河南伏牛山区天然红豆杉种群结构与动态研究
2016-10-28张志录刘中华陈明辉张巧莲杨风岭
张志录, 刘中华, 陈明辉, 张巧莲, 杨风岭
(1.平顶山学院, 河南 平顶山 467000; 2.许昌职业技术学院, 河南 许昌 461000)
河南伏牛山区天然红豆杉种群结构与动态研究
张志录1, 刘中华1, 陈明辉1, 张巧莲2, 杨风岭1
(1.平顶山学院, 河南 平顶山 467000; 2.许昌职业技术学院, 河南 许昌 461000)
采用聚集度指标和编制静态生命表的方法,对伏牛山区红豆杉种群的空间格局及其演化动态进行了研究,结果显示:(1) 伏牛山区红豆杉种群的空间分布格局总体上呈现聚集分布状态,聚集程度从幼苗阶段到老龄阶段的发育过程中,呈现先降后升的U型变化趋势;(2) 不同龄级的红豆杉树存活量差别较大,种群存活曲线属于Deevey-Ⅲ型;(3) 幼苗期和幼树期的红豆杉死亡率高,Ⅲ级(3 m高)以后死亡率明显下降,种群数量趋于稳定。本研究表明,伏牛山自然保护区天然红豆杉种群呈现明显的衰退趋势,严酷的环境筛致使红豆杉幼苗死亡率高可能是该种群衰退的主要原因,但树木资源缺乏保护而被滥伐或盗伐、生境被破坏等影响也不容忽视。施行严格的封育政策并辅以人工繁育措施,是保护和恢复该宝贵资源的当务之急。
濒危植物; 生命表; 种群结构; 红豆杉; 时间序列分析
红豆杉(Taxuschinensis)是第四世纪孑遗植物,是世界濒危植物,已在地球上生活250万年,被称为活化石。中国红豆杉是中国特有种,属国家一级保护植物,南北方地区均有分布。在华中地区主要分布于海拔1 000 m以上山地上部未被干扰的生境,在华南和西南地区多分布于1 500~3 000 m的山地落叶阔叶林中[1]。因受花期不遇、幼苗竞争力弱、种子捕食压力和种子萌发条件等的限制,天然红豆杉更新困难[2];自发现含有广谱、强活性的抗癌药物紫杉醇(Taxol)以来,红豆杉资源遭到严重破坏而数量急剧下降,目前已处于严重的濒危状态[3],亟待保护与恢复。
种群结构及空间分布格局是种群最基本的特征,既可反映种群内大小不同的个体的分布和配置状况,也能反映种群的数量动态及其未来发展与演变趋势,对于种群及群落的更新复壮、生态系统的恢复具有重要意义[4-8]。目前,有关红豆杉种群的研究报道,多见于南方红豆杉,集中在种群结构[9-11]、种群分布格局[12-13]和种间关系[14-15]以及天然更新与种子库[2,16]等方面。这些研究揭示了南方红豆杉种群结构及其动态的影响因素、形成机制,对于南方红豆杉濒危种群的保护发挥了重要作用。然而,红豆杉的分布存在着地域广泛性和生境异质性,目前,有关华中地区红豆杉种群结构与动态的研究比较缺乏。
伏牛山国家级自然保护区处于暖温带向亚热带的过渡区,是长江、黄河和淮河3大水系的分水岭,是河南省天然林分布最大的区域,区内动植物种类繁多,分布有红豆杉等多种珍稀植物种群[17]。目前,有关该区域红豆杉种群的空间分布格局与种群数量结构及动态研究尚未见报道。2012—2014年对伏牛山自然保护区天然红豆杉资源开展了专项调查,在此基础上,本文就濒危天然红豆杉种群的空间分布格局与种群动态进行分析,以期为该地区濒危植物红豆杉种群的保护与恢复提供科学依据。
