团头鲂谷胱甘肽S-转移酶基因的克隆及其在氨氮胁迫中的表达分析
2016-12-06孙盛明朱健戈贤平张成锋缪凌鸿张武肖章琼
孙盛明,朱健,*,戈贤平,张成锋,缪凌鸿,张武肖,章琼
1.中国水产科学研究院淡水渔业研究中心,无锡214081
2.南京农业大学无锡渔业学院,无锡214081
团头鲂谷胱甘肽S-转移酶基因的克隆及其在氨氮胁迫中的表达分析
孙盛明1,朱健1,*,戈贤平1,张成锋1,缪凌鸿1,张武肖2,章琼2
1.中国水产科学研究院淡水渔业研究中心,无锡214081
2.南京农业大学无锡渔业学院,无锡214081
谷胱甘肽S-转移酶(glutathione S-transferase,GST)是一类多功能蛋白家族,主要参与解毒和抗氧化防御过程。为了研究GST在团头鲂(Megalobrama amblycephala)肝脏解毒过程中的作用,克隆并分析了团头鲂1个谷胱甘肽S-转移酶基因(命名为MaGST)cDNA序列,采用实时荧光定量PCR研究了其在氨氮胁迫下的表达规律。MaGST包含1个长218个氨基酸的完整开放阅读框,具有GST蛋白家族的保守碱基和保守结构域。通过MEGA 5.0软件分析系统进化树发现,团头鲂GST与其他动物mu型GST聚为一簇,表明团头鲂GST属于mu型GST。荧光定量PCR结果显示MaGST基因在团头鲂各组织中均有表达,在肝脏和鳃中表达量最高,肌肉中表达量最低,同时在氨氮胁迫过程中该基因在肝和鳃中的表达规律相似,均在胁迫期间表达量显著上调;氨氮胁迫24 h时鳃和肝组织均存在组织损伤。研究结果提示在团头鲂肝脏和鳃组织中GST基因参与了氨氮胁迫的解毒过程。将该基因的编码区重组到pET-21(a+)载体后在大肠杆菌中得到诱导表达,重组MaGST的GST活力为(10.36±0.68)U·mg-1蛋白。
氨氮胁迫;团头鲂;谷胱甘肽S-转移酶;基因克隆
团头鲂(Megalobrama amblycephala)是我国大宗淡水鱼类主要养殖品种之一。然而,随着团头鲂集约化养殖密度和规模的扩大,其发病率明显增高,而水体中氨氮含量过高是促使其发病的主要影响因素之一,严重制约了团头鲂高密度养殖模式的发展[1]。已有研究表明氨氮胁迫对鱼类的影响主要集中在生长性能[2-3]、鳃增生[4]和氧化损伤脂质和蛋白质[5]等几个方面,而目前有关团头鲂对水环境氨氮胁迫的去毒分子机理、代谢机制等研究尚未见报道。
水生生物解毒过程被分为3个阶段,阶段Ⅰ与阶段Ⅱ能够将外源性化学物质转化成毒性较小的水溶性代谢物,阶段Ⅲ在机体细胞内消除这些代谢产物[6]。谷胱甘肽S-转移酶(glutathione S-transferase, GST,EC.2.5.1.18)作为主要的第二相解毒酶(phaseⅡdetoxification enzyme)参与了外源性或内源性毒素的解毒过程,它可以催化亲核性的谷胱甘肽与各种亲电子外源化学物的结合反应,参与维持了机体内氧化还原状态的平衡,从而提高机体对环境的适应能力以及成活率[7]。文献报道GST参与细胞耐菌性、植物抗除草剂和昆虫抗药性等分子过程[8-10],上述研究表明GST在动植物解毒、抗逆和抗病过程中起着重要作用。
谷胱甘肽S-转移酶是一组广泛分布于各类生物细胞的超家族酶类[11],哺乳动物中目前报道的GST主要分为细胞质GSTs(包括7个类型Alpha、Mu、Pi、Theta、Sigma、Omega和Zeta)、线粒体GST (kappa)和微粒体GST[12]。鉴于鱼类中抗氧化分子如谷胱甘肽或抗氧化物酶如谷胱甘肽S-转移酶能够保护机体氧化损伤,近年来鱼类不同亚型的GST基因cDNA全序列已相继被克隆[13-15],而关于草食性鱼类团头鲂GST基因的克隆与表达研究未见报道。本研究以实验室前期构建的团头鲂全组织cDNA文库中筛选出的EST序列为基础,通过RACE技术获得了团头鲂谷胱甘肽S-转移酶基因的全长序列,采用荧光定量PCR技术分析该基因在团头鲂氨氮胁迫下的表达模式。采用体外重组表达技术获得了谷胱甘肽S-转移酶基因的重组表达产物,检测了重组蛋白的活性,以期为团头鲂功能基因开发及解毒系统研究提供技术支撑。
1 材料与方法(Materials and methods)
1.1 实验材料
1.1.1 实验鱼
