内蒙古贺兰山某块菌的形态解剖学与分子生物学研究
2016-11-04赵艳玲王海峰王兆锭樊永军
赵艳玲,王海峰,王兆锭,樊永军*
(1 包头师范学院 生物科学与技术学院,内蒙古包头 014030;2 包头轻工职业技术学院 食品药品工程学院, 内蒙古包头 014035;3 内蒙古贺兰山国家级自然保护区管理局,内蒙古阿拉善 750306)
内蒙古贺兰山某块菌的形态解剖学与分子生物学研究
赵艳玲1,王海峰2,王兆锭3,樊永军1*
(1 包头师范学院 生物科学与技术学院,内蒙古包头 014030;2 包头轻工职业技术学院 食品药品工程学院, 内蒙古包头 014035;3 内蒙古贺兰山国家级自然保护区管理局,内蒙古阿拉善 750306)
利用形态解剖学和分子生物学方法,对采自内蒙古贺兰山地区实验样地青海云杉林下的块菌两菌株(菌株a和b)进行分析鉴定。研究发现:(1)两菌株子囊果均为黄褐色,表面光滑,没有明显的疣状突起和棱角。(2)菌株a产孢组织乳白色、致密团状,菌肉组织褐色;球形、棒状子囊呈蜂窝状排布,内含有1~4个带包被的、表面具有突起状纹饰的球型子囊孢子;子囊孢子双层壁,厚约1.7 μm,直径约20 μm(含纹饰)。(3)菌株b产孢组织有裂隙,松散,子实层内除了具有上述蜂窝状排布的子囊和内部的球型孢子外,还具有“口袋”状子囊,该子囊内含有大量两端尖、外壁光滑、褐色的椭球型孢子。(4)分子生物学进化分析表明,两菌株聚为一支,但属于块菌属的支持率相对较低;推断两菌株可能为中国猪块菌属Choiromyces新记录种。
贺兰山;块菌; 子囊果;子囊;子囊孢子;解剖学;进化树
近年来,关于块菌(Tuber)的采集、分类和鉴定工作一直是国内学者研究的热点问题[1-4]。王云[1]认为东亚尤其是中国的块菌属资源丰富且独特,尚存在大量的未被发现或描述的新分类群。同时指出块菌主要分布于中国的东北到西南地区这一条带上。有研究者对中国块菌属进行研究,其块菌材料主要分布在吉林、辽宁、北京、内蒙古赤峰、山西、成都、昆明和西藏等地[5-6]。陈应龙认为,在中国块菌主要分布在西南的四川和云南,以及新疆、西藏、山西、辽宁、吉林、福建等省区[7]。目前,鲜见关于贺兰山块菌资源的报道。贺兰山脉位于宁夏回族自治区与内蒙古自治区交界处。山脉为近南北走向,具有日照充足,气候多变等特点[8]。由于其特殊的地理环境和生态因子, 孕育了垂直带变化明显的植物类群,同时为多种大型真菌的生长发育提供了十分有利的条件[9]。本研究针对采自内蒙古贺兰山地区的真菌材料进行形态解剖学和分子生物学分析,以期为丰富贺兰山真菌资源多样性和中国块菌的系统分类研究奠定基础。
1 材料和方法
1.1材料
两真菌试验材料于2015年1月采自贺兰山地区实验样地青海云杉林下,并暂分类为菌株a和b。两菌株采集地理信息为:海拔2 757 m, E105°57′12.90″,N38°11′27.24″。
1.2研究方法
1.2.1形态解剖学分类1) 外部形态观察。观察内容主要包括子囊果的外部形态、颜色;菌体表面是否具疣状突起;菌脉的密集程度、粗细等特征。
2) 内部显微观察。观察内容包括:包被、产孢组织的颜色和结构等。同时,在解剖镜(Nikon-JSZ5)下观察拍照。
3) 扫描电镜观察。菌株经3%戊二醛前固定24 h,吸除固定液,加入0.1 mol/L磷酸缓冲液(pH 7.2)漂洗2~3次,再用1%锇酸固定2 h,吸除固定液,以双蒸水洗2 h(中间换3次新液);用50%、70%、80%、90%乙醇逐级脱水,每次20 min,吸除乙醇,再用无水乙醇脱水两次,每次20 min;将菌株取出用日立HCP-2临界点干燥仪进行CO2临界点干燥,用日立IB-5真空离子溅射仪进行喷金处理,用日立S-570扫描电子显微镜和Hitachi S-4800场发射扫描电子显微镜观察并拍照。观察记录子囊的形状、大小以及子囊内孢子的数目;子囊孢子的形状、大小;表面的纹饰(刺、网格或刺-网格)等。
1.2.2分子生物学研究采用常规CTAB 法提取两菌株基因组DNA,以ITS1f(5′-CTTGGTCATTTAGAGGAAGTAA-3′)和ITS4(5′-TCCTCCGC-TTATTGATATGC-3′)为引物,进行PCR扩增。PCR反应参数为:50 μL体系,94 ℃预变性 5 min,94 ℃变性 40 s, 57 ℃退火1 min, 72 ℃延伸1 min, 72 ℃ 10 min,共30个循环。PCR扩增产物电泳后,用Biospin真菌回收试剂盒割胶回收纯化,纯化产物交由生工上海测序部测序。
