了黄土丘陵区油松人工林与天然林的生物循环特征,王仲等[12]对喀斯特城市森林生物循环进行了研究等,但对喀斯特地区适生的光皮桦林生物循环研究还较缺乏,为此,本研究在光皮桦生物量和净生产力[13]研究的基础上,对贵州西部光皮桦天然次生林养分含量、积累、分布和生物循环等进行研究,以期揭示养分元素的特征和循环规律以及其对土壤肥力的影响,为光皮桦天然次生林的保护与管理提供理论依据和积累基础数据。
1 研究区概况
六盘水玉舍林场坐落在贵州省西部水城县境内, 地 理 位 置 为 104°47'813″ ~ 104°50'385″E,26°27'508″ ~ 26°27'881″N,属乌蒙山支脉,地处珠江水系上游,最高海拔2 503 m,最低海拔1 700 m,相对高差803 m。林场年均温13~14 ℃,1月均温3.0~6.3 ℃,7月均温19.8~22.0 ℃,无霜期230~300 d,降水量1 200~1 500mm。土壤类型以黄壤为主。场内人工林和原生植被交错分布,人工林以华山松Pinus armandii、杉木Cunninghamia lanceolata、柳杉Cryptomeria fortunei为主,天然林主要是光皮桦和十齿花Dipentodon sinicus,森林覆盖率达到80%以上。本研究光皮桦次生林是森林受破坏后经10多年封育保护后天然形成的。林内灌木主要有悬钩子Rubus corchorifolius、杜鹃Rhododendron simsii、铁线莲Clematis fl orida、金丝桃Hypericum monogynum、菝葜Smilax china、木姜子Litsea cubeba等,草本植物主要有禾本科Poaceae、菊科Asteraceae、唇形科Labiatae、紫堇科 Fumariaceae、毛茛科Ranunculaceae及蕨类Pteridophyta 植物等[14]。
2 研究方法
2.1 样地的设置与调查
样地设置在六盘水玉舍林场,2009年9月到10月期间,经实地踏勘后选取立地条件相近的30 m×30 m光皮桦林分3块,采取每木检尺获取林分相关参数,其基本情况见表1。
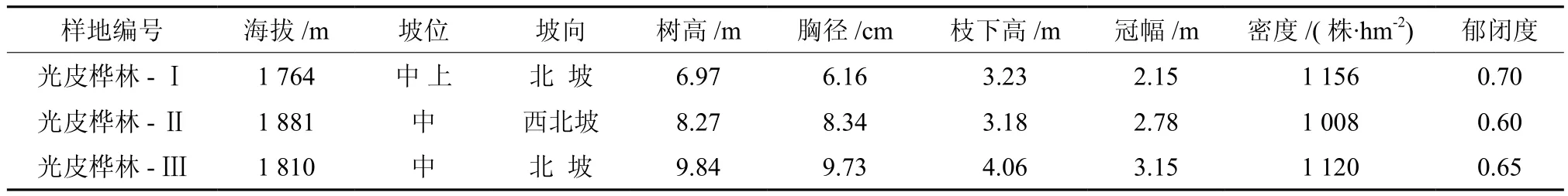
表1 样地基本情况Table 1 Basic information of sampling plots
2.2 生物量测定及生产力估算
生物量测定及生产力估算方法参见文献[13]。另外,在每个固定样地内设置1 m×1 m的尼龙网收集器3个(共9个),用于年凋落物归还量的确定,2009年9月至2010年9期间,每月收集凋落物1次,测定其干质量后推求。
