高原鼠兔干扰对高寒草甸植物多样性与土壤养分间关系的影响
2016-11-28庞晓攀贾婷婷郭正刚
王 莹,庞晓攀,肖 玉,贾婷婷,王 倩,于 成,郭正刚
兰州大学草地农业科技学院, 草地农业生态系统国家重点实验室,兰州 730020
高原鼠兔干扰对高寒草甸植物多样性与土壤养分间关系的影响
王 莹,庞晓攀,肖 玉,贾婷婷,王 倩,于 成,郭正刚*
兰州大学草地农业科技学院, 草地农业生态系统国家重点实验室,兰州 730020
高原鼠兔干扰虽然能够改变高寒草甸植物多样性与土壤养分含量,但植物多样性与土壤养分间的关系对高原鼠兔干扰的响应尚不清晰。利用高原鼠兔有效洞口密度将高原鼠兔干扰程度划分为T1(7个/625 m2)、T2(12个/625 m2)、T3(22个/625 m2)、T4(38个/625 m2)4个水平,运用RDA冗余分析法研究了高原鼠兔不同干扰程度下高寒草甸植物多样性与土壤养分间的关系。结果表明:随着高原鼠兔干扰水平的增加,优势种高山嵩草(Kobresiapygmaea)的重要值先增加后降低,而伴生种小花草玉梅(Anemonerivularisvar.flore-minors)和莓叶委陵菜(Potentillafragarioides)的重要值先降低后增加;当高原鼠兔干扰水平从T1到T2时植物多样性指数变化不显著,而高原鼠兔干扰程度超过T2时则植物多样性指数具有降低趋势;土壤全氮和硝态氮含量随高原鼠兔干扰水平增加而降低,而土壤铵态氮含量则降低后增加,土壤有机碳和全磷先增加后降低;多样性指数与0—10cm土壤深度硝态氮、10—20cm土壤深度全钾间的相关性从T1到T3时为正相关,而到T4时则变为负相关,而与0—10cm土壤深度全氮的相关性则表现T1到T3时为负相关,T4时为正相关,与铵态氮间相关性只有T1时为负相关,这说明高原鼠兔干扰改变了植物多样性与土壤养分间的关系,其变化阈值介于T2和T3。
高原鼠兔;植物多样性;协同性;土壤养分
土壤是植物生长的基础和营养库[1-2],而不同植物正常生长所需的养分元素种类和数量具有一定的分异性[3-5]。若土壤养分种类及其含量发生变化时,草地植物群落组分可能会发生一定程度的改变[6],原有物种会被更适宜生长于此土壤条件下的物种所代替,植物多样性也随之变化[7],以适应土壤养分及其种类的供给。植物群落内组分的种类发生明显变化时,植物多样性则可能发生变化,植物群落消耗或增加土壤中某些养分元素的含量[8],引起土壤性质发生改变,进而影响土壤的稳定性[9]。因此,任何健康的草地生态系统,其土壤养分和植物多样性总是处于相互适应和协同变化的过程中,这种协同变化表现为植物与土壤间相互依存、彼此适应和协同[10],以维持生态系统的健康和动态稳定。高寒草甸植物多样性与土壤养分间的协调关系往往因外界环境的干扰而改变,当干扰程度较小时可以促进高寒草甸植物组分更新[11],土壤性质改良[12],增强植物与土壤间的协同性[13],利于高寒草甸生态系统的正向演变[13- 14],而当干扰程度过大时有利于高寒草甸植物群落组分中杂类草,毒草等增加[15],土壤性质劣化[16],减弱植物与土壤间的协同性,迫使高寒草甸生态系统的逆向演变。
高原鼠兔是青藏高原特有的小型哺乳类动物,也是高寒草甸生态系统的关键种[17],其往往通过掘洞、采食牧草以及排泄粪便等活动对高寒草甸产生直接或间接的干扰[18- 20]。目前,关于高原鼠兔干扰的研究仅限于土壤或植被,集中于植被地上数量特征的改变及地下土壤养分量化指标的变化。已有研究表明,当高原鼠兔干扰程度较小时,能够增加土壤水分和养分含量[21],增加植物群落中禾本科植物比例和植物物种多样性[22],提高可食牧草生物量的比例[23- 24],增强草甸生态系统的稳定性[25],而干扰程度过大时会降低土壤养分含量,促进生境旱化[21,26],杂类草和毒草等组分增加[27],弱化草甸生态系统的稳定性[28],这说明高寒草甸植物多样性和土壤养分对高原鼠兔干扰程度的响应具有明显的分异性,这种分异性可能主要来源于植物的生态位宽度随着高原鼠兔干扰强度增加而发生明显的变化,有些植物的生态位宽度增加,有些植物的生态位宽度减小[11],而植物生态位的变化过程受碳、氮、磷元素计量比的影响,不同物种由于利用氮、磷的比例存在分异而导致植物生态位的分化[13],这预示着高原鼠兔干扰高寒草甸时植物组成与土壤的关系可能发生了变化。然而,高原鼠兔干扰下高寒草甸植物多样性与土壤养分间关系究竟如何变化的,目前尚不清晰,因此研究高原鼠兔干扰对高寒草甸植物多样性与土壤养分间关系的影响,对确定提高植物多样性和土壤养分关系协同性的高原鼠兔干扰强度具有重要的意义。本研究通过分析高原鼠兔不同干扰程度下植物多样性和土壤养分的变化特征,查明不同高原鼠兔干扰水平下高寒草甸植物多样性与土壤养分间的相关性,为证实高原鼠兔干扰是否改变了高寒草甸植物多样性与土壤养分间相关性提供科学依据。