2 结果与分析
2.1外部形态观察
两菌株子囊果近球形或椭球形,未见子囊柄。表面光滑,不具有疣状突起,呈红褐色,有不规则的沟状凹陷。子囊果直径约2~4 cm。新鲜子囊果具特殊香气(图1,A、B)。马麝(Moschuschrysogaster)喜食该菌体[10]。
2.2内部显微结构观察
2.2.1子实体横切面观察由两菌株子囊果的横切面可以看出,菌体外观呈黄褐色,光滑,无明显的疣状突起;菌体边缘具薄的包被;包被内子实层具迷走状脉纹和褐色菌肉组织。菌株a的子实层致密,呈乳黄色大理石菌脉封闭,迷走团状,在多处可到达包被边缘处(图1,C)。菌株b子实层较松散,在子实层内肉眼可见黑色颗粒状子囊,菌脉同样封闭,呈迷走状(图1,D)。
菌株a子实层内形成浅黄色脉纹(明纹),并与外围不育菌丝形成的菌肉组织形成的暗纹呈明显对比。菌株b侧丝构成的菌肉组织较窄;子实层中有裂隙。分析原因,可能是由于产孢组织中不断发育的子囊和子囊孢子不断挤压侧丝造成。
2.2.2扫描电镜观察由图2扫描电镜照片可见,菌株a产孢组织内具大量蜂窝状分布的棒状、球形子囊(图2,A)。单个成熟子囊中常含1~4个子囊孢子(图2,B),子囊孢子球形、游离状分布、双层壁,壁厚约1.7 μm(不含纹饰,图2,C)。外壁上每3~4个呈圆形坑状的网眼交汇处形成一个具有明显的、完整的、均匀的指状突起(图2,D)。由图2,B可见,子囊内的子囊孢子具有膜状的包被。龙月娟[11]研究印度块菌子囊果的发育认为,具有双层膜的子囊孢子在发育后期,外膜分化断开与内膜共同形成双层膜,且外膜在分化、断裂之前就有明显的刺状突起纹饰。电镜照片中的孢子(图2,B)不属于这一发育阶段。因为该菌株子囊内的膜(图2,B)是平整光滑的,且包裹着的子囊孢子已经具有突起状纹饰。可见,在该阶段子囊孢子的内膜和带纹饰的次生壁已经形成,说明子囊孢子已经发育完成,而该膜应属于子囊膜。
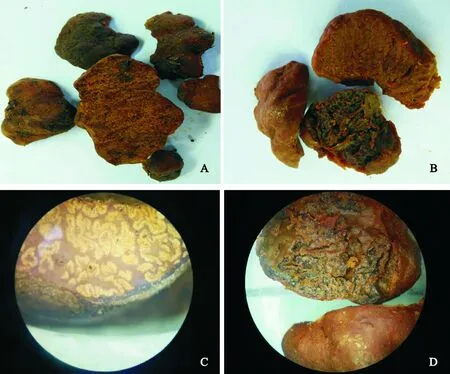
图1 菌株a(A,C)和b(B,D)子囊果的形态结构和切面图Fig.1 The morphological features and the section of strain a and b

A.菌体表面扫描(×60);B.子囊和带有包被的子囊孢子(×1 000);C.子囊孢子的双层壁(×1 000);D.子囊孢子的突起状纹饰(×2 457)图2 菌株a扫描电镜系列照片A.Surface scanning, ×60; B.Sac and ascospore with package, ×1 000; C.The scanning a double wall of ascospore, ×1 000; D.Protrusions ornamentation of ascospore,×2 457Fig.2 Photos of strain a under scanning electron microscope (SEM)

A.表面扫描结构(×150);B.子囊孢子及子囊扫描结构(×600);C.子囊果内呈蜂窝状子囊及球型、椭球形子囊孢子(×300);D.椭球形的子囊孢子(×3000)图3 菌株b扫描电镜系列照片A.Surface scanning,×150; B.Sac like pocket,×600; C. The scanning of ascus like honeycomb and spherical and ellipsoidal ascospore with SEM,×300; D. Amplification of ellipsoid ascospore,×3 000Fig.3 Photos of strain a under scanning electron microscope (SEM)
由图3扫描电镜照片可见,菌株b产孢组织内子囊的形状较不规则。除了呈蜂窝状分布的棒状、圆球状子囊外,还有似“口袋”结构的子囊。蜂窝状分布的子囊内含有表面具疣状突起和网纹结构的球型、近球形子囊孢子;而似“口袋”结构的子囊内含有大量褐色、两端呈“芒果尖”状的椭球型孢子(图3,D)。