2.3 样品的采集及化学分析
在测定生物量的同时对乔木层的干、枝、叶和根分别取样,灌木层、草本层的地上部分和地下部分分别取样,枯枝落叶层以未分解层和半分解层分别取样。样品粉碎过筛后,在80℃下烘干至恒质量,然后进行各元素化学分析,全N采用半微量凯氏法测定,全P采用钼锑抗比色法测定,全K采用火焰光度法测定。
3 结果与分析
3.1 养分元素含量
光皮桦林养分元素含量见表2。就各组分而言,草本层地上部分和乔木层树叶中营养元素含量最高,约为含量最低树干的9倍。就营养元素而言,草本层、灌木层中K元素含量最高,分别是N的1.6倍和1.4倍、P元素的6.2倍和3.7倍,乔木层和凋落物层则以N元素含量占绝对优势,分别是K的2.0倍和13.6倍、P元素的6.5倍和16.1倍,均以P元素含量最低。就林分各层次而言,灌草层的养分含量较高,普遍高于乔木层除树叶之外的其他组分含量,尤以K元素最为突出。凋落物层养分含量占整个林下层养分含量的39%左右,可见光皮桦天然林中灌草层、凋落物层在林分养分循环占有重要的地位。各组分营养元素含量总的序列是草本层>树叶>凋落物层>灌木层>枝>根>干。
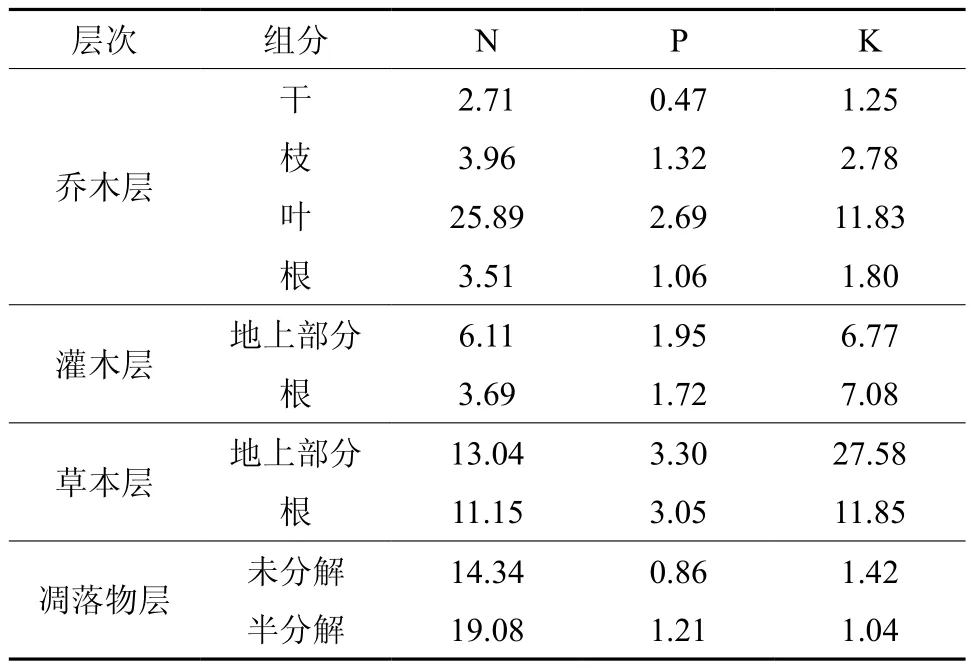
表2 光皮桦天然次生林各组分养分元素含量Table 2 Nutrient concentrations in different components of Betula luminifera natural secondary forest (g·kg-1)
3.2 营养元素积累量和分配
3.2.1 养分元素积累量及空间分布
由表3可知,乔木层营养元素的积累量为236.18 kg/hm2,表现为枝(34.15%)>干(33.74%)>叶(18.31%)>根(13.81%),可见各组分中树干、树枝的养分储量较大,根、叶的积累量较小,是因为各组分营养元素的差异除与元素含量有关外,更主要的是由于不同组分生物量的差异造成的[15]。如叶中N元素含量为25.89 g/kg,是树干中含量的9.5倍,而叶中N元素的积累量为27.7 kg/hm2,仅是树干积累量的0.57倍。
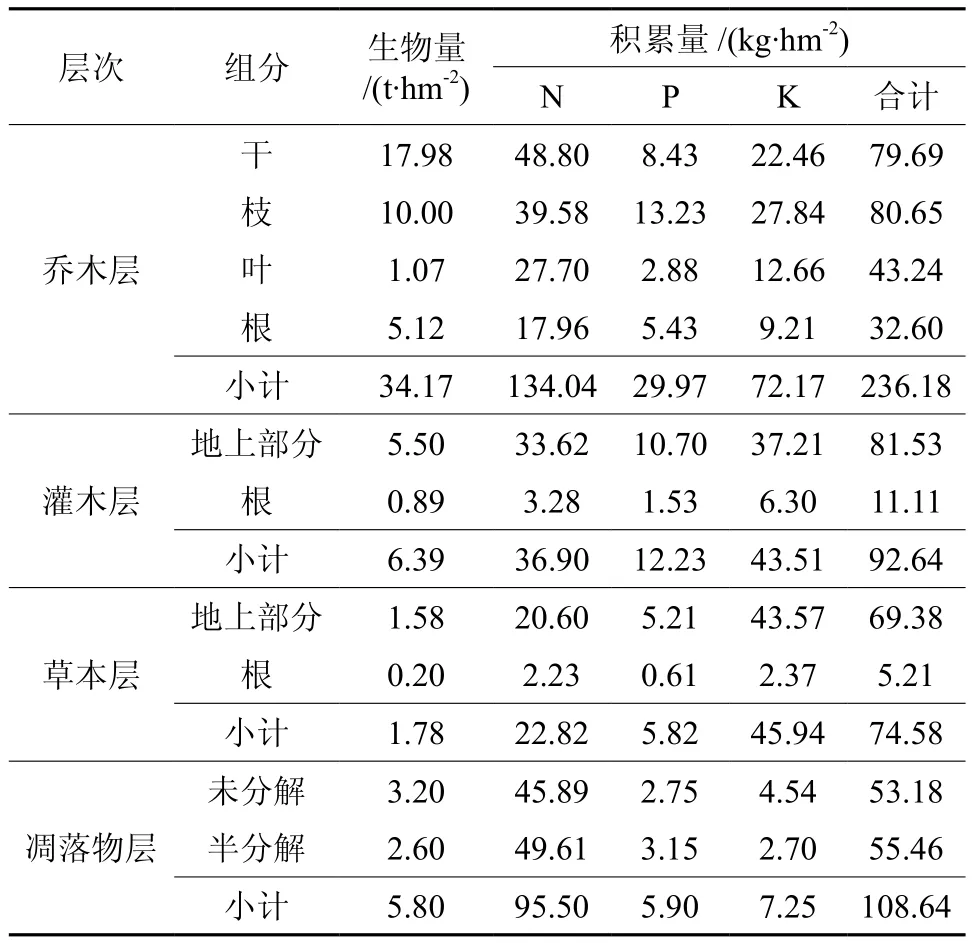
表3 光皮桦天然次生林主要营养元素的积累量Table 3 Nutrient element accumulation of Betula luminifera natural secondary forest