1 研究地区概况与方法
1.1 研究地区概况
研究地区位于青藏高原东部,行政区划隶属于甘肃省甘南自治州玛曲县,地理坐标为北纬 33°06′30″—34°30′15″,东经 100°40′45″—102°29′00″,海拔 3300—4806 m。气候属高原湿润气候,年降水量和年蒸发量分别大约为564mm和1000—1500mm,年均温度 1.2 ℃,最冷的1月份和最热的7月份平均温度分别为-10 ℃和11.7 ℃。全年日照时数 2613.9 h,年内霜期大于270d,年相对无霜期仅19d,无绝对无霜期。土壤为亚高山草甸土。植物类型主要是高寒草甸,优势种主要为高山嵩草(Kobresiapygmaea),主要伴生种钝裂银莲花(Anemoneobtusiloba),小花草玉梅(Anemonerivularisvar.flore-minors),鹅绒萎陵菜(Potentillaanserina),莓叶萎陵菜 (Potentillafragarioides)、蒲公英(Taraxacummongolicum)和长毛凤毛菊(Saussureahieracioides)等。1999年对甘南藏族自治州的统计资料显示,全州约30%的草地受到高原鼠兔危害的影响[29],其鼠害危害的草地面积具有逐年增大的趋势,其中高原鼠兔是主要危害啮齿类动物,主要生活于草丛低矮的环境,一旦入侵,整个生境几乎全部占据,只是密度有所不同,已经严重威胁高寒草甸生态系统的健康。
1.2 研究方法
1.2.1 试验设计和野外调查
本研究采用高原鼠兔有效鼠洞口密度划分高原鼠兔干扰强度[21,25,30]的方法,分析高原鼠兔干扰对高寒草甸植物多样性和土壤养分间的关系。2012年5月在甘肃境内玛曲县阿孜站附近的试验地随机选取了36个25 m×25 m(625 m2)的样地,采用连续3d堵洞法测定每个样地的有效洞口数,同时调查各样地的裸斑面积和草丛高度。根据有效洞口数,裸斑面积和草丛高度将36个样地聚类为4个类群,并以每个类群的平均值为基准确定高原鼠兔有效洞口梯度,其分别为10(10±3)、15(15±2)、21(21±3)、31(31±4)[11,13],每个有效洞口梯度选择3个样地作为重复,因此共建立了12个面积为25 m ×25 m(625 m2)的固定监测样地,划分为4个干扰程度处理。2014年8月,再次采用堵洞法调查了高原鼠兔有效洞口数密度,虽然每个样地有效洞口数稍有变化,但仍然遵循样地设置时的梯度,分别为7个/625 m2(7±2)、12个/ 625 m2(12±2)、22个/625 m2(22±5)、38个/625 m2(38±4),用T1、T2、T3、T4表示。每个样地内,采用“W”型布置5个 1 m×1 m的样方,样方间距离约8 m,样方均选择离鼠兔洞口约1m处的草甸上。统计样方内植株个体的种类、高度、频度、盖度等,其中高度测量采用样方内随机测定50次植物高度求其平均值法,盖度采用针刺点测法。调查完植被特征后采集土壤样品,按照0—10 cm和10—20 cm分层收集土样,每个样方内打钻3次且混合,装入密封袋带回实验室分析。
1.2.2 土壤测定方法
土壤有机碳(OC)采用重铬酸钾法测定,土壤全氮(TN)采用凯氏定氮法测定,土壤全磷(TP)采用钼锑抗比色法测定,土壤全钾(TK)和速效钾(AK)采用火焰分光光度计测定,土壤铵态氮(AN)和硝态氮(NN)采用连续流动分析仪测定,土壤速效磷(AP)采用双酸浸提钼锑抗比色法测定,测定方法参考中国土壤学会所编《土壤农业化学分析方法》[31]。
1.3 数据处理与分析
1.3.1 重要值
计算公式:
重要值=(相对盖度+相对频度+相对高度) /3
1.3.2 植物多样性
采用Margalef指数、Pielou指数和Shannon-Weiner指数分别测度丰富度指数,均匀度指数和多样性指数,计算公式如下:
Margalef指数=(S-1)/lnN
Pielou指数=-∑PilnPi/lnS
Shannon-Weiner指数=-∑PilnPi
式中,S为1 m2样方中的物种数,N为样方中物种总个体数,Pi为种的相对重要值[32]。
1.3.3 软件统计分析
