1961—2010年内蒙古草原植被分布和生产力变化——基于MaxEnt模型和综合模型的模拟分析
2016-10-24叶永昌周广胜殷晓洁
叶永昌,周广胜,殷晓洁
1 中国科学院植物研究所 植被与环境变化国家重点实验室,北京 100093 2 中国气象科学研究院,北京 100081 3 中国科学院大学,北京 100049
1961—2010年内蒙古草原植被分布和生产力变化
——基于MaxEnt模型和综合模型的模拟分析
叶永昌1,3,周广胜1,2,*,殷晓洁1,3
1 中国科学院植物研究所植被与环境变化国家重点实验室,北京100093 2 中国气象科学研究院,北京100081 3 中国科学院大学,北京100049
定量评估气候变化对内蒙古草原植被分布及其净第一性生产力的影响有助于理解干旱区域生态系统结构和功能对气候变化的响应。基于最大熵模型(MaxEnt)评价了气候因子的重要性,进而模拟了1961—2010年内蒙古草原植被的地理分布,同时应用综合模型模拟了净第一性生产力变化。研究表明,湿润指数(MI)、年降水量(P)、最暖月平均温度(Tw)和最冷月平均温度(Tc)是决定草原植被分布的主导气候因子。1961—2010年内蒙古草甸草原、典型草原和荒漠草原分布面积分别减少了5%、1%和62%,草原面积整体减少了11%,预示着草原向着荒漠化的方向发展。降水是决定内蒙古草原净第一性生产力变化的最重要因素。
内蒙古草原;MaxEnt模型;植被地理分布;综合模型;净第一性生产力
CO2等温室气体浓度增加使得全球温度持续上升,降水格局发生变化[1],严重影响了陆地生态系统的结构与功能[2- 6]。研究表明,受气候变化的影响,近年来内蒙古草原盐渍化、荒漠化严重[7- 9],退化面积达40%[10],生产力下降30%—50%[11],沙尘暴频发[12],导致了严重的生态问题与经济损失[13- 14]。
气候变化对内蒙古草原植被的影响主要体现在其分布区域及其生产力的变化方面。植被生态学认为,主要的植被类型表现着植被对主要气候类型的适应,每个气候分区都有一套相应的植被类型[15]。在植被地理分布模拟中,气候相关模型被广泛用于植被分布的模拟[16]。Phillips等[17]以最大熵原理为基础,构建了模拟物种分布的最大熵模型(MaxEnt)。MaxEnt模型具有良好的模拟效果和友好的使用界面,已经广泛地用于物种分布的预测[18- 22]。植被净第一性生产力(NPP)是连接生物圈与气候系统的桥梁[23],衡量了生态系统固定太阳能的速率,即能进入食物链的能量[24]。常用于模拟NPP的气候相关模型有Miami模型[25]、Thornthwaite模型[25]、Chikugo模型[26]和综合模型[27]。其中,综合模型以与植被光合作用密切相关的实际蒸散为基础,综合考虑了诸因子的相互作用,模拟效果最好,特别是对干旱半干旱地区。该模型已被广泛应用于植被净第一性生产力的模拟[7, 28- 31]。
尽管大量研究指出当前荒漠化在加剧,但对历史植被覆盖和植被地理分布格局知之甚少[32],通过模型模拟现有植被分布有利于回答区域尺度上的生态问题[33]。本研究试图基于MaxEnt模型和综合模型模拟1961—2010 年内蒙古草原植被地理分布和NPP的时空变化,以期为进一步预测未来气候变化对植被的潜在影响提供方法参考。
1 资料和方法
1.1内蒙古草原植被样本数据
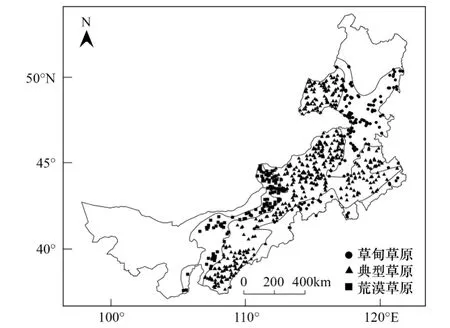
图1 内蒙古3种植被类型样本点的地理分布 Fig.1 Geographical distribution of samplings of three vegetation types in Inner Mongolia