综合以上结果可知,菌株a子囊果内的单个子囊中含有1~4个子囊孢子,常见3~4个子囊孢子。子囊(48)~74~(98) μm×(41)~48~(53) μm,具有薄膜状包被的球型子囊孢子的直径约为18.3~20~22.0 μm (含子囊孢子的纹饰),子囊孢子纹饰的突起高(1)~2~(2.5) μm。电镜下,突起低于2 μm的纹饰呈乳突状,高于2 μm的纹饰呈指状突起。所以该菌株的子囊孢子外壁上的纹饰特征以指状或者乳突状的纹饰为主。
菌株b,菌体内主要有两种类型的子囊。蜂窝状分布的棒状子囊内含1~4个子囊孢子,常见3~4个子囊孢子,子囊(42)~63~(88) μm×(38)~42~(61) μm,球型子囊孢子直径(含表面突起状的纹饰)(15)~20~(22) μm,该子囊里的子囊孢子的纹饰与菌株a的极为相似;菌株b子囊果内的似“口袋”的子囊囊径(59)~96~(139) μm,该子囊内存在大量的没有子囊包被的两端呈“芒果尖”、表面光滑、褐色椭球形子囊孢子(17)~21~(25) μm×(12)~14~(16) μm。
2.3分子生物学分析
采用CTAB法提取的两菌株基因组DNA经PCR扩增,纯化后测序,将测得的两菌株的序列经Chromas软件进行序列校准,运用GenBank 的序列局部相似性查询系统(Blast),引用 DNA 基因库中同源菌株rDNA的ITS(Internal Transcribed Spacer,内转录间隔区)区段序列,然后运用软件MEGA5.1 采用Neighbor-Joining Tree(相邻比对法)分别对两供试菌种进行进化树的构建(图4),由图可知,菌株a和b被聚在一起,且与Tubertaiyuanense(太原块菌)和Tubertexense(德州块菌,=T.lyoniiBtters(莱氏块菌)[4])为一枝。

图4 基于rDNA ITS序列的Neighbor-Joining系统发育树Fig.4 Neighbor-joining phylogenetic tree based on partial rDNA ITS sequences of strain a and b
3 讨 论
本研究中两菌株子囊果均呈黄褐色,表面无明显的疣状突起,子囊近球形至不规则形,不具明显的棱角。子实层内具有不育菌丝形成的褐色菌肉和乳白色、呈迷走状的产孢组织。其中,菌株a致密的产孢组织内具有呈蜂窝状分布的球形、棒状子囊,内含有典型的突起状纹饰的球型孢子,突起纹饰的高度约2 μm,孢子直径(含纹饰)约20 μm;菌株b子实层松散,有裂隙,产孢组织内不但具有蜂窝状分布的子囊和球型孢子,还具有似“口袋”的大子囊,其内含有大量的褐色、表面光滑、两端“芒果尖”状椭球型孢子。
与龙月娟等[11-12]描述的印度块菌形态结构略有不同,本研究中菌株子囊果横切面上的明暗相间的纹脉中的明脉呈亮色,可能是由于产孢组织内蜂窝状排布的子囊和子囊孢子腔室的存在而呈现亮色;不可育菌丝缠绕在产孢组织的外围构成褐色菌肉组织而呈现暗纹。同时,子囊果的包被构造显示,包被较薄,且外部无明显的疣状突起。这些特点又区别于黑孢块菌、夏块菌、冬块菌等块菌[3-4,6-7]。而在两菌株子实层内,蜂窝状排布的子囊以及其内具有的1~4个子囊孢子,这与龙月娟等[12]的研究结果相同;但是对于该球型孢子和呈“口袋”状子囊内的光滑型、两端“芒果尖”状的椭球形孢子,明显区别于中国已经报道的各块菌类群的子囊孢子[2-4,13-14]。然而,对于这种褐色、外壁光滑、两端芒果尖的椭球型孢子,有学者认为外壁光滑的孢子是败育的子囊孢子,具体原因目前还没有统一的说法[11-12]。
对系统进化树分析表明,菌株a和b聚为一枝,但与其它块菌属成员的相对支持率较低。结合形态解剖学的研究,推断菌株a和b可能是新发现类群。根据Gabriel[15]和Chen[16]的研究,从形态解剖学角度,笔者认为本研究描述的两菌株可能属于猪块菌属Choiromyces。原因是除了本研究中描述的菌体包被薄、菌体外无明显刺突、菌丝属于交织组织等特点与猪块菌属Choiromyces相一致外,本研究中带刺突纹饰的球状类型孢子与Choiromycesmagnusii电镜照片[15]中的孢子在大小、形态等方面极为相似。1990年,刘波等报道中国关于蜂窝孢猪块菌Choiromycesalevolatus的新记录[17]。所以进一步推断,本文中的菌株a和b可能是猪块菌属Choiromyces的中国新记录种。Choiromyces属是Tuberaceae(块菌科)的一个属,该属1831年被Vittadini首次描述[15]。有学者研究该属与Tuber属的亲缘关系比Terfezia地菇属近[18]。Kirk等[19]认为该属有5个种,但Gabriel[15]提到的有10种之多。猪块菌属Choiromyces在国外有很多别称,例如“猪块菌(猪的松露)”、“高加索块菌”、“高加索地区的松露”、“特兰西瓦尼亚的白松露”、“匈牙利松露”等[15]。