团头鲂幼鱼取自中国水产科学研究院淡水渔业研究中心南泉养殖基地。选择活泼健康、规格一致的团头鲂幼鱼(15±0.86)g共300尾,试验开始前用基础饲料驯养2周,每天投喂3次(08:00、11:30和17:00),根据摄食情况适当调整投喂量,实验前提前2天停止投喂饲料。整个试验期间水质如下:水温为23~24℃,溶氧≥5mg·L-1,pH为6.8~7.2。通过本实验室预实验得出团头鲂幼鱼96 h氨氮半致死浓度的基础上[16],设计低氨氮组(对照组0.46mg·L-1)和高氨氮组(25mg·L-1),每个处理设置3个平行,用氯化铵为母液配制实验组的总氨氮浓度。挑选体色正常、健康的团头鲂幼鱼随机放入6个可控温循环流水圆形蓄养槽(规格为φ820 mm×700 mm)内,每桶40尾。实验期间停止喂食,每隔4小时进行总氨氮浓度的测定。
1.1.2 试剂和仪器
RNAiso Plus、3’-Full RACE Kit、5’-Full RACE Kit、LA Taq TM、pMD 18-T Vector、PrimeScript RT reagent Kit、SYBR Premix Ex Taq TM、DH5α感受态细胞购自TaKaRa;分析纯NH4Cl、苯酚氯仿异戊醇(25:24:1)、氯仿、异丙醇、DEPC水、Tryptone、Yeast extract、NaCl、Agar、氨苄青霉素、SanPrep柱式DNA胶回收试剂盒购自Sangon Biotech;引物由上海生工合成。低温离心机(湘仪TGL-16M高速台式冷冻离心机)、凝胶成像系统(Beckman Coulter CEQ 8000)、紫外分光光度计(eppendorf BioPhotometer)、PCR仪(eppendorf Autherized Thermal cycler)、荧光定量PCR仪(Applied Biosystems 7500 Real Time PCR System)、摇床(华利达HZ-9511K恒温振荡器)、荧光显微镜(奥林巴斯)、德国徕卡切片机(Leica RM 2135)。
1.2 实验方法
1.2.1 总RNA的提取
取团头鲂肝组织转入液氮预冷的研钵中,边磨边加入液氮,迅速将组织研磨成粉末状,RNA提取方法按照RNAiso Plus的操作步骤进行组织裂解后用传统的酚仿抽提法提取总RNA并使用RNA-free DNaseI纯化试剂盒用于各组织总RNA的纯化,紫外分光光度计测定RNA浓度及纯度并通过琼脂糖凝胶电泳检测各组织总RNA的完整性。
1.2.2 GST3’端和5’端扩增
从本实验室构建的团头鲂肝脏cDNA文库的测序结果中,通过拼接组装及比对分析,从中发现1条EST序列(长度为756 bp)与鲤鱼(Cyprinus carpio)的GST(ABD67509.1)氨基酸序列相似度较高,根据已知中间序列,采用 Primer 5.0软件分别设计5′RACE和3′RACE的特异性引物(详见表1),以团头鲂肝脏cDNA为模板,按照3'RACE和5'RACE试剂盒(Takara,大连)说明书分别进行3’和5’的扩增。
1.2.3 序列分析
使用ContigExpress软件将团头鲂GST的中间序列、3’测序结果、5’测序结果进行拼接得到该基因的全长cDNA序列。在Compute pI/Mw程序中预测蛋白分子量及等电点(http://web.expasy.org/compute _pi/)。登陆NCBI,在该网站中的ORF Finder程序中(http://www.ncbi.nlm.nih.gov/gorf/gorf.html)查找所拼接团头鲂GSTcDNA序列的开放阅读框及其所对应的氨基酸序列,同时对开放阅读框进行Blastp比对,运用DNAMAN软件进行氨基酸多序列比对分析,利用MEGA 5.0软件构建系统进化树。采用NCBI的Conserved Domains(http://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)查找工具分析蛋白质结构域。
1.2.4 团头鲂不同组织GST的表达分析
提取团头鲂肌肉、鳃、肾、脑、肝、肠道6个组织的总RNA,荧光定量PCR反应的cDNA合成按照TaKaRa公司的PrimeScriptTMRT reagent Kit试剂盒说明书进行反转,根据所获得的GST基因全长cDNA序列设计1对荧光定量特异引物MaGST qF和MaGST qR(见表1),扩增长度为151 bp;以团头鲂的β-actin基因(序列号AY170122)为荧光定量反应的内参基因,设计1对内参引物β-actin-F和β-actin-R (见表1)。对荧光定量基因特异性引物和内参引物进行普通PCR扩增,用琼脂糖检测其在团头鲂中的扩增情况,判断目的片段的大小并检测其正确性。

表1 基因克隆所用的引物及其序列Table 1 Primers and their sequences used in gene cloning