利用Excel软件进行基础数据处理,采用SPSS17.0进行方差分析,采用Pearson相关性分析法分析了植物多样性与土壤养分间的相关性,采用CANOCO4.5软件对植物多样性与土壤因子间关系进行RDA冗余分析并作图。
2 结果与分析
2.1 高原鼠兔干扰对植物群落物种组成及其重要值的影响
不同高原鼠兔干扰程度下高寒草甸植物组分具有一定的差异,样地内物种数表现为先增加后降低趋势,干扰程度T2时物种数最多,而干扰程度T4时物种数最小(表1)。高原鼠兔干扰程度从T1增加至T4的过程中,优势种高山嵩草(Kobresiapygmaea)的重要值虽然呈现先增加后降低的变化过程,但其仍然是重要值最大的物种,说明高山嵩草占据主导地位,但这种主导地位却随着高原鼠兔干扰水平增加表现先增加后降低的趋势;伴生种小花草玉梅(Anemonerivularisvar.flore-minors)和莓叶委陵菜(Potentillafragarioides)重要值先降低后增加,其在植物群落内的作用表现为先弱化后增强;鹅绒委陵菜(Potentillaanserina)和秦艽(Gentianamacrophylla)的重要值随着干扰程度的加强而增加;圆叶筋骨草(Ajugaovalifolia)、大黄(Rheumpalmatum)和珠芽蓼(Polygonumviviparum)则随着高原鼠兔干扰程度增加而逐渐退出植物群落;少见种车前(Plantagoasiatica)、大蓟(Cirsiumjaponicum)、蒲公英(Taraxacummongolicum)和黄帚橐吾(Ligulariavirgaurea)等植物仅出现在某个干扰程度生境内。说明随高原鼠兔干扰强度增加,高寒草甸植物群落物种组成及其优势地位发生了不同程度的改变。

表1 不同高原鼠兔干扰程度下高寒草甸植物群落的物种组成及其重要值
―表示此物种在该样方中未出现
2.2 高原鼠兔干扰对植物多样性的影响
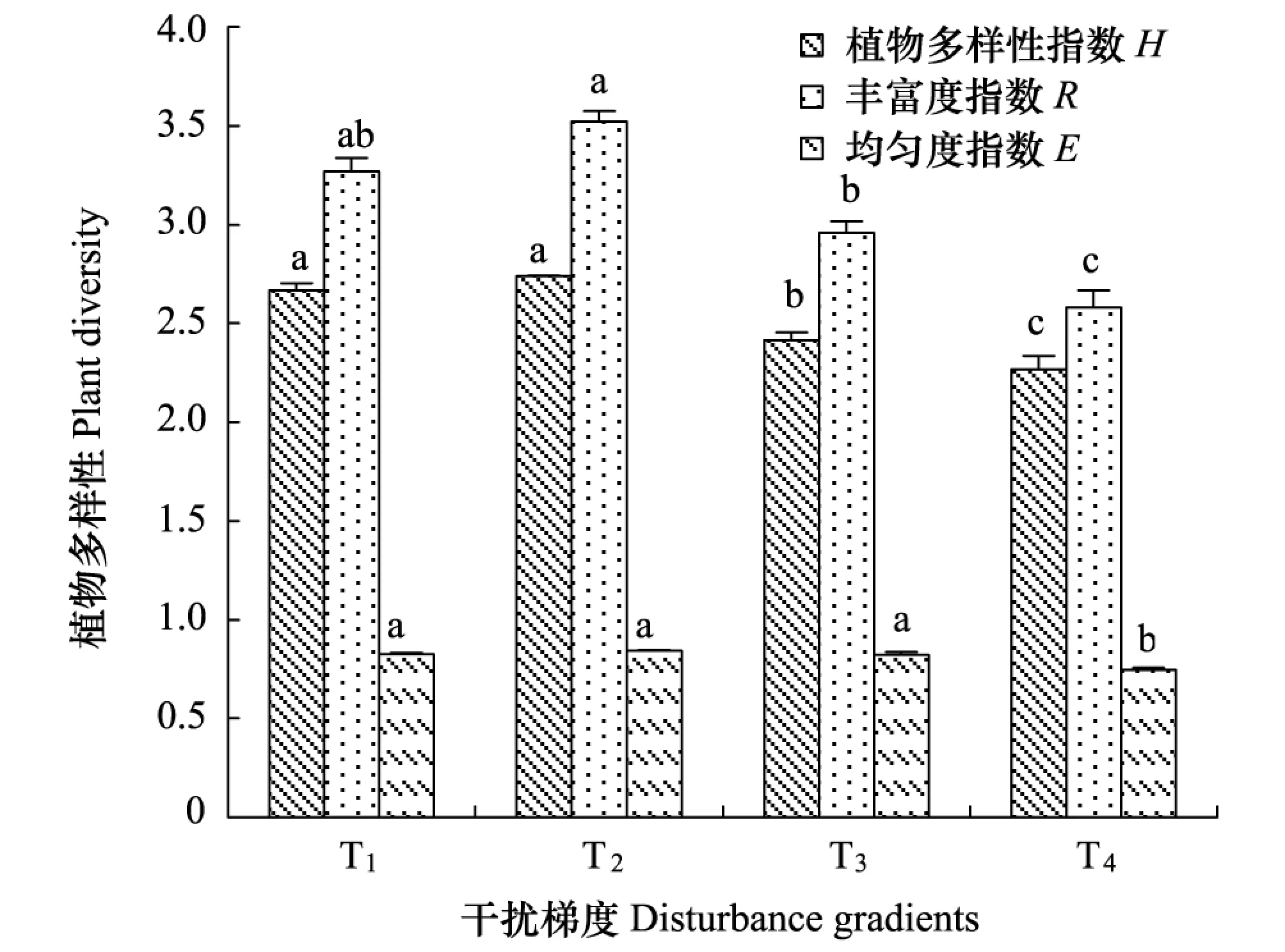
图1 不同高原鼠兔干扰梯度下高寒草甸的植物多样性 Fig.1 Plant diversity of alpine meadow at different disturbance gradients of plateau pika不同小写字母表示差异显著(P<0.05)
随着高原鼠兔干扰程度增加,丰富度指数、均匀度指数和多样性指数的变化趋势一致,均表现为降低的态势(图1)。但不同指数显著降低时对应的干扰程度水平存在分异,T2生境内的丰富度指数显著大于T3和T4生境内的丰富度指数(P<0.05),但与T1生境内的丰富度指数差异不显著;T4生境内的均匀度指数显著小于T1、T2和T3生境内的均匀度指数(P<0.05),但T1、T2和T3生境内的均匀度指数差异不显著;T3生境内的多样性指数显著小于T1和T2生境内的多样性指数(P<0.05),却显著大于T4生境内的多样性指数(P<0.05)。说明丰富度指数和均匀度指数发生明显变化的干扰梯度分别是T2和T4、T3和T4之间,而多样性指数发生明显变化的干扰梯度则为T2和T3之间。
2.3 高原鼠兔干扰对土壤养分的影响
高原鼠兔干扰和土壤深度对土壤有机碳,全磷,全氮,铵态氮和硝态氮均有显著影响(P<0.05)(图2),高原鼠兔干扰对土壤全钾、速效钾和速效磷均没有显著影响(P>0.05)(图2),但土壤深度却显著影响了三者的含量(P<0.05)。随高原鼠兔干扰程度增加,0—10cm土壤深度和10—20cm土壤深度土壤有机碳和全磷均表现为先增加后降低的趋势,T2干扰梯度时土壤有机碳含量最高,T4干扰梯度时土壤全磷含量最低;土壤全氮和硝态氮具有降低趋势,铵态氮却表现为先降低后增加的趋势。各个干扰梯度土壤有机碳、全钾、速效钾、全磷、速效磷、全氮和铵态氮含量均表现为0—10cm土壤深度显著高于10—20cm土壤深度(P<0.05),而硝态氮含量却表现为0—10cm土壤深度显著低于10—20cm土壤深度(P<0.05)。高原鼠兔干扰和土壤深度互作对土壤有机碳、全磷、速效磷、速效钾、全氮、硝态氮、铵态氮均具有显著影响(P<0.05),而对土壤全钾没有明显影响,说明多数土壤养分含量不仅受土壤深度和高原鼠兔干扰的影响,还受土壤深度和高原鼠兔干扰互作的影响。
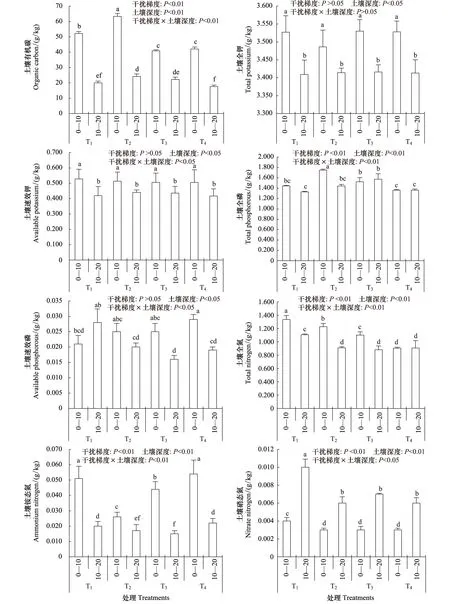
图2 高原鼠兔干扰梯度和土壤深度对土壤养分含量的影响Fig.2 Effect of disturbance gradients of plateau pika and soil deeps on soil nutrient contents 不同小写字母表示差异显著(P < 0.05)