1 材料与方法
1.1研究地概况
伏牛山国家自然保护区位于河南省西南部,地理坐标为东经110°30′—113°05′,北纬32°45′—34°00′。伏牛山是秦岭山脉向中原延伸的东端支脉,呈西北—东南走向,东西绵延约400 km,南北宽约40~70 km。海拔高度为500~2 200 m,平均海拔1 000 m。在我国地势的三大阶梯中,伏牛山正处在第二阶梯向第三阶梯过渡地带,山体完整高大,山峰突兀耸立,地形、土壤复杂多样;地理位置特殊,处在我国暖温带和北亚热带过渡区,也是华北、华中、西南植物的镶嵌地带,属暖温带落叶阔叶林向北亚热带常绿落叶混交林的过渡区。由于高差大,区内气候、土壤和自然植被的垂直带谱明显。伏牛山还是长江、黄河、淮河3大水系的分水岭和一些支流的发源地,为重要的水源涵养林区。区内气候温暖湿润,雨量充沛,年降水量800~1 100 mm,是著名的暴雨区;年最高气温26.5~28.5℃,最低气温-1.5~-2℃,年均温12.1~15.1℃。复杂多样的生态环境条件,区内生物资源多样性丰富,仅珍稀、濒危植物就有38科70种之多,中国特有属37个,国家重点保护野生植物32种,是我国中部重要的植物资源宝库,在研究植物区系、生态学等方面具有重要的科学价值[17]。
1.2样地设置与调查
于2012—2014年,采用样地调查法,对伏牛山自然保护区的天然红豆杉种群进行调查。其中尧山泉水沟村:红豆杉主要生长在纵长270 m山坡的中、上坡位,设置乔木调查样方9个;宝天曼石鼓村:红豆杉主要分布在纵长180 m山坡的下坡位,设置乔木调查样方15个;玉皇山毛河村(玉皇山1):红豆杉种群沿山谷呈狭长状分布,主要集中在纵长约340 m山坡的中、上坡位,设置乔木调查样方12个;柴家沟村(玉皇山2):红豆杉主要分布在中、下坡位,设置乔木调查样方14个。
乔木层每个样方面积为20 m×20 m,对样方内胸径2 cm以上的乔木进行每木调查,记录树名、株数、树高、胸径、冠幅、盖度等:用胸径围尺测量乔木胸径;用测高器测量树高;按南北和东西两个方向,用皮尺测量树冠在地面上的垂直投影的直径,作为冠幅。在乔木样方的四角和中心设置5个5 m×5 m的小样方进行灌木层调查,记录灌木种名、株数、高度、基径、冠幅、盖度等;用游标卡尺记录灌木的基径,用钢卷尺测量株高和冠幅。随机选取5个l m×1 m的小样方进行草本层调查,记录种名、株数、高度、盖度、多度、干鲜重等。同时,对每个样方的生境进行调查,记录经纬坐标、海拔高度、地形起伏、坡向、坡位、坡度、土壤类型、土层厚度及剖面等。
1.3数据处理
1.3.1龄级结构分析本文采用空间序列替代时间变化的方法,应用大小结构分析法来研究种群年龄结构特征[18-19]。刘佳庆等[7]根据红豆杉的生长规律和前人的研究结果,以树高(H)结构反映种群年龄结构。参考此法,将伏牛山国家自然保护区的红豆杉种群高度划分为6个级别,以此代替该区域红豆杉的年龄级(表1)。

表1 伏牛山国家自然保护区红豆杉种群高度结构分级