林下层养分积累量为275.86 kg/hm2,其中灌木层为92.64 kg/hm2,草本层为74.58 kg/hm2,凋落物层为108.64 kg/hm2,其序列为凋落物层>灌木层>草本层,灌、草层地上部分营养元素含量是根含量的7.3倍和13.3倍。
各层次营养元素的积累量,乔木层、凋落物层N>K>P,灌草层K>N>P,表现出群落不同层次养分利用的不同,这也是群落稳定性的养分基础。
该林分营养元素空间分布见表4,总积累量为512.04 kg/hm2,分配比例为乔木层占46.12%,灌木层占18.09%,草本层占14.57%,凋落物层占21.22%。乔木层生物量占70.98%,相对应养分储量仅占46.12%,还不到总储量1/2,可见在整个生态系统中乔木层养分积累量很低。林下植物层生物量占16.97%,而对应的养分储量占到32.66%,可见林下层对养分富集力强,在养分循环中也占有绝对的比例,具有不可忽视的作用。凋落物层养分含量占21.22%,与养分归还量密切相关,是林分实现养分持续供给的重要途径[16]。
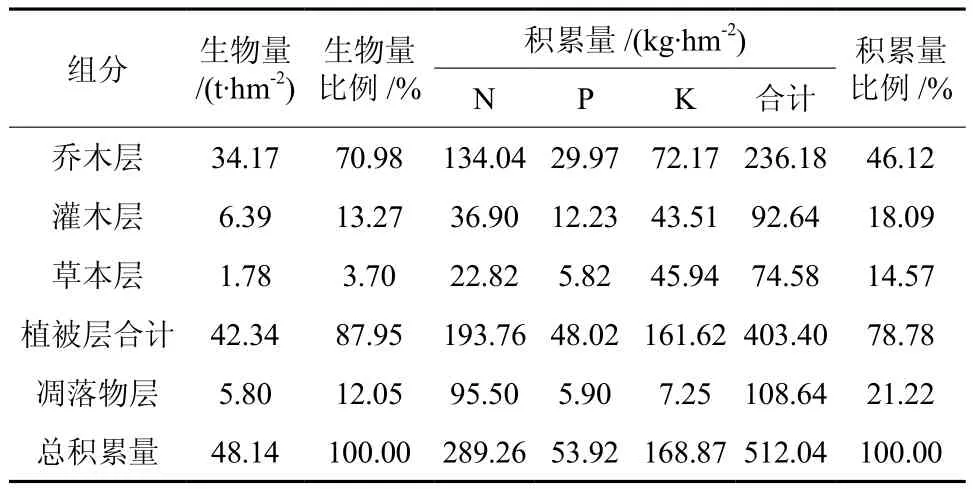
表4 光皮桦天然次生林分养分空间分布Table 4 Nutrient spatial distribution of Betula luminifera natural secondary forest
3.2.2 养分元素年积累量
乔木层主要营养元素年积累量为64.17 kg/hm2(见表5),其序列为叶>干>枝>根,叶的年积累量占了相当大的比重,约为69.2%,这是落叶阔叶林较针叶林循环快的原因之一。同一组分中3种营养元素的年积累量与总积累量大小顺序相同,表现为N> K > P,这与各元素的生理特性及树种对不同元素的需求性有关[11,17]。
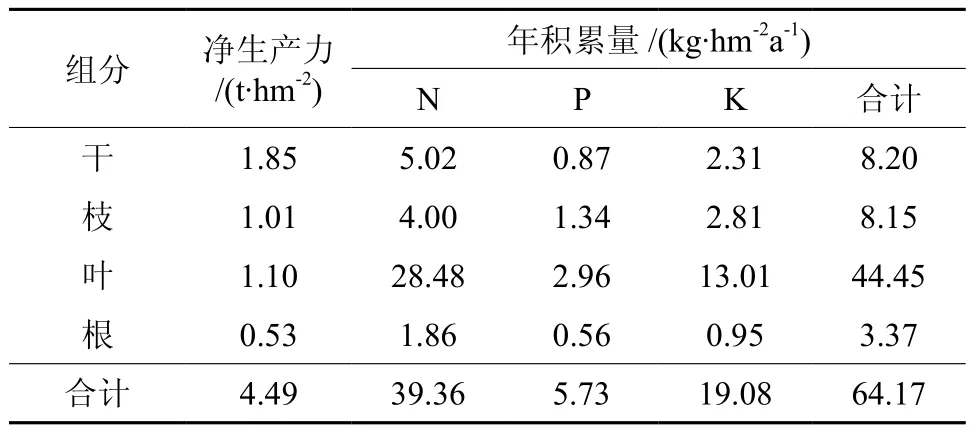
表5 天然光皮桦林主要元素的年积累量Table 5 Nutrient annual accumulation of Betula luminifera natural secondary forest
3.3 光皮桦天然次生林养分的生物循环
营养元素的生物循环能揭示森林土壤和植物间养分元素的周期性变化,包括吸收、存留和归还3个环节,以“吸收量=留存量+归还量”这种生物循环平衡来表述[11,18]。本研究归还量仅计算凋落物中的养分量,所以比实际值要小,光皮桦为落叶阔叶树种,树叶为当年形成,当年归还,所以将其列入归还量中,留存量中予以不计。由表6可知,光皮桦天然林总归还量为74.80 kg/(hm2·a),N、P、K归还量分别为 55.22 、4.61 、14.98 kg/(hm2·a);年留存量为 19.72 kg/(hm2·a),N、P、K留存量分别为 10.88 、2.77、6.07 kg/(hm2·a);天然光皮桦林总吸收量为 94.52 kg/(hm2·a),N、P、K养分的吸收量分别为 66.10、7.38、21.05 kg/(hm2·a),可见年归还量远大于留存量,归还量占吸收量的79.13%,且以N的吸收量和归还量最高。