荧光染料使用Takara的SYBR®Premix Ex Taq II,反应在Applied Biosystems 7500 Real Time PCR System定量仪上进行,按照试剂盒上的说明加样,反应总体系为20μL,内含cDNA 2μL、引物各0.8μL(10μmol·L-1)、SYBR®Premix Ex Taq II 10μL、ROX Reference Dye or Dye II 0.4μL、dH2O 6μL,采用两步法进行扩增,反应程序为95℃30 s、95℃5 s、60℃34 s,共40个循环。每个样品3次重复实验以减少误差,先用标准品梯度稀释进行定量PCR获得内参基因及谷胱甘肽S-转移酶基因的标准曲线,荧光定量的实验数据采用双标准曲线法进行相对定量分析,计量数据资料均用平均值±标准误表示[17],使用SPSS 17.0软件进行显著性分析,用Turkey法进行多重比较分析(P<0.05为显著水平)。
1.2.5 团头鲂肝和鳃组织应答氨氮胁迫后GST基因的表达分析
在氨氮胁迫后0 h、3 h、6 h、12 h、24 h、48 h时,分别在处理组和对照组中随机取3尾团头鲂,取肝和鳃保存于-80℃冰箱用以提取RNA,再经反转录获得cDNA模板,进行荧光定量PCR分析。反应在Applied Biosystems 7500 Real Time PCR System中进行,对照组和处理组的团头鲂肝和鳃组织cDNA样品在每个时间点做3个荧光定量特异引物和内参基因β-actin基因重复。
1.3 MaGST重组表达载体构建
根据所得到的MaGST的cDNA编码区设计特异性引物“5′AAGATGGCAATGAAATTGGCATA 3′,含有NdeⅠ酶切位点”和“(5′AAATGGGGAAACAAGGAGTGAA 3′,含有XhoⅠ酶切位点和6×His纯化标签”进行GST体外重组表达。PCR产物经1.0%琼脂糖凝胶电泳纯化后连入pMD18-T载体,转化Top10F’感受态细胞,通过特异性PCR进行阳性克隆筛选。培养经测序验证后的阳性克隆进行质粒提取并NdeⅠ和XhoⅠ双酶切,纯化后连接到pET21(a+)表达载体,转化大肠杆菌origami(DE3)感受态之后,菌液PCR验证并测序。
1.4 MaGST重组蛋白纯化、复性及活性检测
在200 mL LB培养基(含100μg·mL-1氨苄青霉素)中接入5 mL含有重组质粒的宿主菌origami (DE3),200 r·min-1,37℃培养至OD600达到0.5~0.7时,加入IPTG至终浓度为1 mmol·L-1,相同条件下继续培养,分别在诱导2 h和4 h后各取2 mL菌液,SDS-PAGE电泳检测。菌体加入20 mL缓冲液(50 mmol·L-1pH 7.4的PBS,含8 mol·L-1尿素),混匀,冰浴条件下超声破碎至液体变澄清,然后4℃下10 000 g离心30 min,收集上清。上清经过镍琼脂糖凝胶柱亲和纯化。纯化后的重组蛋白,依次在含有6、4、2、1和0 mol·L-1尿素的50 mmol·L-1PBS(pH 7.4)中过夜透析(4℃),之后用不含尿素的50 mmol·L-1PBS(pH 7.4)连续透析2次。利用15%SDS-聚丙烯酰胺凝胶电泳(SDS-PAGE)法检测透析后所获得的纯化样品,采用牛血清白蛋白(BSA)作为标准制作标准曲线,蛋白浓度的测定参考Bradford(1976)的方法进行[18]。在进行测定时,以透析结束时的透析缓冲液代替待测蛋白作为对照。重组MaGST活性的测定采用南京建成的谷胱甘肽-S转移酶测定试剂盒,单位为U·mg-1蛋白。

图1 团头鲂GST基因cDNA全序列以及由此推测的氨基酸序列注:方框表示起始密码子和终止密码子,星号表示翻译终止,大写字母表示开放阅读框,阴影部分表示Mu loop结构域,方框表示GST基因N-terminal和C-terminal结构域,下划线表示PolyA加尾信号。Fig.1 cDNA sequence of GST gene and putative amino acid sequence inM.amblycephalaNote:The box respresents the initiation codon and stop codon,“*”means translation termination;the capital letter means ORF;the Mu loop domain was shaded in gray and N-terminal and C-terminal domain were boxed;the PolyA signal was marked by single underline.