2.4 高原鼠兔干扰下土壤养分与植物多样性关系
RDA分析结果表明,0—10cm土壤深度的4个排序轴特征值分别为0.868、0.018、0.011、0.079(表2),其中第一轴是约束性排序轴,对响应变量的解释比例达86.8%,后3个轴是非约束性的,4个排序轴的特征值之和占总特征值的89.7%。植物多样性与土壤养分间的相关性在前两个排序轴分别为0.958和0.648,且物种养分关系分别达到96.8%和98.7%。而10—20cm土壤深度的4个排序轴特征值分别为0.620、0.021、0.010、0.333,4个排序轴解释变量达98.4%,植物多样性和土壤养分的相关性在前两轴分别为0.812和0.676。上述结果说明,0—10cm土壤深度和10—20cm土壤深度均能够很好地表达植物多样性与土壤养分间关系。
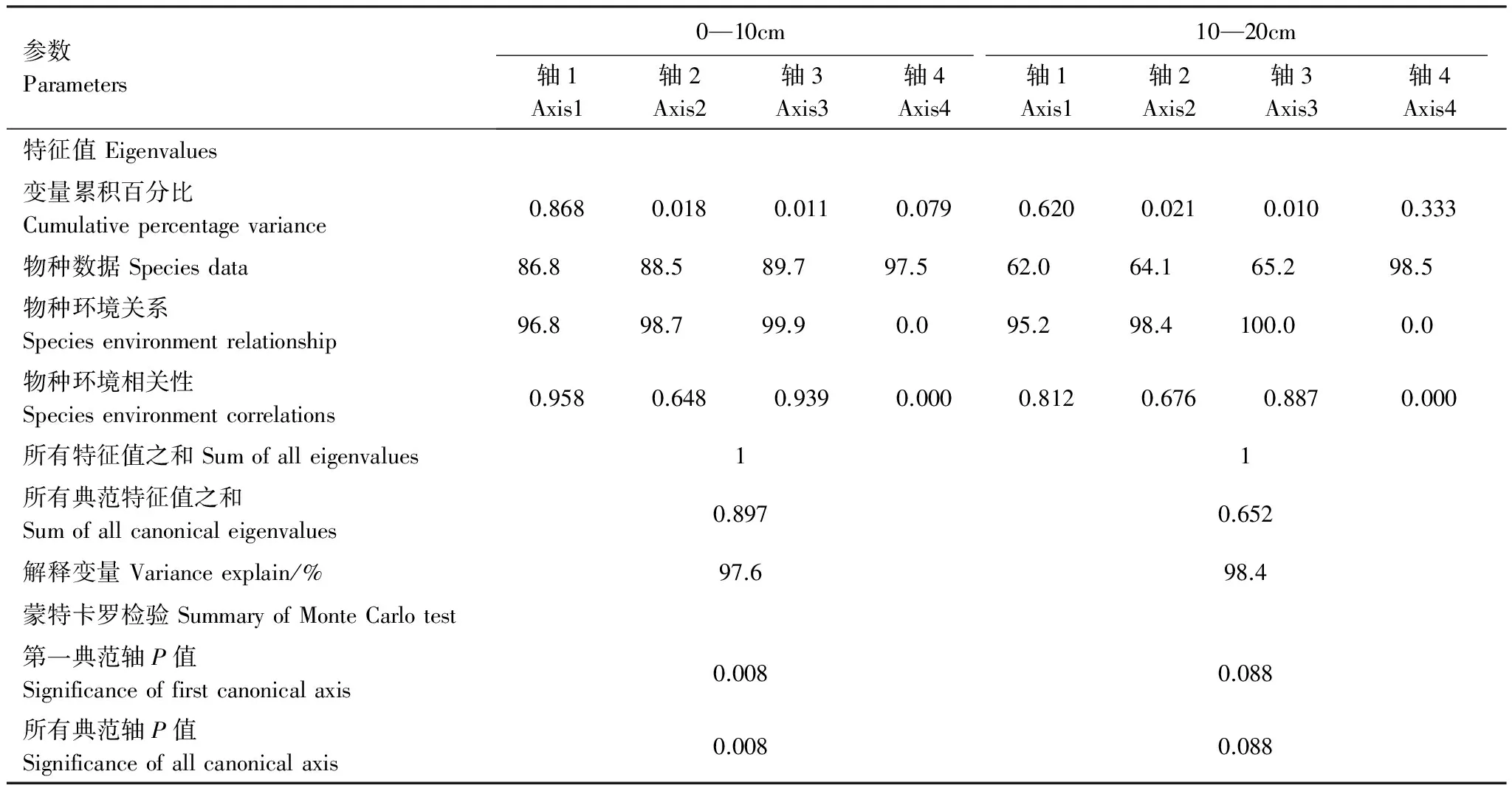
表2 植物多样性和土壤养分的RDA分析结果
不同植物多样性指数与土壤养分间的相关性在不同高原鼠兔干扰水平下是不一致的。植物多样性指数(H)与0—10cm土壤深度全氮(TN)间的相关性从T1干扰水平时的极显著负相关(P<0.01)(图3),转变为T2和T3时的负相关(P>0.05),至T4时则变为极显著正相关(P<0.01),与0—10cm土壤深度硝态氮(NN)间的相关性从T1干扰水平时的正相关(P>0.05)转变为T2和T3时的显著正相关(P<0.05),至T4时则变为极显著负相关(P<0.01),与0—10cm土壤深度铵态氮(AN)间的相关性从T1干扰水平时的负相关(P>0.05),转变为T2、T3和T4时的显著正相关(P<0.05);植物丰富度指数(R)与0—10cm土壤深度全氮(TN)间的相关性从T1干扰水平时的负相关(P>0.05)转变为T2时的极显著正相关(P<0.01)和T3时的极显著负相关(P<0.01),至T4时则变为显著正相关(P<0.05),与0—10cm土壤深度有机碳(OC)间的相关性从T1到T3时的极显著负相关(P<0.01)转变为T4时的极显著正相关(P<0.01),与0—10cm土壤深度硝态氮(NN)间的相关性从T1干扰水平时的极显著正相关(P<0.01)转变为T2时的极显著负相关(P<0.01)和T3时的正相关(P>0.05),至T4时则变为负相关(P>0.05),与0—10cm土壤深度铵态氮(AN)间的相关性从T1干扰水平时的极显著负相关(P<0.01)转变为T2到T4时的显著正相关(P<0.05);植物均匀度指数(E)与0—10cm土壤深度全磷(TP)间的相关性从T1干扰水平时的极显著正相关(P<0.01)转变为T2时的显著负相关(P<0.05)和T3时的正相关(P>0.05),至T4时则变为负相关(P>0.05),与0—10cm土壤深度速效钾(AK)间的相关性从T1干扰水平时的显著负相关(P<0.05)转变为T2时的负相关(P>0.05)和T3时的正相关(P>0.05),至T4时则为极显著正相关(P<0.01),与0—10cm土壤深度速效磷(AP)间的相关性从T1和T2干扰水平时的极显著正相关(P<0.01)转变为T3和T4时的负相关(P>0.05)。
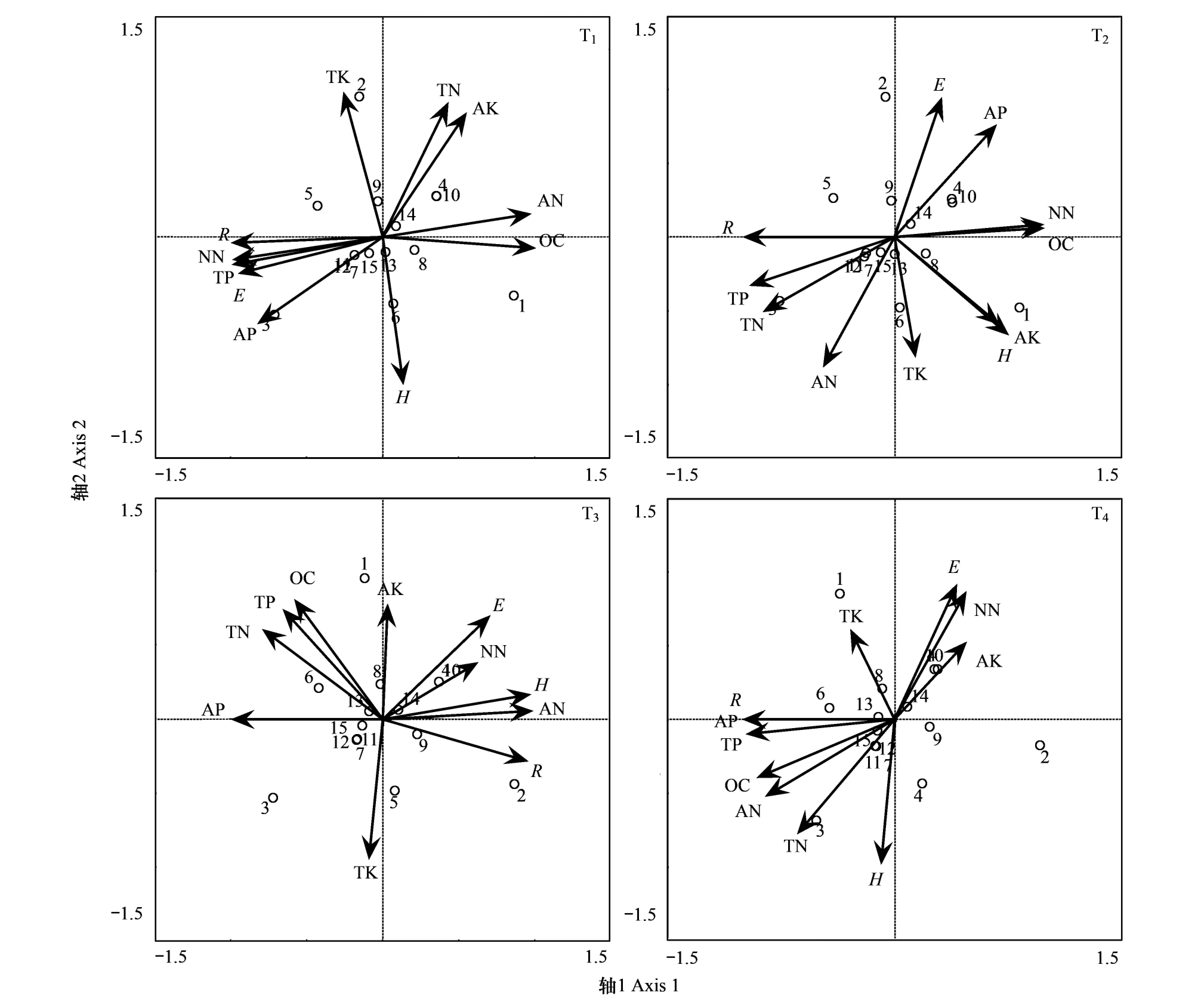
图3 高原鼠兔不同干扰梯度下植物多样性与0—10cm土壤养分的RDA排序图Fig.3 RDA ordination of plant diversity and soil nutrients at 0—10 cm soil depth in different degrees of plateau pikaH:多样性指数 Shannon-Weiner index;E: Pielou均匀度指数 Pielou evenness index;R:丰富度指数 Margalef index;TN:土壤全氮 Soil total nitrogen;AN:土壤铵态氮 Soil ammonium nitrogen;NN:土壤销态氮Soil nitrate nitrogen;OC:土壤有机碳 Soil organic carbon;TK:土壤全钾 Soil total potassium;AK:土壤速效钾 Soil available potassium;TP:土壤全磷 Soil total phosphorous;AP:土壤速效磷 Soil available phosphorous