植被分布的样本数据利用ArcGIS平台中的ArcMap从1∶100万中国植被图中提取。1∶100万中国植被图来源于《1∶1000000中国植被图集》的数字化,该图集由中国植被图编辑委员会编纂,于2001年由科学出版社出版。资料主要依据1949年以来全国各地进行的植被调查研究成果,特别是在20世纪80年代进行的大量的实地调查,能够真实地反映1961—1990年中国植被的分布状况。首先,将内蒙古地区提取出来,再提取所需的植被类型,然后通过创建随机点进行随机取样。已有研究表明,当样本量达到120个之后,MaxEnt模型的模拟精度趋于稳定[34]。为此,以120个样本点为基数,依据不同类型植被的面积比例关系,将样本数量设定为草甸草原135,典型草原470,荒漠草原120 (图1)。
1.2气象数据
气象数据来自中国气象科学数据共享服务网提供的1961—2010 年的内蒙古基本、基准地面气象观测站的日值数据集,包括:站点经伟度、日降水量、日平均气温、相对湿度、气压等要素。采用Thornton等[35]给出的截断高斯滤波算子空间插值方法,与数字高程模型 (Digital Elevation Model, DEM)数据结合,将气候数据插值成10km × 10km分辨率的空间栅格数据,并利用Thornton和Running[36]提出的方法得到日值空间格点的太阳辐射数据。基于气候标准年考虑,按照1961—1990年、1966—1995年、1971—2000年、1976—2005年、1981—2010年建立气候标准年数据库。
1.3气候因子的选取
影响植被地理分布的主要因子有3类:植被的耐寒性;完成生活史所需的生长季长度和热量供应;用于植被冠层形成和维持的水分供应[37]。翁恩生和周广胜[38]详细阐述了用于BIOME模型的气候因子在中国植被分类中的适用性,参考其研究成果,本研究选取最冷月平均温度(Tc,1月平均温度)反映植被的耐寒性,用最暖月平均温度(Tm,7月平均温度)和大于5℃积温(GDD5) 共同反映热量需求,采用年降水量(P)和湿润指数(MI=P/PET,年降水量和年潜在蒸散量的比值)表示水分需求,选取气温年较差(Td)反映气温变幅。年潜在蒸散(PET)采用Thornthwaite方法计算得来[39]。
1.4MaxEnt模型
MaxEnt模型基于贝叶斯定理,利用Gibbs 分布族将特征集进行加权并作为参数,进行一系列运算得到物种分布的最大熵联合分布估计而建立[40]。为了避免偶然误差,将模型迭代次数设定为10次,其它参数不变。首先模拟1961—1990年草原植被的分布,基于模型输出的各因子百分贡献率,筛选贡献更高的因子。以筛选的气候因子作为主导气候因子重新建模,模拟1961—1990年内蒙古草原植被的分布,并对模拟效果进行验证。然后以1961—1990年为基准,预测1966—1995、1971—2000、1976—2005、1981—2010年4个时期内草原植被的分布。MaxEnt模型默认输出为logistic形式,表示某种植被在整个模拟区域每个栅格上的存在概率(P),取值范围为0—1。根据统计学上小概率事件不可能发生的原理,当P<0.1 时,认为该类型植被不可能存在;当P≥0.1 时,认为该类型植被可以在此格点分布。由于不同类型植被的分布区域存在重叠,因此,规定草原植被的优先等级:草甸草原>典型草原>荒漠草原。当某个栅格被同时划分为不同类型的植被时,以优先等级的植被作为该栅格的植被类型。
1.5综合模型