1.5 鳃和肝脏组织光镜样品制备与观察
氨氮胁迫实验期间,分别于0 h和24 h采集样品。每次从每个循环桶中随机取出3尾进行活体解剖,取各实验组的肝和鳃经生理盐水漂洗后,用Bouin氏液固定,用常规梯度酒精逐级脱水,二甲苯透明,石蜡包埋,切片机连续切片,厚度为5μm,苏木精和伊红染色,脱水封片,显微观察并拍照。
2 结果(Results)
表2 团头鲂GST基因氨基酸序列与其他脊椎动物的相似性比较Table 2 The similarity between the GST amino acid sequence ofM.amblycephalaand other vertebrates
2.1 团头鲂GST基因3’RACE和5’RACE扩增、克隆及全长拼接
如图1所示,利用Contig Express软件对中间序列、5’端序列、3’端序列进行拼接,得到团头鲂GST基因全长1 011 bp的cDNA序列,其中有133 bp的5’末端非翻译区(untranslated regions,UTR),224 bp的3’UTR,并且有明显的AATAAA加尾信号和Poly(A)尾,654 bp的开放阅读框(open reading frame, ORF),编码218个氨基酸。团头鲂GST主要有Mu loop(GEAPDYD),GST N-terminal和GST C-terminal三个结构域。通过Compute pI/Mw程序预测该基因的蛋白分子量为25.95 kD,等电点(pI)为5.44。团头鲂GST基因cDNA序列已提交到GenBank,登陆号为KM586249。

小鼠Mus musculus Mm Alpha1 AAH61134 Mm Alpha2 AAH30173 Mm Alpha3 AAH09805 Mm Alpha4 AAH12639 Mm Kappa1 NP-083831 Mm Mu4 AAH30444 Mm Mu6 AAH31818 Mm Pi AAH61109 Mm Theta1 AAH12254 Mm Theta2 Q61133 Mm Theta3 AAH03903 Mm mGST1 AAQ93322 Mm mGST2 NP-778160 Mm mGST3 AAH29669斑马鱼Danio rerio Dr Alpha AAH60914 Dr Kappa1 XP-686115 Dr Mu NP-997841 Dr Mu1 XP-690427 Dr Pi AAH83467 Dr Pi2 NP-001018349 Dr Theta NP-956878 Dr Theta3 XP-692427 Dr Rho CAK10882 Dr mGST1 AAH74022 Dr mGST2 XP-700815 Dr mGST3 XP-695658河豚Tetraodon nigroviridis PlRho CAA64493 PlRho CAA64496大口黑鲈Micropterus salmoides Ms Rho AAQ91198 Tn Alpha CAG09409 Tn Mu CAG07510 Tn Theta CAG09655 Tn mGST1 CAF97117 Tn mGST2 CAG04538 Tn mGST3 CAG09920鲽鱼Pleuronectes platessa真鲷Pagrus major Pm Rho BAD98443鲦鱼Pimephales promelas Pr Rho AAF78081鲫鱼Carassius auratus Ca Pi ABF57553欧鳗Anguilla anguilla Aa Pi AAS01601团头鲂Megalobrama amblycephala Ma Mu KM586249
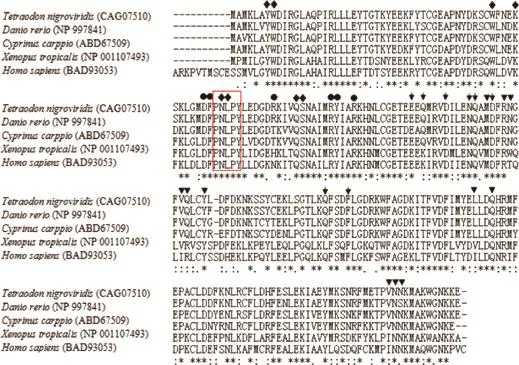
图2 团头鲂mu型GST基因推导氨基酸序列的多序列比较注:Mu型GST基因结构域用红色方框框出;GSH结合位点用菱形标出;底物结合位点用倒三角标出;N端二聚体用圆形标出,C端二聚体用箭头标出。Fig.2 Multiple sequence alignments of mu-MaGST with other five homologous mu-GST amino acid sequencesNote:The mu class GST motif is boxed in red.Diamond,GSH binding sites;Inverted triangle,substrate binding sites;Circular,dimer interface in N-terminal domain;and arrow,dimer interface in C-terminal domain.

图3 利用MEGA4.0软件构建各物种GST基因进化树Fig.3 Phylogenetic tree of GST amino acid sequence in diffrerent groups by MEGA 4.0
2.2 团头鲂GST基因的同源性分析
氨基酸序列比对结果显示团头鲂GST的氨基酸序列与草鱼GST同源性最高,相似度高达91%,运用DNAMAN软件将团头鲂GST的氨基酸序列与其他已公布的6种鱼类的氨基酸序列进行比对分析,结果如图2所示,显示该基因在鱼类中的同源性较高,均有比较保守的3个酶催化位点以及底物结合位点。
2.3 团头鲂GST基因的系统进化树分析
团头鲂与其他脊椎动物的GST基因氨基酸序列的亚型与基因登录号如表2所示,其他脊椎动物包括人、小鼠、斑马鱼、河豚、大口黑鲈、真鲷、鲦鱼、鲫鱼等。
用MAGE4.0软件对已经公布的序列以及团头鲂的GST基因氨基酸序列采用邻位相联法构建系统进化树。如图3所示,团头鲂GST基因与其他脊椎动物mu型GST基因亲缘关系较近并处在同一分支,表明团头鲂GST基因属于mu型。鱼类与哺乳动物GST亲缘关系较远,可能与鱼类GST基因在水环境毒素去毒代谢中承担的特殊功能有关。
2.4 团头鲂GST基因的组织表达
以团头鲂 β-actin为内参基因采用荧光定量PCR检测健康团头鲂肌肉、鳃、肾、脑、肝、肠的这6个组织中GST基因的相对表达量,结果如图4所示:该基因在肌肉、鳃、肾、脑、肝、肠道内均有表达,其中在肝和鳃组织中表达量较高,在肠道组织中表达量最低。

图4 荧光定量PCR方法检测团头鲂各组织中GST mRNA的相对表达量Fig.4 GST mRNA expression in different tissues of M.amblycephalaby qRT-PCR
2.5 团头鲂GST的mRNA在受到氨氮胁迫后不同时间点表达规律
采用荧光定量PCR方法检测团头鲂受到氨氮胁迫6、12、24、48 h后肝和鳃中GST的表达规律。肝组织和鳃组织的胁迫前后表达模式类似,如图5所示,团头鲂肝组织中的GST基因在氨氮胁迫的6~48h时,其表达量显著高于对照组(P<0.05);由图6中可知,与对照组相比,团头鲂鳃组织中的GST基因表达水平在胁迫6~48 h显著上升(P<0.05)。
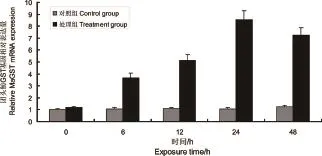
图5 氨氮胁迫后团头鲂肝组织中GST基因mRNA的表达量变化注:*表示与对照组差异显著(n=3,*P<0.05)。Fig.5 Expression of GST in liver ofM.amblycephalaunder acute ammonia exposueNote:* means significant different compared with control(n=3,*P<0.05).