植物多样性指数(H)与10—20cm土壤深度全钾(TK)间的相关性从T1到T3时的显著正相关(P<0.05)转变为T4时的极显著负相关(P<0.01)(图4),与10—20cm土壤深度速效磷(AP)间相关性从T1干扰水平时的极显著正相关(P<0.01)转变为T2时的显著正相关(P<0.05)和T3时的极显著负相关(P<0.01),至T4时则为极显著负相关(P<0.01);植物丰富度指数(R)与10—20cm土壤深度全磷(TP)间相关性从T1干扰水平时的极显著正相关(P<0.01)转变为T2时的显著负相关(P<0.05)和T3时的正相关(P>0.05),至T4时则为显著负相关(P<0.05),与10—20cm土壤深度硝态氮(NN)间相关性从T1干扰水平时的显著正相关(P<0.05)转变为T2时的显著负相关(P<0.05),至T3和T4时则为极显著负相关(P<0.01),与10—20cm土壤深度有机碳(OC)间相关性从T1干扰水平时的显著负相关(P<0.05)转变为T2时的显著正相关(P<0.05),至T3和T4时则为正相关(P>0.05);植物均匀度指数(E)与10—20cm土壤深度全氮(TN)间相关性从T1干扰水平时的显著负相关(P<0.05)转变为T2时的正相关(P>0.05)和T3时的极显著负相关(P<0.01),至T4时则为正相关(P>0.05),与10—20cm土壤深度全钾(TK)、速效钾(AK)间相关性从T1干扰水平时的负相关(P>0.05)转变为T2时的显著负相关(P<0.05)和T3时的极显著正相关(P<0.01),至T4时则为极显著正相关(P<0.01),与10—20cm土壤深度有机碳(OC)间的相关性从T1干扰水平时的显著负相关(P<0.05)转变为T2和T3时的负相关(P>0.05),至T4时则为显著正相关(P<0.05),与10—20cm土壤深度铵态氮(AN)间的相关性从T1干扰水平时的正相关(P>0.05),转变为T2时的正相关(P>0.05)和T3时的极显著负相关(P<0.01),至T4时则为负相关(P>0.05)。

图4 高原鼠兔不同干扰梯度下植物多样性与10—20cm土壤养分的RDA排序图Fig.4 RDA ordination of plant diversity and soil nutrients at 10—20 cm soil depth in different degrees of plateau pika
3 讨论
高原鼠兔作为青藏高原高寒草甸生态系统内群居性的小型哺乳类动物,其通过掘洞和采食行为干扰高寒草甸的生境,对植物多样性和土壤养分产生深刻的影响,这种影响依赖于高原鼠兔的干扰程度。本研究结果表明,高原鼠兔干扰水平为T1、T2、T3时(小于或等于22个/625m2)不会降低高寒草甸植物丰富度和均匀度,T1和T2时(小于或等于12个/ 625m2)维持了植物多样性,然而当高原鼠兔干扰水平为T4时(38个/625m2),植物多样性明显降低,这是因为高原鼠兔干扰程度从T1增加到T2时,土壤含水量[30]和主要物种高山嵩草及伴生种矮火绒草等的盖度、密度差异不明显[32],表明高寒草甸生境变化很小,因此植物多样性变化不大。当干扰程度增加到T3时,植物多样性具有降低趋势,这是因为高寒草甸内裸斑数明显增多,即高原鼠兔数量增加、干扰水平增加,其对喜食牧草的啃食加大,对土壤的掘土程度增加,致使植被盖度降低,形成大小不一的裸地。裸斑窗数量的增加,改变了样地内水热过程[33],土壤含水量明显降低[26],促进草地生境从湿生向中生转变,从而影响植物群落的动态变化,珠芽蓼等湿生植物退出植物群落,青藏苔草[34]、矮火绒草、大耳叶风毛菊[35]等湿中生植物长势减弱,重要值降低,导致植物多样性整体降低,这与南非Wayland Flower自然保护区的高山硬叶灌木丛(Fynbos)植物多样性对鼹形鼠(Bathyergidae)的响应一致[36],即随鼹形鼠干扰强度增加,植物物种丰富度降低,说明当地面啮齿类动物干扰强度较小时,湿生植物栖息于草甸植被大面积覆盖生境,中旱生植物栖息于裸斑或草甸植被覆盖区的边缘,然而随着裸斑数量增加,大面积草甸植被覆盖区逐渐被镶嵌分布的裸斑碎化,湿生植物因不适应环境而退出群落,草甸主要由中旱生植物组成,迫使植物多样性降低。