周广胜和张新时[27]根据植物生理生态学特点及联系能量平衡方程和水量平衡方程的区域蒸散模式,利用Efimova在国际生物圈计划(International Biological Program, IBP)期间获得的世界各地的23组森林、草地及荒漠等自然植被的生产力资料及相应的气候资料,建立了联系植物生理生态学特点和水热平衡关系的植物的净第一性生产力模型,即综合模型,该模型的计算方法如下:
Rn=RDI×r×L×2.38×10-4
RDI=0.629+0.237PER-0.00313PER2
PER=PET/r
PET=58.93BT
BT=∑t/365 (0
式中,NPP表示净第一性生产力(g干重m-2a-1),Rn表示陆地表面所获得的净辐射量(J m-2a-1),r表示年均降水量(mm),L表示潜热蒸散(2503J/g),RDI表示辐射干燥度,PER表示年均潜在蒸散与年均降水的比值,BT表示年均生物温度(℃),t表示日均温(℃)。
2 结果分析
2.1主导气候因子筛选及其年际动态
首先采用MaxEnt模拟1961—1990年内蒙古草原植被的地理分布,通过模型自身的百分贡献率模块对气候因子的重要性进行评价,由于同一因子在3类草原植被中的百分贡献率不完全一致,因此,以某种因子在3种草原植被中百分贡献率的平均值作为衡量其重要性的标准。不同气候因子重要性排序为:湿润指数(MI)>年降水(P)>最暖月平均温度(Tw)>最冷月平均温度(Tc)>气温年较差(Td)>5℃积温(GGD5)。其中前4个因子的贡献率分别为46%、32%、9%和6%,总和达到了94%,能够充分解释分布结果。因此,湿润指数、年降水量、最暖月平均温度和最冷月平均温度是决定内蒙古草原植被地理分布的主导气候因子。
内蒙古草原植被主导气候因子动态的最优拟合曲线清晰地展现了气候的变化趋势(图 2)。结果表明,湿润指数和年降水量呈现出一致的三段式变化:1961—1970年呈下降趋势,1971—1990年为上升趋势,1991—2010年呈下降趋势。最暖月平均温度则与湿润指数和年降水量呈现出完全相反的变化趋势。最冷月平均温度在小幅波动中呈现上升趋势。总的来说,伴随着温度的升高和降水的减少,尤其是1991—2010年,内蒙古的气候呈现暖干化的趋势。
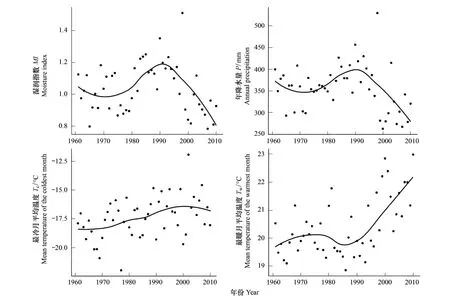
图2 1961—2010年内蒙古草原植被主导气候因子动态Fig.2 Dynamics of dominant climatic factors of steppe vegetation in Inner Mongolia during1961 to 2010
2.2MaxEnt模型的验证
将筛选的4个主导气候因子输入MaxEnt模型,再次模拟1961—1990年内蒙古草原植被的地理分布,然后采用ROC(Receiver operating characteristic curve)曲线下的面积AUC (Area Under ROC Curve) 值和Kappa值检验MaxEnt模型模拟的准确性。AUC值和Kappa值的评估标准见表 1。将模型迭代次数设定为10次,模型的验证方式选择交叉验证,即随机地将所有数据等分为10份,每次以90%的样本数据作为训练子集,以剩余的10%数据作为验证子集。训练子集用于训练模型,获取模型相关参数,构建草原植被与气候因子的最大熵模型。验证子集用于模拟准确度的验证,且验证子集每次都不重复,最终所有数据都将参与验证。基于验证子集得到的草甸草原、典型草原和荒漠草原的平均AUC值分别为0.90、0.85和0.95,表明MaxEnt模型的模拟准确性达到了非常好和极好的水平。