图6 氨氮胁迫后团头鲂鳃组织中GST基因mRNA的表达量变化注:*表示与对照组差异显著(n=3,*P<0.05)。Fig.6 Expression of GST in gill ofM.amblycephalaunder acute ammonia exposueNote:* means significant different compared with control(n=3,*P<0.05).

图7 重组团头鲂GST蛋白SDS-PAGE电泳结果注:M,标准分子质量Marker;1,未加IPTG的大肠杆菌培养物(对照);2,添加IPTG诱导2 h;3,添加IPTG诱导4 h;4,纯化后的团头鲂GST重组蛋白。Fig.7 The SDS-PAGE result of recombinant MaGSTNote:Line M shows the protein marker;Lane 1,negative control for recombinant MaGST(without induction);lane 2,induced expression for 2 h of recombinant MaGST;lane 3,induced expression for 4 h of recombinant MaGST;lane 4,purified recombinant MaGST.
2.6 团头鲂GST原核重组表达及活性测定
将团头鲂GST的编码区重组到pET21(a+)中,通过IPTG诱导表达,利用凝胶柱纯化得到带有6× His标签的团头鲂GST蛋白。从图7所示,所获得的纯化蛋白的分子量为26 kDa左右,与预测的分子量结果相吻合。经GST检测试剂盒测定,重组团头鲂GST的GST活力为(10.36±0.68)U·mg-1蛋白。
2.7 氨氮胁迫导致鳃和肝脏组织病理变化
氨氮胁迫前,团头鲂幼鱼的鳃丝和鳃小片泌氯细胞较少,鳃丝排列整齐规则(图8),而氨氮胁迫24 h时,泌氯细胞较少、鳃小片融合、变短,基部毛细血管破裂。氨氮胁迫前,团头鲂幼鱼的肝细胞排列整齐,细胞轮廓清晰(图8),而氨氮胁迫24 h时,肝细胞水样变性严重,细胞溶解,轮廓模糊,血窦扩张严重。

图8 氨氮胁迫对团头鲂幼鱼鳃和肝脏组织显微结构的影响注:A表示氨氮胁迫0 h(对照组)鳃丝组织显微结构图;B分别表示氨氮浓度(25mg·L-1)胁迫24 h鳃丝组织显微结构图;C表示表示氨氮胁迫0 h(对照组)肝组织显微结构图;D表示氨氮浓度(25mg·L-1)胁迫24 h肝组织显微结构图。H.E染色(×200),标尺50μm。CC,泌氯细胞;SL,鳃小片;LF,鳃小片融合;SSL,鳃小片变短;H,肝细胞;NH,细胞核肿大;DS,血窦扩张。Fig.8 Effect of ammonia exposure on gill and liver microstructure in juvenileM.amblycephalaNote:A means the microscopical gill structure of juvenileM.amblycephalaexposed to ammonia at 0 h;B means the microscopical gill structure of juvenileM.amblycephalaexposed to ammonia(25mg·L-1)at 24 h;C means the microscopical liver structure of juvenileM.amblycephalaexposed to ammonia at 0 h;D means the microscopical liver structure of juvenileM.amblycephalaexposed to ammonia(25mg·L-1)at 24 h.H.E(×200), bar=50μm.CC,chloride cells;SL,secondary lamellae;LF,lamellar fusion;SSL,shortening of secondary lamellae;H,hepatocytes;NH,nuclear hypertrophy;DS,dilatation in sinusoids.