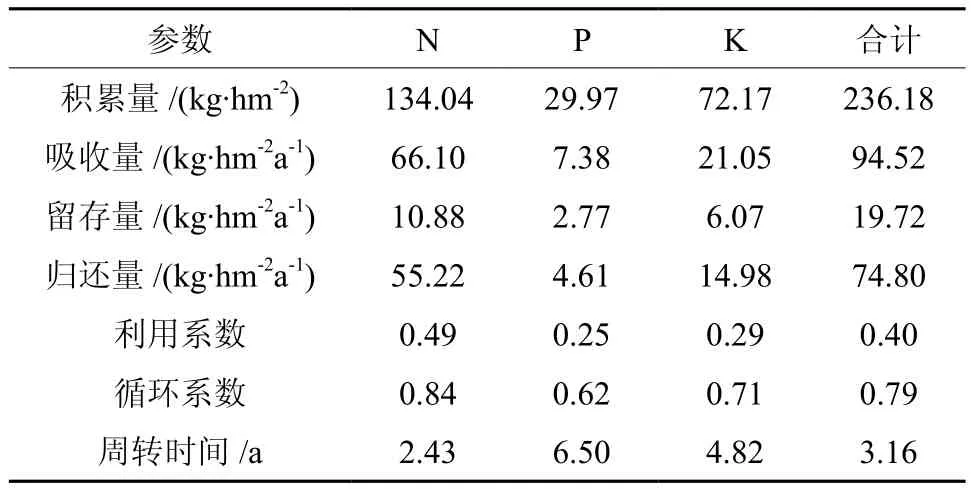
表6 天然光皮桦林养分循环参数Table 6 Biological cycling characteristic parameters of nutrient element in Betula luminifera natural secondary forest
通常用循环系数、周转时间、利用系数反映生态系统的养分循环特点[19]。养分循环系数为0.79,大小排序为N>K>P。循环系数越大,生态系统元素循环的速率越快,系统中存留量的比例就越小,林木生长对土壤库存元素的耗费也就越小[20]。N的流动性最大,说明光皮桦林分对土壤中N消耗最小;P则相反,对地力的消耗需求最大。周转时间平均为3.16 a,3种元素以N的最短,大约2.43 a;P的最长,为6.50 a。利用系数为0.40,3种元素的大小排序为N>K>P。总体来看,N循环系数最大,周转时间最短,说明光皮桦天然次生林N利用较高,循环较快;P循环系数最低,周转时间最长,需要的养分最多。
4 结论与讨论
光皮桦乔木层各组分养分含量序列为叶>枝>根>干。叶是光合作用的器官,植物生长过程中代谢活跃,需要大量的养分参与同化作用,叶还作为植物生长的养分库,为其他器官形成发育提供营养,因此其养分含量最高。Oliveira等[21]对Quercus suber的研究发现叶达到养分浓度高峰时,茎的养分浓度相对恒定,但随后叶和茎都转为养分库。因此枝的养分含量仅次于叶。而干的生理及代谢活动最弱,需要的养分少,因而养分含量也最少。各元素含量大小顺序均为N>K>P,N的含量最高,充分显示了N在植物生长中的重要作用,是森林生产力的主要限制因子之一[22]。各组分养分积累量为枝>干>叶>根;年积累量为叶>干>枝>根;各元素积累量及年积累量大小顺序均为N>K>P,植物体内各组分生理活性不同及不同组分间生物量的差异是导致植物体内不同组分养分贮量差异的主要原因[17]。