高原鼠兔干扰强度的改变导致高寒草甸生境发生变化,势必引起土壤养分含量的改变。本研究发现,土壤有机碳含量随着高原鼠兔干扰程度增加呈现先增加后降低的态势,这与青海果洛地区高原鼠兔干扰引起土壤有机碳变化的趋势一致[37],其原因可能是高原鼠兔同大多数地面啮齿类动物对草地的干扰类似,当它们干扰强度适当时,其掘洞活动疏松了表层土壤结构[38],增加了毛细管持水量及土壤总空隙度[19],表层土壤湿度的增加促进了有机物分解速率,提高土壤表层有机碳含量,而当地面啮齿动物鼠类干扰强度过大时,地表植被盖度下降[26,38],迫使土壤有机质的输入源减少,一定程度上降低了土壤有机碳含量。高原鼠兔干扰对高寒草甸土壤全氮和速效氮的影响出现明显的分异,其中土壤全氮和硝态氮随干扰梯度增加呈降低态势,而铵态氮随干扰梯度增加却表现为先降低后增加的趋势。土壤全氮降低主要原因是当啮齿类动物干扰增大时,一方面其消耗的牧草量增加,降低植物地上生物量[30,37]和凋落物归还生态系统的量[39],客观上减少了土壤氮素的来源;另一方面裸斑数量的增加会增加土壤侵蚀,从而导致土壤全氮含量下降。土壤硝态氮和铵态氮对高原鼠兔干扰的响应存在明显的分异,当高原鼠兔干扰从T1增加到T2的过程中,禾本科植物初级生产力较大[13],因此土壤中的铵态氮和硝态氮因禾本科植物吸收利用而逐渐降低;而当高原鼠兔干扰程度达T3后,禾本科植物生物量降低,这虽然降低了植物对硝态氮和铵态氮的消耗量,但铵态氮含量表现为增加,硝态氮含量持续降低,原因可能是随着土壤裸斑面积增加,土壤侵蚀量增加,铵态氮因带正电荷被土壤胶体所吸附固定于土壤中因此含量增加,而硝态氮因带负电荷不易被土壤胶体吸附更容易淋失[40],导致其含量持续降低。土壤全磷含量随高原鼠兔干扰程度的增加而先增加后降低,说明适宜的高原鼠兔干扰能够增加土壤全磷含量,而过高或过低的高原鼠兔干扰均可降低土壤全磷含量,这可能因为虽然土壤中全磷主要来源于成土母质,但有机质积累是除母质全磷以外的又一影响青藏高原地区土壤磷含量的重要因素[41- 43]。而本研究结果中有机质的含量呈现出先增加后降低的趋势,且由于高原鼠兔干扰使植物群落组成发生改变,其也会导致土壤磷素含量分布上的差异[42],因此,土壤全磷含量随高原鼠兔干扰程度增加的变化趋同于土壤有机碳。
草地植物多样性与土壤环境因子间具有一定的相关性[6,9],这是草地生态系统维持相对稳定的基础。赵景学等[44]的研究结果表明,青藏高原藏北地区高寒草甸植物多样性与土壤有机质、全氮、速效氮和全磷含量呈显著的正相关关系。本研究结果表明,高寒草甸植物多样性与土壤养分间的相关性随着高原鼠兔干扰水平不同而发生明显的变化,表现为植物多样性指数与土壤养分间的关系从T1时的正相关或负相关转变为T2或T3之后的负相关或正相关,譬如植物多样性指数与0—10cm土壤深度硝态氮的相关性从T1到T3时的正相关转变为T3之后(T4)时的负相关,与0—10cm土壤深度全氮的相关性则从T1到T3时的负相关转变为T3之后(T4)的正相关,而植物多样性指数与0—10cm土壤深度铵态氮的相关性从T1时的负相关转变为T2时的正相关,说明高原鼠兔干扰改变了植物多样性指数与土壤养分间的相关性,而植物多样性指数与土壤养分间相关性发生变化时而对应的高原鼠兔干扰水平介于T2(12个/625 m2)和T3(22个/625 m2)之间。植物多样性指数是物种数与均匀度结合起来的一个单一统计量[45],因此植物多样性指数与土壤养分间相关性的变化源于植物丰富度和植物均匀度与土壤养分间关系的变化。植物丰富度指数与土壤养分间的关系从高原鼠兔干扰水平为T1时的正相关或负相关转变为干扰水平为T2或T3之后的负相关或正相关,例如植物丰富度指数与0—10cm土壤深度有机碳间的相关性从高原鼠兔干扰水平为T1到T3时的负相关转变为干扰水平为T4时的正相关,植物均匀度指数与0—10cm土壤深度速效磷间的相关性从高原鼠兔干扰水平为T1到T2时的正相关转变为T3和T4时的负相关,主要是高寒草甸植物群落组分和生境异质性随着高原鼠兔干扰水平增加而发生明显变化,表现为随着高原鼠兔干扰水平增加,裸斑数量增加,生境异质性增加,同时湿生植物适应性降低,而中旱生植物适应性增加。因此,高原鼠兔干扰程度的变化,改变了高寒草甸土壤养分和植物丰富度指数和均匀度指数间的关系,导致土壤养分与植物多样性指数间的关系发生了变化。然而高原鼠兔在青藏高原地区分布的海拔大约为3200—5200m之间[15,24,27],不同海拔带高原鼠兔干扰对植物多样性与土壤养分间的关系影响是否趋同,有待深入研究。
[1] 周华坤, 赵新全, 周立, 刘伟, 李英年, 唐艳鸿. 青藏高原高寒草甸的植被退化与土壤退化特征研究. 草业学报, 2005, 14(3): 31- 40.