3 讨论(Discussion)
GST基因是鱼类重要的Ⅱ相去毒酶基因,其表达产物一方面缓解氧化压力、抑制活性氧对机体的损伤,另一方面催化毒素形成亲水化合物经排泄系统排出体外[19]。GST基因亚型的分类基于对该基因N端结构域的氨基酸、结构、抗体反应以及对抑制剂的敏感性[7]。本研究通过RACE PCR技术获得了团头鲂GST基因的cDNA序列,包含1个典型的GST-N结构域和1个GST-C结构域,其编码的氨基酸序列与其他近源物种比对结果可判断扩增得到的cDNA序列属于GST基因家族,同时与其他物种的Mu-型GST相比,具有一致的Mu-loop结构域[20]。此外,系统进化树分析结果也表明团头鲂GST属于mu型的1个成员,团头鲂 GST与斑马鱼 mu型GST在系统树上处于相邻位置,且与其他鱼类(河豚)聚成1簇,与团头鲂分类学上的地位吻合。团头鲂GST的组织定量表达分析显示,该基因在各组织中均有表达,但在肝组织中的表达量最高,然后依次是鳃、肾、肌肉、脑和肠道,这与已报到的鲤鱼(Cyprinus carpio)GST基因的组织表达结果相一致[21]。
鳃和肝脏组织是鱼类重要的呼吸和解毒器官,鉴于GST基因在团头鲂肝脏和鳃组织表达量较高,本实验选择肝和鳃组织作为研究对象,在氨氮急性胁迫后不同时间段对团头鲂肝和鳃组织中GST基因的定量表达分析显示,肝和鳃组织中该基因的表达规律相似,表达水平均呈先升高后降低的趋势,研究表明团头鲂幼鱼鱼体处于在高浓度氨氮应激初始状态下防御与损伤或许处于平衡状态,随着急性胁迫时间延长,平衡系统被打破组织发生损伤(肝和鳃组织结构变化),机体抗氧化能力下降,这是因为GST的过氧化物酶活性能利用谷胱甘肽向氢过氧化物发动亲核攻击,使其还原为低毒的一元醇(monohydroxy alcohol),从而缓解氧化胁迫[22],而随着氨氮胁迫时间的增加,过量的自由基和过氧化产物或许抑制了GST基因的转录与翻译。最近在哺乳动物的研究中已经证实氨中毒引发脑组织谷氨酰胺的积累进一步导致其中枢神经系统(CNS)的氧化应激并产生大量活性氧分子,这些活性氧分子不但直接对细胞造成损伤,还将通过引发脂质过氧化作用介导细胞膜的功能紊乱[23],而Hegazi等[24]在氨氮胁迫下的尼罗罗非鱼肝脏和肌肉中也发现大量的活性氧自由基,抗氧化酶活性显著升高,Sun等[25]也发现慢性氨氮胁迫下的鳙鱼也存在类似的氧化应激反应,这与本实验室结果相吻合。为了进一步确定氨氮胁迫是否产生一定程度氧化应激损伤,本实验选取GST表达量较高的时间点进行鳃和肝脏组织结构观察,氨氮胁迫24 h下团头鲂幼鱼鳃丝毛细血管扩张,泌氯细胞增生,鳃小片卷曲,鳃小片融合、变短;而氨氮胁迫24 h下团头鲂幼鱼肝细胞肿大、细胞核变形溶解、血窦扩张、细胞轮廓模糊,这与张武肖等[16]研究结果相一致。过量的自由基介导脂质过氧化产物产生,丙二醛(MDA)被认为是脂类过氧化损伤最具代表性的指标[26],胡毅等[27]的研究表明青鱼MDA含量随着氨氮胁迫时间的延长而显著增加,蒋玫等[28]的研究表明慢性氨氮胁迫对鲻鱼MDA活性与氨氮浓度呈一定的正相关。故此,我们可推断活性氧自由基与内源性抗氧化能力的不平衡是导致氨氮毒性分子机制的主要原因之一。
鉴于mu型GST在细胞解毒和抗氧化防御中起着非常重要的作用[29]。本研究利用原核表达方法获得了重组MaGST蛋白,其GST活力为(10.36± 0.68)U·mg-1蛋白,这与在真鲷(Pagrus major)GST重组蛋白活性相类似[30]。GST活力能反映机体抗氧化能力的高低,Lee等[31]通过原核表达的方法获得了重组GST蛋白,其活性为天然GST的45%,故此本研究测定的重组MaGST蛋白的活力也可能只是其活性的部分。本研究纯化的重组蛋白可用于制备MaGST的单克隆抗体,并用于GST相关表达的检测,为团头鲂mu-型GST功能的进一步研究奠定了坚实基础,如何增强淡水鱼类解毒酶基因的表达以减轻急性氨氮胁迫导致的致毒作用有待于进一步研究。
[1]江晓浚,孙盛明,戈贤平,等.添加不同碳源对零换水养殖系统中团头鲂鱼种生长、肠道生化指标和水质的影响[J].水产学报,2014,38(8):1113-1122 Jiang X J,Sun S M,Ge X P,et al.The effect of different carbon sources on growth,intestinal biochemical parameters and water quality in the juvenileMegalobrama amblycephalacultured in zero water exchanged system[J]. Journal of Fisheries of China,2014,38(8):1113-1122(in Chinese)
[2]El-Shafai S A,El-Gohary F A,Nasr F A,et al.Chronic ammonia toxicity to duckweed-fed tilapia(Oreochromis niloticus)[J].Aquaculture,2004,232(1–4):117-127
[3]Hegazi M M,Hasanein S S.Effects of chronic exposure to ammonia concentrations on brain monoamines and ATPases of Nile tilapia(Oreochromis niloticus)[J].Comparative Biochemistry and Physiology C,2010,151(4):420-425
[4]Benli A C K,Köksal G,Özkul A.Sublethal ammonia exposure of Nile tilapia(Oreochromis niloticusL.):Effectson gill,liver and kidney histology[J].Chemosphere, 2008,72(9):1355-1358
[5]Lushchak V I,Bagnyukova T V.Effects of different environmental oxygen levels on free radical processes in fish [J].Comparative Biochemistry and Physiology B,2006, 144(3):283-289
[6]Sheehan D,Meade G,Foley V M,et al.Structure,function and evolution of glutathione transferases:Implications for classification of non-mammalian members of an ancient enzyme superfamily[J].Biochemistry Journal, 2001,360(1):1-16
[7]Kim M,Ahn I Y,Cheonand J,et al.Molecular cloning and thermal stress induced expression of a pi-class glutathione S-transferase(GST)in the Antarctic bivalveLaternula elliptica[J].Comparative Biochemistry Physiology A,2009,152(2):207-213
[8]Arca P,Hardisson C,Suarez J.Purification of a glutathione S transferase that mediates fosfomycin resistance in bacteria,Antimicrob[J].Agents Chemother,1997,34(5): 844-848
[9]Hatton P J,Cummins I,Cole D J,et al.Glutathione transferase involved in herbicide detoxification in the leaves of Setaria faberi(giant toxtail)[J].Physiology Plant,1999, 105(1):9-16
[10]Ranson H,Rossiter L,Ortelli F,et al.Identification of a novel class of insect glutathione S-transferase involved in resistance to DDT in the malaria vectorAnopheles gambiae[J].Biochemistry Journal,2001,359(2):295-304