注:Ⅰ为红豆杉1级幼苗,包括Ⅰ1(1年生幼苗)和Ⅰ2(2~3年生幼苗);Ⅱ为红豆杉幼树;Ⅲ为红豆杉小树;Ⅳ为红豆杉壮树;Ⅴ为红豆杉大树;Ⅵ为红豆杉老树。
1.3.2种群空间分布格局分析采用聚集度指标,对天然红豆杉种群的空间分布格局进行分析。聚集度指标包括方差均值比(DI)、负二项参数(K)、丛生指数(I)、平均拥挤指数(m*)、聚块指数(Pai)和Green指数(GI),各指标计算公式及意义参考文献[7,20—22]。
1.3.3种群结构与动态分析采用陈晓德[23]的量化方法,对红豆杉的种群结构与动态进行分析。
(1) 种群内两相邻年龄级间个体数量变化动态(Vn):
(1)
式中:n——年龄级(高度级);Sn,Sn+1——第n,第n+1年龄级的个体数;Vn——种群个体数量的动态变化,其值为正值时表示种群内两相邻年龄级个体数量呈增长的动态趋势,负值则表示衰退,零值表示稳定。
(2) 种群的年龄结构动态量值(P)由各年龄级间动态Vn对年龄级个体数(Sn)进行加权分配而得:
(2)
式中:k——年龄级数量。
(3) 考虑种群年龄级数量(K)及种群年龄级个体数(Sn)因素的种群年龄结构动态(E):
(3)
1.3.4种群静态生命表和存活曲线采用静态生命表编制方法[24]编制红豆杉种群静态生命表,选用一次性移动平均法对红豆杉的种群龄级结构进行模拟和预测[25],公式如下:
(4)式中:n——需要预测的未来时间年限;Mt——未来n年时t龄级的种群大小;Xk——当前k龄级的种群大小。
2 结果与分析
2.1种群年龄-个体大小结构分析
由图1可知,尧山泉水沟村红豆杉种群共79棵,龄级结构不完整。幼苗数量少且全为根蘖苗,3年来的调查均未发现种子萌发苗;9 m以上的老龄树缺失。总体上种群年龄结构呈正态分布,为稳定型种群。但因幼苗的严重缺失,未来可能会发生衰退演替。

图1 伏牛山国家自然保护区红豆杉种群年龄结构
宝天曼石鼓村红豆杉更新种群653棵,龄级结构完整。有大量的更新幼苗,幼苗数占总数的67.69%;幼树数量严重不足,仅占总数的2.14%。因Ⅱ级幼树的严重不足和林龄偏大,未来将呈现衰退演替。
玉皇山毛河村(玉皇山1)红豆杉更新种群276棵,种群龄级结构完整,结构形态近钟形,呈现为龄级偏高的稳定型种群。其中Ⅲ龄级小树个体数量的比例突然减少,仅为7.61%,将严重影响种群的未来发展。
玉皇山柴家沟村(玉皇山2)红豆杉更新种群1 117棵,幼苗占总数的77.98%,老树比例为1.61%,种群结构呈现基部宽顶部窄的金字塔型,属增长型种群。
2.2种群空间分布格局
由表2可知,伏牛山各段的天然红豆杉种群中各龄级的DI值都大于1,均符合聚集分布的特征。其中,尧山种群的Ⅰ,Ⅴ龄级的K值分别为0.15,0.40,数值较小,呈现较高的集群分布特征;而Ⅴ龄级的GI值为0.01,高度接近0,又符合随机分布特征。宝天曼种群各龄级聚集度波动变化明显:第Ⅰ龄级高度聚集,第Ⅱ龄级有所下降,第Ⅲ龄级则有所提高,而第Ⅳ,Ⅴ龄级明显下降,第Ⅵ龄级又明显提高。玉皇山南坡种群各龄级亦呈现由幼龄级到中龄级聚集度下降而由中龄级至大龄级又有所增加的趋势;北坡种群从第Ⅰ龄级至第Ⅴ龄级均为集群分布,第Ⅵ龄级服从随机分布。