与南方常见的其他林分相比,光皮桦天然次生林乔木层营养元素积累量比较低。如同为桦木科10 a林龄的桤木林[10],其乔木层各营养元素积累量(kg/hm2)分别为 364.45 (N)、40.07(P)、121.67(K);广西14年林龄的马尾松林[7]乔木层积累量(kg/hm2)分别为 365.71 (N)、23.63(P)、143.74(K);湖南10年林龄的杉木林[23]乔木层积累量(kg/hm2)分别 为332.41(N)、26.02(P)、158.53(K)。林分营养元素的积累量是树种、林分密度、林龄、林地肥力等多种因素相互影响的结果[24]。加之,本研究光皮桦天然次生林是在森林破坏后荒山灌丛之上逐渐恢复形成的,属于低生物量森林群落[13],也是造成光皮桦天然次生林养分积累量较低的原因之一。
林下层植被养分积累量与乔木层相比含量相对较高,尤其K元素积累量高于乔木层。林下层占整个林分系统的32.66%,该比例远远高于其他林分系统(多数在20%以内),如马占相思林[25]为6.2%~13.74%,马尾松林[7]为11.75%,桤木林[10]为20.42%。可见光皮桦林下层植被对养分的富集力很强,主要是因为光皮桦天然次生林散生性强,郁闭度不高,有着适宜的林下环境,林下植被物种多样性丰富,生长迅速,对养分的需求量大的缘故。且林下层植被积累的养分归还又较快,在养分循环中具有不可忽视的作用,加强对林下层植被的管理对光皮桦林分养分循环有积极意义。同时也显示乔木层的养分积累量有一定的提升空间,如通过适当增加林分密度或合理的改造林分结构(营造光皮桦-杉木针阔混交林[1]),可实现林分乔木层养分积累量的提高,因为在一定的的密度范围内养分积累量随着林分密度的增大而增大[9,26]。
光皮桦林年吸收量、归还量、留存量分别为94.52、74.80、19.72 kg/(hm2·a)。10 年生桤木林[10]吸收量、归还量分别为 324.67、257.8 kg/(hm2·a),4.5年生厚荚相思[28]吸收量、归还量分别为179.46、79.88 kg/(hm2·a),14年生的马尾松林[17]吸收量、归还量分别为 136.82、98.74 kg/(hm2·a),10年生杉木林[24]吸收量、归还量分别为164.07、83.34 kg/(hm2·a),与以上速生树种相比,其吸收量、归还量较低,说明光皮桦适应性较强,较低的营养物质量就能满足其生长需求。其循环系数为0.79,利用系数为0.40,周转时间为3.16 a,循环系数高于(等于)桤木林(0.79)、厚荚相思(0.45)、马尾松林(0.72)、杉木林(0.51)[10,27,7,23]等大部分森林群落,说明光皮桦生长迅速,元素循环周转快,归还率大,对土壤库存元素的耗费小,利于土壤肥力的维持,这也是光皮桦能较好适应南方石漠化瘠薄土壤环境的原因之一,可以作为石漠化地区治理的优选树种。