[11]Eaton D L,Bammler T K.Concise review of the glutathione S-transferases and their significance to toxicology[J]. Toxicological Sciences,1999,49(2):156-164
[12]Hayes J D,Flanagan J U,Jowsey I R.Glutathione transferases[J].Annual Review of Pharmacology and Toxicology,2005,45:51-88
[13]程炜轩,梁旭方,李观贵,等.鳜鱼两种谷胱甘肽S-转移酶基因 cDNA的克隆与分析[J].生态毒理学报, 2009,4(4):537-543 Cheng W X,Liang X F,Li G G.Molecular Cloning and sequence analysis of alpha-and rho-classes of glutathione S-transferases in Chinese perch(Siniperca chuatsi)[J].A-sian Journal of Ecotoxicology,2009,4(4):537-543(in Chinese)
[14]Angelucci S,Sacchetta P,Moio P,et al.Purification and characterization of glutathione transferase from the sea bass(Dicentrarchus labrax)liver[J].Archives of Biochemistry and Biophysics,2000,373(2):435-441
[15]Melgar-Riol M J,Nóvoa-Valiñas M C,García-Fernández M A,et al.Glutathione S-transferase from rainbow trout liver and freshly isolated hepatocytes:Purification and characterization[J].Comparative Biochemistry and Physiology C,2001,128(2):227-235
[16]张武肖,孙盛明,戈贤平,等.急性氨氮胁迫及毒后恢复对团头鲂幼鱼鳃、肝和肾组织结构的影响[J].水产学报,2015,39(2):233-244 Zhang W X,Sun S M,Ge X P,et al.Acute effects of ammonia exposure on histopathology of gill,liver and kidney in juvenileMegalobrama amblycephalaand the postexposure recovery[J].Journal of Fisheries of China, 2015,39(2):233-244(in Chinese)
[17]Livak K J,Schmittgen T D.Analysis of relative gene expression data using realtime quantitative PCR and the 2(-Delta Delta C(T))method[J].Methods,2001,25(4): 402-408
[18]Bradford M M.A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding[J].Analytical Biochemistry,1976,72:248-254
[19]Gehringer M M,Shcphard E G,Downing T G,et al.An investigation into the detoxification of microcystin-LR by the glutathione pathway in Balb/c mice[J].International Journal of Biochemistry& Cell Biology,2004,36(5): 931-941
[20]Dirr H,Reinemer P,Huber R.X-ray crystal structures of cytosolic glutathione S-transferase.Implications for protein architecture,substrate recognition and catalytic function[J].European Journal Biochemistry,1994,220(3): 645–661
[21]Fu J,Xie P.The acute effects of microcystin LR on the transcription of nine glutathione S-transferase genes in common carp,Cyprinus carpioL[J].Aquatic Toxicology, 2006,80(3):261-266
[22]Edwerds R,Dixon D P,Walbot V.Plant glutathione S-transferases enzymes with multiple functions in sickness and in health[J].Trends in Plant Science,2000,5(5): 193-198
[23]Kosenko E,Venediktova N,Kaminsky Y.Sources of oxygen radicals in brain in acute ammonia intoxication in vivo[J].Brain Research,2003,981(1–2):193–200
[24]Hegazi M M,Attia Z I,Ashour O A.Oxidative stress and antioxidant enzymes in liver and white muscle of Nile tilapia juveniles in chronic ammonia exposure[J].Aquatic Toxicology,2010,99(2):118-125
[25]Sun H,Wang W,Li J,et al.Growth,oxidative stress responses,and gene transcription of juvenile bighead carp (Hypophthalmichthys nobilis)under chronic-term exposure of ammonia[J].Environmental Toxicology Chemis-try,2014,33(8):1726-1731
[26]李磊,蒋玫,沈新强,等.Cr(VI)对脊尾白虾(Exopalaemon carinicauda)幼虾暴露和恢复期肝胰脏的SOD活性、MDA及MTs含量的影响[J].生态毒理学报,2014, 9(6):1226-1231 Li L,Jiang M,Shen X Q,et al.Effects of Cr(VI)exposure and recovery on the SOD activities,contents of MDA and MTs in hepatopancreas tissue of juvenileExopalaemon carinicauda[J].Asian Journal of Ecotoxicology,2014,9 (6):1226-1231(in Chinese)
[27]胡毅,黄云,钟蕾,等.氨氮胁迫对青鱼幼鱼鳃丝Na+/ K+-ATP酶、组织结构及血清部分生理生化指标的影响[J].水产学报,2012,36(4):538-545 Hu Y,Huang Y,Zhong L,et al.Effects of ammonia stress on the gill Na+/K+-ATPase,microstructure and some serum physiological-biochemical indices of juvenile black carp(Mylopharyngodon piceus)[J].Journal of Fisheries of China,2012,36(4):538-545(in Chinese)