表2 种群空间分布格局分析
注:SD为空间分布;C为集群分布;R为随机分布;E为均匀分布;K为负二项参数;I为丛生指数;m*为平均拥挤指数;Pai为聚块性指数;DI为方差均值比;GI为Green指数;t为T-test中的t值,查t值表所得。
总体上看,由幼龄级至中龄级,聚集程度下降而由中龄级至大龄级聚集程度增强,聚集度随龄级增长而呈现U形动态变化。
2.3静态生命表和存活曲线
由表3可知,伏牛山自然保护区内红豆杉种群之间、种群内各龄级之间的消失率,有明显的差异。尧山境内的天然红豆杉种群,Ⅰ,Ⅱ龄级的消失率为负值,说明幼苗数量不足,该种群需要补充一定数量的幼苗来维持发展,否则将走向衰退;宝天曼种群在从Ⅰ龄级向Ⅱ龄级发展过程中的消失率极高,说明该龄级个体数量严重缺少,同时也说明该种群缺少种苗生长到幼苗的存活条件;玉皇山南坡种群中,Ⅲ,Ⅳ,Ⅴ龄级的消失率均为负值,说明种群个体数量波动明显,中、幼龄级个体数量较少;玉皇山北坡种群,各龄级个体消失率低,且随着龄级的升高而逐步减少,表明该种群更新状态较为理想。
由图2可以看出,伏牛山东段的尧山天然红豆杉种群,存活曲线属于Deevey-Ⅰ型,为衰退型的幼龄种群。伏牛山中部的宝天曼天然红豆杉种群,存活曲线在Ⅰ,Ⅱ龄级间斜率较大,说明幼苗在向幼树的生长过程中,死亡率很高;而在Ⅱ,Ⅲ,Ⅳ龄级间曲线平缓并逐级上升,而Ⅴ龄级向Ⅵ龄级生长的过程中再现较高的死亡率。总体来看,该种群存活曲线介于Deevey-Ⅰ,Deevey-Ⅱ型之间,但因Ⅱ龄级个体存活数量太少,未来将发生衰退演替。伏牛山西段的玉皇山内,毛河村(玉皇山1)天然红豆杉种群的存活曲线属于Deevey-Ⅱ型,而柴家沟(玉皇山2)种群的存活曲线属于典型的Deevey-Ⅲ型,有良好的更新状态。伏牛山3个不同地段的天然红豆杉种群,存活曲线类型不同,表明其未来的发展态势不同,东段种群将发生衰退演替,中段种群在未来一段时间内呈现稳定状态,而西段种群将呈现增长态势。分析原因,除种群或个体所处微环境的差异外,主要是伏牛山东段是低山丘陵区,人为活动较为频繁,天然种群受之影响明显。

表3 伏牛山区红豆杉种群静态生命表

图2伏牛山红豆杉种群存活曲线
2.4种群动态
从表4可以看出,尧山种群中第Ⅰ,Ⅱ龄级的Vn为负值,为衰退型结构,Ⅲ,Ⅳ级为增长型结构;宝天曼种群中,Ⅱ,Ⅲ,Ⅳ龄级为衰退型结构;Ⅰ,Ⅴ龄级为增长型结构。玉皇山南坡种群中,Ⅲ,Ⅳ,Ⅴ龄级为衰退型结构;玉皇山北坡种群各龄级均为增长型结构。从种群整体的P,E来看,所有种群均为正值,表明各个种群皆为增长型种群。其中玉皇山南坡种群E值0.001 4接近于0,说明该种群为稳定种群。
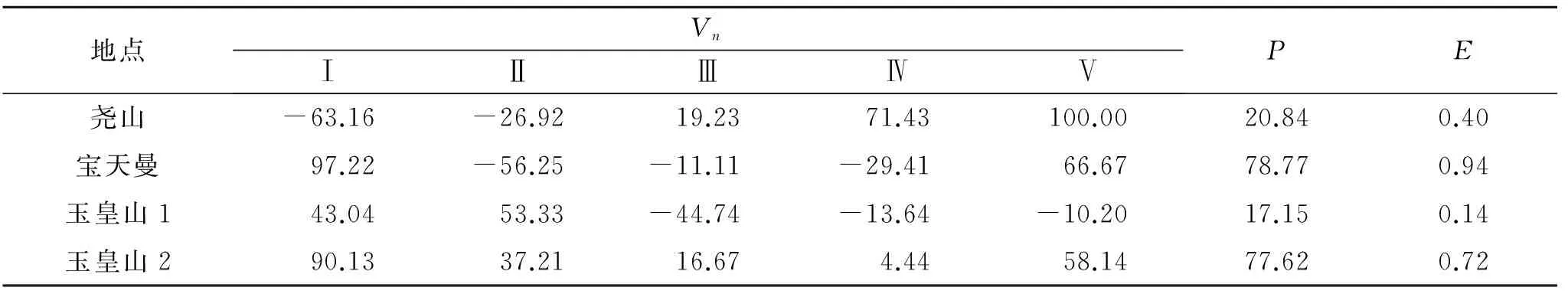
表4 伏牛山红豆杉种群动态分析