[2] 许岳飞, 益西措姆, 付娟娟, 陈浩, 苗彦军, 陈俊, 呼天明, 镡建国. 青藏高原高山嵩草草甸植物多样性和土壤养分对放牧的响应机制. 草地学报, 2012, 20(6): 1026- 1032.
[3] Luo Y Q, Su B, Currie W S, Ducks J S, Finzi A, Hartwig U, Hungate B, McMurtrie R E, Oren R, Parton W J, Pataki D E, Shaw R M, Zak D R, Field C B. Progressive nitrogen limitation of ecosystem responses to rising atmospheric carbon dioxide. BioScience, 2004, 54(8): 731- 739.
[4] 王长庭, 龙瑞军, 王根绪, 刘伟, 王启兰, 张莉, 吴鹏飞. 高寒草甸群落地表植被特征与土壤理化性状、土壤微生物之间的相关性研究. 草业学报, 2010, 19(6): 25- 34.
[5] 蒋婧, 宋明华. 植物与土壤微生物在调控生态系统养分循环中的作用. 植物生态学报, 2010, 34(8): 979- 988.
[6] Janssens F, Peeters A, Tallowin J R B, Bakker J P, Bekker R M, Fillat F, Oomes M J M. Relationship between soil chemical factors and grassland diversity. Plant and Soil, 1998, 202(1): 69- 78.
[7] van der Putten W H, Bardgett R D, Bever J D, Martijn Bezemer T, Casper B B, Fukami T, Kardol P, Klironomos J N, Kulmatiski A, Schweitzer J A, Suding K N, van de Voorde T F J, Wardle D A. Plant-soil feedbacks: the past, the present and future challenges. Journal of Ecology, 2013, 101(2): 265- 276.
[8] Cong W F, van Ruijven J, Mommer L, de Deyn G B, Berendse F, Hoffland E. Plant species richness promotes soil carbon and nitrogen stocks in grasslands without legumes. Journal of Ecology, 2014, 102(5): 1163- 1170.
[9] Pohl M, Alig D, Körner C, Rixen C. Higher plant diversity enhances soil stability in disturbed alpine ecosystems. Plant and Soil, 2009, 324(1): 91- 102.
[10] 闫东峰. 太行山低山丘陵区不同植被恢复措施下植被与土壤协同演替机制[D]. 郑州: 河南农业大学, 2012.
[11] 贾婷婷, 毛亮, 郭正刚. 高原鼠兔有效洞穴密度对青藏高原高寒草甸群落植物生态位的影响. 生态学报, 2014, 34(4): 869- 877.
[12] 王长庭, 龙瑞军, 王启兰, 景增春, 施建军, 杜岩功, 曹广民. 三江源区高寒草甸不同退化演替阶段土壤有机碳和微生物量碳的变化. 应用与环境生物学报, 2008, 14(2): 225- 230.
[13] 李倩倩, 赵旭, 郭正刚. 高原鼠兔有效洞穴密度对高寒草甸优势植物叶片和土壤氮磷化学计量特征的影响. 生态学报, 2014, 34(5): 1212- 1223.
[14] 贾婷婷, 袁晓霞, 赵洪, 杨玉婷, 罗开嘉, 郭正刚. 放牧对高寒草甸优势植物和土壤氮磷含量的影响. 中国草地学报, 2013, 35(6): 80- 85.
[15] 赵国琴, 李广泳, 马文虎, 赵殿智, 李小雁. 高原鼠兔扰动对高寒草地植物群落特征的影响. 应用生态学报, 2013, 24(8): 2122- 2128.
[16] 孙飞达, 郭正刚, 尚占环, 龙瑞军. 高原鼠兔洞穴密度对高寒草甸土壤理化性质的影响. 土壤学报, 2010, 47(2): 378- 383.
[17] Smith A T, Foggin J M. The plateau pika (Ochotonacurzoniae) is a keystone species for biodiversity on the Tibetan plateau. Animal Conservation, 1999, 2(4): 235- 240.
[18] 尚占环, 龙瑞军. 青藏高原“黑土型”退化草地成因与恢复. 生态学杂志, 2005, 24(6): 652- 656.
[19] 李文靖, 张堰铭. 高原鼠兔对高寒草甸土壤有机质及湿度的作用. 兽类学报, 2006, 26(4): 331- 337.
[20] Arthur A D, Pech R P, Davey C, Jiebu, Zhang Y M, Lin H. Livestock grazing, plateau pikas and the conservation of avian biodiversity on the Tibetan Plateau. Biological Conservation, 2008, 141(8): 1972- 1981.
[21] Guo Z G, Zhou X R, Hou Y. Effect of available burrow densities of plateau pika (Ochotonacurzoniae) on soil physicochemical property of the bare land and vegetation land in the Qinghai-Tibetan Plateau. Acta Ecologica Sinica, 2012, 32(2): 104- 110.
[22] Bagchi S, Namgail T, Ritchie M E. Small mammalian herbivores as mediators of plant community dynamics in the high-altitude arid rangelands of Trans-Himalaya. Biological Conservation, 2006, 127(4): 438- 442.
[23] Guo Z G, Li X F, Liu X Y, Zhou X R. Response of alpine meadow communities to burrow density changes of plateau pika (Ochotonacurzoniae) in the Qinghai-Tibet Plateau. Acta Ecologica Sinica, 2012, 32(1): 44- 49.
[24] 严红宇, 张毓, 赵建中, 许庆民, 李里, 刘伟. 高原鼠兔对高寒草甸植物群落生物量的影响. 兽类学报, 2013, 33(4): 333- 343.
[25] 孙飞达, 龙瑞军, 郭正刚, 刘伟, 干友民, 陈文业. 鼠类活动对高寒草甸植物群落及土壤环境的影响. 草业科学, 2011, 28(1): 146- 151.
[26] Liu Y S, Fan J W, Harris W, Shao Q Q, Zhou Y C, Wang N, Li Y Z. Effects of plateau pika (Ochotonacurzoniae) on net ecosystem carbon exchange of grassland in the Three Rivers Headwaters region, Qinghai-Tibet, China. Plant and Soil, 2013, 366(1/2): 491- 504.
[27] 刘伟, 严红宇, 王溪, 王长庭. 高原鼠兔对退化草地植物群落结构及恢复演替的影响. 兽类学报, 2014, 34(1): 54- 61.