本研究光皮桦森林平均林龄10 a,处于迅速生长发育阶段,加之其凋落物的高N量和较大的归还量,可以推测随着林分的发展其仍会维持较高的归还速率及较短周转时间,林地土壤循环将处于良性循环状态,土壤肥力一定程度上会呈现进展趋势,利于林分生产力的发展,因为林分在达到成熟期前随着林龄增加,其归还速率快,周转期短,有利于林地生产力维持[8,11]。综上在生产实践中合理经营和管理光皮桦林分,对森林生态系统的营养循环及维持和改善林地土壤肥力有着积极的意义。
[1]曹健康,杨新虎,万乐金.光皮桦杉木混交林效应研究初报[J].黄山学院学报,2007,9(3):77-80.
[2]杨安敏,曾亚军,邓伯龙,等.贵州桦木资源现状与开发利用策略[J].资源与环境,2010,26(5):438-440.
[3]Chapin F S. The mineral nutrition of wild plants[J]. Ann. Rer .Ecol. Syst. , 1980, 12(1): 233-260.
[4]杨丽丽,文仕知,何功秀.长沙市郊枫香人工林微量元素分布与生物循环特征[J]. 中南林业科技大学学报,2011,31(12):85-91.
[5]刘世荣.兴安落叶松人工林生态系统营养元素生物地球化学循环特征[J]. 生态学杂志,1992,11(5):1-6.
[6]刘增文,李雅素.黄土残塬沟壑区刺槐人工林生态系统的养分循环通量与平衡分析[J].生态学报,1999,19(5):632-637.
[7]田大伦,项文化,闫文德.马尾松与湿地松人工林生物量动态及养分循环特征[J].生态学报,2004,24(10):2207-2210.
[8]项文化,田大伦.不同年龄阶段马尾松人工林养分循环的研究[J].植物生态学报,2002,26(1):89-95.
[9]肖兴翠,李志辉,唐作钧,等.林分密度对湿地松人工林养分循环速率和利用效率的影响[J].生态学杂志, 2013, 32(11):2871-2880.
[10]文仕知,黄采艺,杨丽丽,等.桤木人工林营养元素的季节动态、空间分布与生物循环研究[J].水土保持学报,2012, 26(6):96-101.
[11]张希彪,上官周平.黄土丘陵区油松人工林与天然林养分分布和生物循环比较[J].生态学报, 2006,26( 2):373- 382.
[12]王 仲,田大伦,宁晓波,等.喀斯特城市森林微量元素生物循环的研究[J]. 中南林业科技大学学报,2013,33(7):107-113.
[13]高艳平,潘明亮,丁访军,等. 贵州西部光皮桦天然次生林生物量和净生产力的研究[J].中南林业科技大学学报,2012,32(4): 55-60.