由表5可以看出,不同生境的种群,动态变化规律有明显差异。在未来2个龄级时间后,尧山红豆杉种群Ⅱ,Ⅲ龄级的个体数量减少,Ⅳ,Ⅴ,Ⅵ龄级个体数量有少量增加;宝天曼红豆杉种群Ⅱ龄级个体数量剧增,Ⅲ,Ⅳ,Ⅴ龄级个体数量均有所减少;玉皇山南坡种群Ⅱ,Ⅲ龄级个体数量微增,而Ⅳ,Ⅴ,Ⅵ龄级均微降;玉皇山北坡种群Ⅱ—Ⅵ龄级的个体数量均有所增加,Ⅱ龄级剧增。经过未来4个龄级时间后,尧山种群中Ⅳ龄级个体数量减少,而Ⅴ,Ⅵ龄级个体数量均呈现小幅度增加趋势;宝天曼种群的Ⅵ龄级个体数量剧增,而Ⅴ龄级个体数量有所减少,Ⅵ龄级个体数量保持稳定;玉皇山南坡种群Ⅳ龄级个体数量增加,而Ⅴ,Ⅵ龄级个体数量均减少;玉皇山北坡种群Ⅳ,Ⅴ,Ⅵ龄级个体数量均增加。经过6个龄级时间后,各种群Ⅵ龄级个体数量,除了尧山种群稳定在上个时间外,其余3个地方的种群Ⅵ龄级个体数量均呈不同程度的增加。

表5 伏牛山红豆杉种群动态时间序列分析
3 结论与讨论
伏牛山东段的尧山红豆杉种群和西段的玉皇山南坡种群存活曲线为Deevey-Ⅰ型,为衰退型种群;中段的宝天曼种群和西段的玉皇山北坡种群的存活曲线为Deevey-Ⅲ型,属于增长型种群。在未来的2,4,6个龄级时间后,尧山种群和玉皇山南坡种群,由于幼苗数量不足,小树数量呈现衰减趋势;宝天曼种群的Ⅵ龄级个体数量呈现增长趋势,表明该种群呈现增长动态;玉皇山北坡种群各龄级稳定增长,呈现良好的更新状态。
种群动态反映了植物种群的未来命运。从种群的静态生命表和存活曲线来看,在伏牛山红豆杉种群的早期(如幼苗期、幼树期)死亡率高,而中龄期以后则种群个体数量稳定。这说明红豆杉幼苗和幼树的生长过程中,存在严格的环境筛选,多数个体难以通过环境筛选而死亡。
处于伏牛山西段深山处的柴家沟村红豆杉种群结构完整,而处于东段浅山区的泉水沟村的红豆杉种群龄级结构不完整,种群的“大小级结构金字塔”并不呈现为三角形状,这可能与结构龄级的划分幅度有关,抑或是严酷的环境筛的作用结果。此外,频繁的人为活动,干扰或破坏了红豆杉的生境,致使红豆杉数量减少,可能是造成某些“大小级结构”缺失的原因。
种群分布格局是物种与环境长期相互作用的结果。伏牛山各段的天然红豆杉种群,均呈现聚集分布状态;不同发育阶段的种群,聚集程度不同,这与王磊等[13]的研究结果相一致。伏牛山自然保护区天然红豆杉种群的空间分布格局呈现聚集分布,聚集程度随龄级变化而呈U形动态变化,这种动态变化,客观地反映了红豆杉种群在生长发育过程中的生态策略和适应机制[22],因种子散落和母株发生萌蘖相对集中,导致幼苗期和幼树期的红豆杉呈现高度聚集分布;在后来的成长过程中,因个体所处的微环境不同,以及个体间对空间和养分的激烈竞争,导致幼苗和幼树个体大批死亡而呈现聚集度有所下降。大树以后阶段的聚集度不降反升的生态机理,有待今后进一步研究。
特定时间生命表反映多个世代重叠的年龄动态历程中的一个特定时间断面,它是用同一时期收集到的种群所有个体的年龄数据编制而成,而不是对同生群的全部生活史追踪。由于红豆杉种群为天然林,研究方法是由“空间代时间”,所以调查所得的数据并不完全满足编表的3个假设,在编制的生命表中会出现死亡率为负值的情况。对此,张文辉等[26]认为,这不仅客观反映了种群波动真实情况,而且对种群数量预测有重要生态意义。天然植物种群各龄级株数波动是普遍现象,本文并未对数据作数据匀滑处理,似乎更有利于反映种群现实状况。