[28]蒋玫,李磊,沈新强,等.慢性氨氮胁迫对鲻鱼(Mugil cephalus)幼鱼组织细胞免疫指标的影响研究[J].海洋与湖沼,2014,45(3):529-535 Jiang M,Li L,Shen X Q,et al.Effect of ammonia stress on immunity indicators of juvenileMugil cephaus[J]. Chinese Journal of Oceanology and Limnology,2014,45 (3):529-535(in Chinese)
[29]瞿春梅,梁旭方,张进,等.日本沼虾mu型可溶性谷胱甘肽S-转移酶的克隆与表达[J].华中农业大学学报, 2013,32(2):103-108 Qu C M,Liang X F,Zhang J,et al.Molecular cloning and in vivo expression analysis of mu-class soluble glutathione S-tranferase in shrimp[J].Journal of Huazhong Agricultural University,2013,32(2):103-108(in Chinese)
[30]Konishi T,Kato K,Araki T,et al.Molecular cloning and characterization of alpha-class glutathione S-transferase genes from the hepatopancreas of red sea bream,Pagrus major[J].Comparative Biochemistry Physiology C,2005, 140(3-4):309-320
[31]Lee Y M,Lee K W,Park H,et al.Sequence,biochemical characteristics and expression of a novel Sigma-class of glutathione S-transferase from the intertidal copepod,Tigriopus japonicuswith a possible role in antioxidant defense[J].Chemosphere,2007,69(6):893-902
Molecular Cloning,Characterization and mRNA Expression of Mu-typ Glutathione S-Transferases from Megalobrama amblycephala
Sun Shengming1,Zhu Jian1,*,Ge Xianping1,Zhang Chengfeng1,Miao Linghong1,Zhang Wuxiao2, Zhang Qiong2
1.Freshwater Fisheries Research Center,Chinese Academy of Fishery Sciences,Wuxi 214081,China
2.Wuxi Fishery College,Nanjing Agricultural University,Wuxi 214081,China
9 March 2015 accepted 9 June 2015
Glutathione S-transferases(GSTs)play an important role in cellular detoxification and may have evolved in protecting cells against reactive oxygen metabolites.In this study,we report the molecular characterization of glutathione S-transferase(MaGST)from Bluntnose black breamMegalobrama amblycephala.The full-length cDNA of MaGST was 1 011 bp in length containing an open reading frame of 654 bp that encoded a 218-amino acid pu-tative protein.Its derived amino acid sequence was clustered with other vertebrate mu-type GSTs in a phylogenetic tree(NJ).Quantitative real-time RT-PCR analysis showed that mRNA expression of MaGST was detected in all tissues,including the brain,liver,muscles,gills,spleen,and intestines,with the highest level of expression in the liver and gill,the lowest expression in the muscles.Exposure to waterbrone ammonia significantly increased the mRNA expression of MaGST(P<0.05)in liver and gill,respectively.Fish exposed to waterbrone ammonia also showed liver and gill tissue damage in 24 h.To further characterize the catalytic properties of this enzyme along with MaGST, we constructed the recombinant MaGST plasmid with a 6×His-Tag at the N-terminal of MaGST cDNA.Recombinant MaGST was highly expressed in transformedEscherichia coli,and its soluble fraction was purified by His-Tag affinity column chromatography.The GST activity of the recombinant MaGST was(10.36±0.68)U·mg-1protein.Overall,the results suggest a potential role for MaGST in detoxification and possibly conferring immune protection.
ammonia exposure;Megalobrama amblycephala;glutathione S-transferase;cloning
2015-03-09 录用日期:2015-06-09
1673-5897(2016)1-295-11
X171.5
A
10.7524/AJE.1673-5897.20150309010
孙盛明,朱健,戈贤平,等.团头鲂谷胱甘肽S-转移酶基因的克隆及其在氨氮胁迫中的表达分析[J].生态毒理学报,2016,11(1):295-305
Sun S M,Zhu J,Ge X P,et al.Molecular cloning,characterization and mRNA expression of mu-typ glutathione S-transferases fromMegalobrama amblycephala[J].Asian Journal of Ecotoxicology,2016,11(1):295-305(in Chinese)
中央级公益性科研院所基本科研业务费专项资金(2015C06XK01);国家大宗淡水鱼类产业技术体系华东养殖岗位(CARS-46-14);十二五国家科技支撑计划“长江下游池塘高效生态养殖技术集成与示范”(2012BAD25B07)
孙盛明(1983-),男,博士,助理研究员,研究方向为水生动物生理学,E-mail:sunsm@ffrc.cn;
),E-mail:zhuj@ffrc.cn
简介:朱健(1968—),男,研究员,主要从事水产健康养殖研究。