[14]高艳平,丁访军,潘明亮,等.贵州西部光皮桦天然次生林碳素积累及分配特征[J].南京林业大学学报,2014, 38(4):51-56.
[15]何池全,赵魁义.毛果苔草湿地营养元素的积累、分配及其生物循环特征[J].生态学报, 2001,21(12):2075-2080.
[16]刘文飞,樊后保,高春芬,等.连续年龄序列桉树人工林凋落物量及养分通量[J].生态学杂志,2009,28(10):1928-1934.
[17]刘广全,土小宁,赵士洞.秦岭松栎林带生物量及营养元素的生物循环特征研究[J].林业科学,2001,37(1):28-36.
[18]Duvigneaud P .温带落叶林矿质元素的生物循环[M].陈佐忠译.植物生态学译丛第1集.北京:科学出版社,1974:72-95.
[19]夏 菁,魏天兴,陈佳澜,等.黄土丘陵区人工养分循环特征[J].水土保持学报,2010,24(3):89-93.
[20]罗 辑,程根伟,李 伟,等.贡嘎山天然林营养元素生物循环特征[J].北京林业大学学报,2005,27( 2):13-17.
[21]Oliveira G, Martins-Loução M A, Correia O,et al. Nutrient dynamics in crown tissues of cork-oak (Quercus suberL.) [J].Trees, 1996, 10(4):247-254.
[22]陈伏生,曾德慧,何兴元.森林土壤氮素的转化与循环[J].生态学杂志,2004,23(5):126-133.
[23]田大伦,盘宏华,康文星,等.第二代杉木林养分动态研究[J].中南林学院学报,2001,21(3):6-12.
[24]莫江明,Sandra Brown,孔国辉,等.鼎湖山马尾松林营养元素的分布和生物循环特征[J].生态学报,1999,19(5):635-640.
[25]何 斌,秦武明,余浩光,等.不同年龄阶段马占相思人工林营养元素的生物循环[J].生态学报,2007,27(12):5158-5167.
[26]赵广亮,王继兴,王秀珍.油松人工林密度与养分循环关系的研究[J].北京林业大学学报,2006,28(4):39-44.
[27]秦武明,何 斌,覃世赢,等.厚荚相思人工林营养元素生物循环的研究[J].水土保持学报,2007,21(4):103-108.
Biological cycling of nutrient element ofBetula luminiferanatural secondary forests in western Guizhou province, China
ZHAO Wen-jun, DING Fang-jun, ZHOU Feng-jiao, LIU YAN-hui, WU Peng, CUI Ying-chun
(Guizhou Forestry Academy, Guiyang 550005, Guizhou, China)
Nutrient accumulation, distribution and biological cycling of the main elements of theBetula luminiferanatural forest of 10 years was studied in western Guizhou Province. The results show that the nutrient concentrations in different components ofBetula luminiferanatural forest was in the order of herbal> leaf> litter> shrub> branch> root> stem, the order of nutrient elements concentration was N > K > P in arbor. The total nutrient accumulation was 512.04 kg/hm2, of the total nutrient accumulation 46.12% was distributed in arbor layer, 32.66% at undergrowth vegetation layer ,and 21.22% at litter layer, and the arbor layer nutrient accumulation was lower. The amount of uptake, return and retention of nutrient were 94.52,74.80,19.72 kg/(hm2·a), respectively. The cycling coef fi cient, utilization coef fi cient and turnover period were 0.79, 0.40, 3.16a, respectively. And the cycling speed was N>K>P.Recycling rate was higher and turn over period was shorter, it will bene fi t to the development of the forest land productivity. We should strengthen operation and management ofBetula luminiferanatural forest, it will make sense to nutrient cycle of forest ecosystem and to maintain and improve forest soil fertility.
biological cycle; forest nutrient;Betula luminifera; natural secondary forests
S718.55
A
1673-923X(2016)09-0114-05
10.14067/j.cnki.1673-923x.2016.09.021
2015-04-16
国家公益性行业专项子项目(201204101-5);黔科合人才团队[2014]4004号
赵文君,助理研究员;E-mail:236135040@qq.com
赵文君,丁访军,周凤娇,等. 贵州西部光皮桦天然次生林生物循环特征[J].中南林业科技大学学报,2016,36(9):114-118.
[本文编校:谢荣秀]