由于红豆杉属国家一级重点保护植物,种群资源稀少,不能作伐木解析,而且用生长锥钻木测年的方法也会对树体造成一定的破坏。因此,本文采用了依据高度划分龄级的方法。但生命表中年龄的划分是以年为参照等距划分的,依个体大小划分时,年龄间距与个体大小等级并不是线形的。这样编制生命表和绘制存活曲线,依据其线形判断种群动态时,有时可能会有误判。
本研究表明,伏牛山国家自然保护区红豆杉种群前景堪忧,必须采取严格的封育措施对其和生境加以保护,严禁任何形式的破坏;建议辅以人工繁育措施,促进种群的更新与壮大。
[1]中国科学院植物研究所.中国高等植物图鉴(第1册)[M].北京:科学出版社,1972.
[2]邓青姗,朱琼琼,鲁长虎.南方红豆杉的天然更新格局及食果鸟类对其种子的传播[J].生态学杂志,2008,27(5):712-717.
[3]臧敏,卞新民.江西三清山珍稀濒危植物考察研究[J].武汉植物学研究,2003,21(6):515-520.
[4]Armesto J J, Casassa I, Dollenz O. Age structure and dynamics of Patagonian beech forests in Torres del Paine National Park, Chile[J]. Vegetatio,1992,98(1):13-22.
[5]Fuchs M A, Krannitz P G, Harestad A S. Factors affecting emergence and first-year survival of seedlings of Garry oaks (Quercusgarryana) in British Columbia, Canada[J]. Forest Ecology & Management,2000,137(1):209-219.
[6]刘普幸.疏勒河中下游绿洲胡杨种群结构与动态研究[J].自然资源学报,2011,26(3):429-439.
[7]刘佳庆,李宁,熊天石,等.濒危植物南方红豆杉不同种群的结构和动态变化[J].热带亚热带植物学报,2014,22(5):479-485.
[8]张亚芳,李登武,王梅,等.黄土高原不同地区杜松种群结构与动态[J].林业科学,2015,51(2):1-10.
[9]李先琨,向悟生,苏宗明.南方红豆杉无性系种群结构和动态研究[J].应用生态学报,2004,15(2):177-180.
[10]李先琨.濒危植物元宝山冷杉与南方红豆杉种群生态学研究[M].北京:科学出版社,2006.
[11]李帅锋,刘万德,苏建荣,等.云南兰坪云南红豆杉种群年龄结构与空间分布格局分析[J].西北植物学报,2013,33(4):792-799.
[12]李伟,王瑞雪,张光富,等.南方红豆杉迁地保护种群的点格局分析[J].生态学杂志,2014,33(1):16-22.
[13]王磊,孙启武,郝朝运,等.皖南山区南方红豆杉种群不同龄级立木的点格局分析[J].应用生态学报,2010,21(2):272-278.
[14]茹文明,铁军,张桂萍,等.南方红豆杉群落种间关系的数量分析[J].山西大学学报:自然科学版,2010,33(4):609-614.