[28] 孙飞达, 龙瑞军, 路承香. 高原鼠兔不同洞穴密度对高寒草地植物群落组成及多样性的影响. 干旱区资源与环境. 2010, 24(7): 181- 186.
[29] 张军. 高原鼠兔种群数量对植被调控措施的响应[D]. 兰州: 兰州大学, 2011.
[30] 石红霄, 于健龙. 高原鼠兔洞口密度对高寒嵩草草甸植被及土壤水分的影响. 中国草地学报, 2010, 32(4): 109- 112, 116- 116.
[31] 鲁如坤. 土壤农业化学分析方法. 北京: 中国农业科技出版社, 2000.
[32] 庞晓攀, 贾婷婷, 李倩倩, 雒眀伟, 肖玉, 赵旭, 郭正刚. 高原鼠兔有效洞穴密度对高山嵩草群落及其主要种群空间分布特征的影响. 生态学报, 2015, 35(3): 873- 884.
[33] Archer S, Detling J K. Evaluation of potential herbivore mediation of plant water status in a North American mixed-grass prairie. Oikos, 1986, 47(3): 287- 291.
[34] 李中强, 张萌, 徐军. 青藏高原纳木错水生植物多样性及群落生态学研究. 水生生物学报, 2011, 35(1): 1- 13.
[35] 武高林, 陈敏, 杜国祯. 营养和光照对不同生态幅风毛菊属植物幼苗形态可塑性的影响. 应用生态学报, 2008, 19(8): 1708- 1713.
[36] Hagenah N, Bennett N C. Mole rats act as ecosystem engineers within a biodiversity hotspot, the Cape Fynbos. Journal of Zoology, 2013, 289(1): 19- 26.
[37] 周雪荣. 青藏高原高寒草甸群落和土壤对高原鼠兔密度变化的响应[D]. 兰州: 兰州大学, 2010.
[38] Galiano D, Kubiak B B, Overbeck G E, de Freitas T R O. Effects of rodents on plant cover, soil hardness, and soil nutrient content: a case study on tuco-tucos (Ctenomysminutus). Acta Theriologica, 2014, 59(4): 583- 587.
[39] Smith J G, Eldridge D J, Throop H L. Landform and vegetation patch type moderate the effects of grazing-induced disturbance on carbon and nitrogen pools in a semi-arid woodland. Plant and Soil, 2012, 360(1): 405- 419.
[40] 孙志高, 刘景双, 牟晓杰. 三江平原小叶章湿地土壤中硝态氮和铵态氮含量的季节变化特征. 农业系统科学与综合研究, 2010, 26(3): 277- 282.
[41] 高丽丽. 西藏土壤有机质和氮磷钾状况及其影响因素分析[D]. 成都: 四川农业大学, 2004.
[42] 谭鑫. 青藏高原东缘高寒地区土壤磷素空间分布研究[D]. 成都: 四川师范大学, 2009.
[43] 裴海昆, 朱志红, 乔有明, 李希来, 孙海群. 不同草甸植被类型下土壤腐殖质及有机磷类型探讨. 草业学报, 2001, 10(4): 18- 23.
[44] 赵景学, 曲广鹏, 多吉顿珠, 尚占环. 藏北高寒植被群落物种多样性与土壤环境因子的关系. 干旱区资源与环境, 2011, 25(6): 105- 108.
[45] 郭正刚, 王根绪, 沈禹颖, 程国栋. 青藏高原北部多年冻土区草地植物多样性. 生态学报, 2004, 24(1): 149- 155.
The effect of plateau pika(Ochotona curzoniae) disturbance on the relationship between plant diversity and soil nutrients of alpine meadow
WANG Ying, PANG Xiaopan, XIAO Yu, JIA Tingting, WANG Qian, YU Cheng, GUO Zhenggang*
State Key Laboratory of Grassland Agro-ecosystems, College of Pastoral Agriculture Science and Technology, Lanzhou University, Lanzhou 730020, China
Although plant diversity and soil nutrients content can be affected by plateau pika disturbance, the relationships between plant diversity and soil nutrients of alpine meadows in response to plateau pika disturbance are still unclear. RDA redundancy analysis was used to determine the effect of plateau pika disturbance on the relationship between plant diversity and soil nutrients of alpine meadow by using the active burrow entrance density of plateau pika to substitute for the disturbance levels of plateau pika, in which the disturbance consisted of four levels with density of T1(7 active burrows/625 m2), T2(12 active burrows /625 m2), T3(22 active burrows /625 m2) and T4(38 active burrows /625 m2). This study showed that the important value ofKobresiapygmaea, dominant plant, firstly increased and then decreased with the increase of plateau pika disturbance level, while the important value ofAnemonerivularisvar.flore-minorsandPotentillafragarioides, main associate plants, firstly decreased and then increased with the increase of plateau pika disturbance level. The plant diversity index increased when the plateau pika disturbance increased from T1level to T2level, and showed a decreasing trend when the plateau pika disturbance exceeded T2level. Soil total nitrogen and nitrate nitrogen showed a decreasing trend, and ammonium nitrogen firstly decreased and then increased as the plateau pika disturbance increased, while soil organic carbon and total phosphorus firstly increased and then decreased in the increasing process of the plateau pika disturbance. RDA analysis results showed that the correlation relationships between plant diversity index and nitrate nitrogen at 0—10cm soil depth and potassium at 10—20cm soil depth was significantly positive when plateau pika disturbance ranged from T1level to T3level, and it became negative when plateau pika disturbance was T4level, while the correlation relationships between plant diversity index and total nitrogen at 0—10cm soil depth was significantly negative when plateau pika disturbance increased from T1level to T3level, and it became positive when plateau pika disturbance reached to T4level. The relationship between plant diversity index and ammonium nitrogen at 0—10cm soil depth was negative correlation at T1level environment and it became positive at T2, T3, T4levels. These results indicated that the plateau pika disturbance changed the relationships between plant diversity index and soil nutrients of alpine meadow, and the sensitive disturbance of plateau pika on relationships between plant diversity index and soil nutrients was T2level and T3level.
plateau pika; plant diversity; collaborative; soil nutrients
国家行业(农业)公益资助项目(201203041);国家自然科学基金资助项目(31172258)
2015- 02- 07;
日期:2015- 12- 14
10.5846/stxb201502070311
*通讯作者Corresponding author.E-mail: guozhg@lzu.edu.cn
王莹,庞晓攀,肖玉,贾婷婷,王 倩,于成,郭正刚.高原鼠兔干扰对高寒草甸植物多样性与土壤养分间关系的影响.生态学报,2016,36(17):5485- 5496.
Wang Y, Pang X P, Xiao Y, Jia T T, Wang Q, Yu C, Guo Z G.The effect of plateau pika(Ochotonacurzoniae) disturbance on the relationship between plant diversity and soil nutrients of alpine meadow.Acta Ecologica Sinica,2016,36(17):5485- 5496.