[1]王云.块菌的研究及我国块菌资源的开发利用 [J].食用菌, 1988, 3: 2-3.
WANG Y. Truffles and truffle resources development and utilization in our country [J].Ediblefungus, 1988, 3: 2-3.
[2]王云,刘培贵.块菌名实考证及其资源保护 [J].植物分类与资源学报,2011, 33(6): 625-642.
WANG Y, LIU P G. Verification of Chinese names of truffles and their conservation in natural habitats [J].PlantDiversityandResources, 2011, 33(6): 625-642.
[3]方明.15个块菌菌株的形态学与分子生物学鉴定 [D].长春:吉林农业大学, 2006.
[4]任德军.中国块菌属(Tuber)系统学研究 [D].福州: 福建农林大学, 2003.
[5]任德军,宋曼殳,姚一建.中国块菌属研究概况 [J].菌物研究, 2005, 3(4): 37-46.
REN D J,SONG M S,YAO Y J. Literature review ofTuberspecies in China [J].JournalofFungalResearch, 2005, 3(4): 37-46.
[6]陈娟,邓晓娟,陈吉岳,等.中国块菌属多样性 [J].菌物研究, 2011, 9(4): 252-253.
CHEN J, DENG X J, CHEN J Y,etal. A checklist of the genusTuber(Pezizales, Ascomycota) in China [J].JournalofFungalResearch, 2011, 9(4): 252-253.
[7]陈应龙,弓明钦.块菌资源多样性及其地理分布 [J].中国食用菌,2000, 19(5): 6-7.
CHEN Y L, GONG M Q. Truffles resource diversity and their geographical distribution [J].EdibleFungiofChina, 2000, 19(5): 6-7.
[8]李志刚,梁存柱,王炜,等.贺兰山植物区系的特有性 [J].内蒙古大学学报(自然科学版), 2012, 43(6): 630-631.
LI Z G,LIANG C Z, WANG W,etal. Plant endemism in the Helanshan Mountains [J].JournalofInnerMongoliaUniversityNaturalScienceEdition, 2012, 43(6): 630-631.
[9]刘小平.贺兰山植物群落的空间分异研究 [D]. 呼和浩特: 内蒙古大学, 2008.
[10]王兆锭,张鹏.贺兰山林区马麝的生活习性及保护措施 [J].内蒙古林业调查设计, 1997, 1(6): 19-22.
WANG Z D, ZHANG P. Life habits and the protective measures ofMoschuschrysogasterin forest region Helan Mountains[J].InnerMongoliaForestrySurveyDesign, 1997, 1(6): 19-22.
[11]龙月娟,李荣春.昆明地区印度块菌子囊果的解剖结构研究[J].西北植物学报, 2009, 29(2):269-274.