[15]唐愽言,操国兴,蒋万平.武陵山大木山自然保护区红豆杉群落优势乔木种群生态位和种间联结[J].植物分类与资源学报,2014,36(5):668-674.
[16]岳红娟,仝川,朱锦懋,等.濒危植物南方红豆杉种子雨和土壤种子库特征[J].生态学报,2010,30(16):4389-4400.
[17]王理顺,申洁梅,李华堂,等.伏牛山自然保护区珍稀濒危植物资源现状与保护对策[J].河南林业科技,2003,23(3):42-44.
[18]Frost I, Rydin H. Spatial pattern and size distribution of the animal-dispersed treeQuercusroburin two spruce-dominated forests[J]. Ecoscience,2000,7(1):38-44.
[19]李先琨,苏宗明,向悟生,等.濒危植物元宝山冷杉种群结构与分布格局[J].生态学报,2002,22(12):2246-2253.
[20]张金屯.植物种群空间分布的点格局分析[J].植物生态学报,1998,22(4):344-349.
[21]项小燕,吴甘霖,段仁燕,等.大别山五针松种内和种间竞争强度[J].生态学报,2015,35(2):389-395.
[22]李先琨,向悟生,欧祖兰,等.濒危植物南方红豆杉种群克隆生长空间格局与动态[J].云南植物研究,2003,25(6):625-632.
[23]陈晓德.植物种群与群落结构动态量化分析方法研究[J].生态学报,1998,18(2):214-217.
[24]吴明开,沈志君,刘海,等.梵净山自然保护区珙桐天然种群生命表与生存分析[J].生态学杂志,2012,31(6):1419-1424.
[25]韩路,王家强,王海珍,等.塔里木河上游胡杨种群结构与动态[J].生态学报,2014,34(16):4640-4651.
[26]张文辉,郭连金,刘国彬.黄土丘陵区不同生境沙棘种群数量动态分析[J].西北植物学报,2005,25(4):641-647.
Research on the Population Structure and Dynamics of the NaturalTaxuschinensisin Funiushan Mountain Area of He′nan Province
ZHANG Zhilu1, LIU Zhonghua1, CHEN Minghui1, ZHANG Qiaolian2, YANG Fengling1
(1.PingdingshanUniversity,Pingdingshan,He′nan467000,China;2.XuchangVocationalTechnicalCollege,Xuchang,He′nan461000,China)
Based on the aggregation indices and time-specific life table, the structure and dynamics of naturalTaxuschinensispopulation were studied. The results showed that: (1) the naturalTaxuschinensisresources distributed sporadically, but they relatively concentrated in the distribution area, and the aggregation class decreased first and then seemed to follow the U-shaped curve in the process of seedling to old trees; (2) the differences of survival amount were significant among the age classes, the survival curve followed the Deevey-Ⅲ type;(3) the mortality was higher at the stage of seedling and sapling, but it declined significantly after Ⅲ level (3 m). These results indicated that the trends of naturalTaxuschinensispopulation were a recession in Funiushan Mountain, high mortality of seedlings caused by harsh environmental sieve might be the main reason for the decline of population, meanwhile lack of resources protection induced the illegal felling trees, habitat destruction and so on also could not be ignored. So it is urgent to take measures to protect and restore the precious resources and to take the strict closure policy and auxiliary measures of artificial breeding.
endangered plant; life table; population structure;Taxuschinensis; time series analysis
2015-11-16
2015-12-20
河南省科技攻关计划项目“中国中部伏牛山特有红豆杉生态适应性与繁育关键技术研究”(122102310652);河南省环境科学重点学科(Pxy-zdxk-2013003);平顶山学院高层次人才科研启动经费(2012005)
张志录(1967—),男,河南上蔡人,博士,副教授,主要从事种群生态学和园艺学研究。E-mail:276996899@qq.com
杨风岭(1963—),男,河南许昌人,博士,教授,主要从事环境科学和植物化学研究。E-mail:13903900069@163.com
S718.5
A
1005-3409(2016)03-0262-07