LONG Y J, LI R C. Research on anatomical structure ofTuberindicumascocarps of Kunming, Yunnan Province[J].ActaBot.Boreal.-Occident.Sin, 2009, 29(2): 269-274.
[12]龙月娟,陈严平,李荣春.印度块菌子囊果内部解剖结构的扫描电镜观察[J].西北植物学报,2011, 31(11): 2 222-2 225.
LONG Y J, CHEN Y P, LI R C. Anatomical structure ofTuberindicumascocarps with scanning electron microscope[J].ActaBot.Boreal.-Occident.Sin, 2011, 31(11): 2 222-2 225.
[13]张大成,王云.中国块菌及其生态研究 [J].中国食用菌, 1990, 9(2): 25-27.
ZHANG D C, WANG Y. Study on Chinese truffle and its ecology [J].EdibleFungiofChina, 1990, 9(2): 25-27.
[14]张介平,刘培贵.中华夏块菌及其生态学研究 [J].食用菌学报, 2015,22(1): 34-40.
ZHANG J P, LIU P G. Ecological features associated withTubersinoaestivum[J].JournalofEdibleFungi, 2015, 22(1): 34-40.
[15]GABRIEL M, PABLO A, JOSÉ L M. Phylogenetic affiliation ofChoiromycesmagnusiiandC.venosusTuberaceae Ascomycota from Spain[J].MycolProgress, 2012,(11):463-471.
[16]CHEN JUAN, SUN LI HUA, SU YUN, et al.Choiromyceshelanshanensissp. nov., a new species of subterranean truffle from China [J].Mycoscience, 2016, 57(4): 279-286.
[17]LIU B, TAO K, CHANG M C. New species and new records of hypogenous fungi from China Ⅲ [J].ActaMycologicaSinica, 1990, 9(1): 25-30.
[18]PERCUDANI R, TREVISI A, ZAMBONELLI A,etal. Molecular phylogeny of truffles (Pezizales: Terfeziaceae, Tuberaceae) derived from nuclear rDNA sequences analysis [J].Mol.Phylogenet.Evol., 1999, 13:169-180.
[19]KIRK PM, CANNON PF, MINTER DW, STALPERS JA. Ainsworth & Bisbs Dictionary of the Fungi [M], 10th ed. CABI Europe, UK. 2008.
(编辑:潘新社)
Anatomy and Molecular Biology Study of Truffles Discovered in Helan Mountain of Inner Mongolia
ZHAO Yanling1, WANG Haifeng2, WANG Zhaoding3, FAN Yongjun1*
(1 Department of Biological Sciences and Technology, Baotou Teacher’s College, Baotou, Inner Mongolia 014030, China; 2 The Food and Drug Engineering College, Baotou Light Industry Vocational Technical College, Baotou, Inner Mongolia 014035, China; 3 Inner Mongolia Helan Mountain National Nature Reserve Administration, Ala Shan, Inner Mongolia 750306, China)
The two samples of truffles collected in the Helan Mountain region of Inner Mongolia were studied by Anatomy and molecular biology. (1) Two strains of ascoma were tan, smooth surface, no obvious verrucous protuberance and edges. (2) Gleba of strain a is ivory, dense and vagus veins. Hyphae organization is brown; Spherical or rod-shaped ascus in honeycomb arrangement contained 1-4 ascospore with package; the surface ascospore had a protuberant ornamentation and was spherical. Thickness of double wall was about 1.7 um, diameter 20 microns (including ascospore grain). (3) Gleba of strain b had cracks, loose, in addition to the above ascus and spore, also possessed a special ascus like “pocket” containing a large number of sharp, smooth, brown ellipsoidal spores. (4) Molecular evolution analysis shows that the two strains into one, but there is a relatively low approval ratings for genusTuber. Therefore, the two strains were concluded that they might be a new record species of genusChoiromycesin China.
Helan Mountain; truffles; ascoma; ascus; ascospore; anatomical study; phylogenetic
1000-4025(2016)09-1781-06doi:10.7606/j.issn.1000-4025.2016.09.1781
2016-06-15;修改稿收到日期:2016-09-06
内蒙古自然科学基金(2014MS0302,2016MS0301 );国家自然科学基金(31260132,31460188 )
赵艳玲(1978-),女,博士,副教授,主要从事真菌资源利用研究。E-mail:nmzhaoyanling@163.com
樊永军,教授,主要从事菌根学研究。E-mail:fanyj1975@163.com
Q246; Q248; Q789
A