AUC值是以样本点的形式对模型进行验证,而Kappa值可以从整体上检验模拟的准确度。将模拟的1961—1990年内蒙古草原潜在植被分布与植被图对应的区划图进行Kappa一致性检验。结果表明,草甸草原、典型草原、荒漠草原的Kappa值分别为:0.95、0.70、0.57,均大于0.55。整个内蒙古草原植被的Kappa一致性值为0.76,总体达到了非常好的模拟效果,表明MaxEnt模型能够很好地模拟内蒙古草原植被的地理分布。
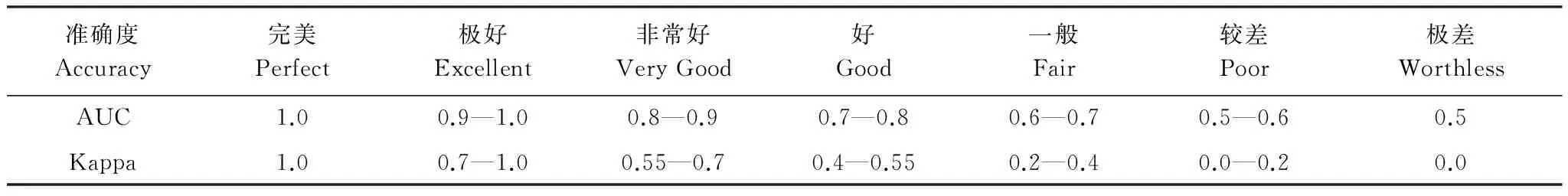
表1 AUC值和Kappa值及其与模型准确性的关系
2.3内蒙古草原植被地理分布变化
基于MaxEnt模型模拟结果可以得到1961—1990、1966—1995、1971—2000、1976—2005、1981—2010年5个时期的内蒙古草原植被地理分布及其面积变化 (图3和表 2)。5个时期3种草原植被均有分布,从东北向西南依次为草甸草原、典型草原和荒漠草原。其中,草甸草原主要分布于两个区域,典型草原主要分布在3个区域,荒漠草原在西北方向上分布较为离散。
草甸草原1966—1995年分布面积相比1961—1990年上升了4%,之后两个时期保持平稳,到了1981—2010年,面积又减少了11%,低于1961—1990年的分布水平。典型草原分布面积在1966—1995、1971—2000、1976—2005年3个时期呈减小趋势,在1976—2005年分布面积最小,相比于1961—1990年减少了12%, 1981—2010年面积开始回升,相比于1976—2005年面积增加了10%,接近于1961—1990年水平。荒漠草原面积在5个时期内呈递减趋势,1966—1995、1971—2000、1976—2005、1981—2010年相对于其前一时期面积分别减少了16%、22%、21%和26%。50年间3种草原植被整体分布面积呈缩小趋势,整个草原分布面积减少了11%。其中,草甸草原面积减少了5%,典型草原面积减少了1%,荒漠草原面积减少了62%。草甸草原向典型草原转化的面积(2.38×104km2)小于后者向前者转化的面积(3.57×104km2),荒漠草原转化为典型草原的面积占荒漠草原总面积的15%。
内蒙古草原植被5个时期分布重心的迁移距离和方向清晰地展现了草原植被对气候变化的响应 (图4和表3)。结果表明,草甸草原和典型草原整体向西南方向迁移,荒漠草原初期向西南方向迁移,随后逐渐转向东北方向。相对于1961—1990年,草甸草原在1976—2005年迁移距离最远(51km),典型草原在1971—2000年迁移距离最远(74km),荒漠草原在1981—2010年迁移距离最远(73km)。草甸草原、典型草原、荒漠草原迁移较为温和的年份分别是1966—1995年(24km)、1981—2010年(30km)、1966—1995(30km)。

图3 1961—2010年内蒙古草原植被地理分布Fig.3 Distribution of steppe vegetation in Inner Mongolia during 1961 to 2010
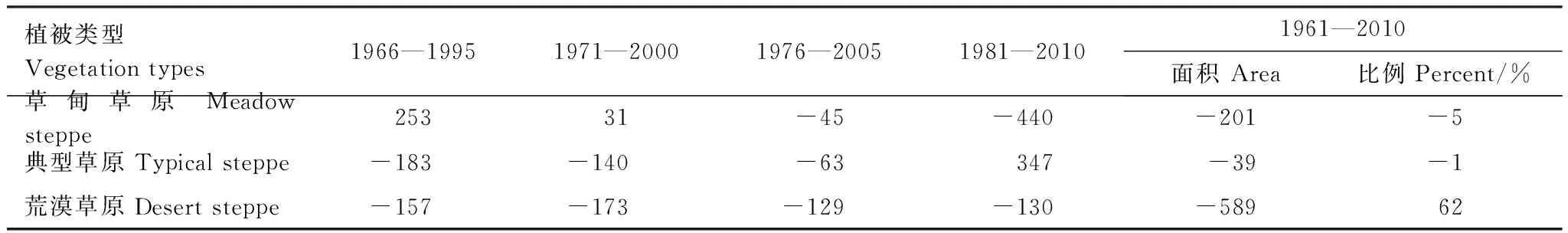
植被类型Vegetationtypes1966—19951971—20001976—20051981—20101961—2010面积Area比例Percent/%草甸草原Meadowsteppe25331-45-440-201-5典型草原Typicalsteppe-183-140-63347-39-1荒漠草原Desertsteppe-157-173-129-130-58962
-表示面积减少
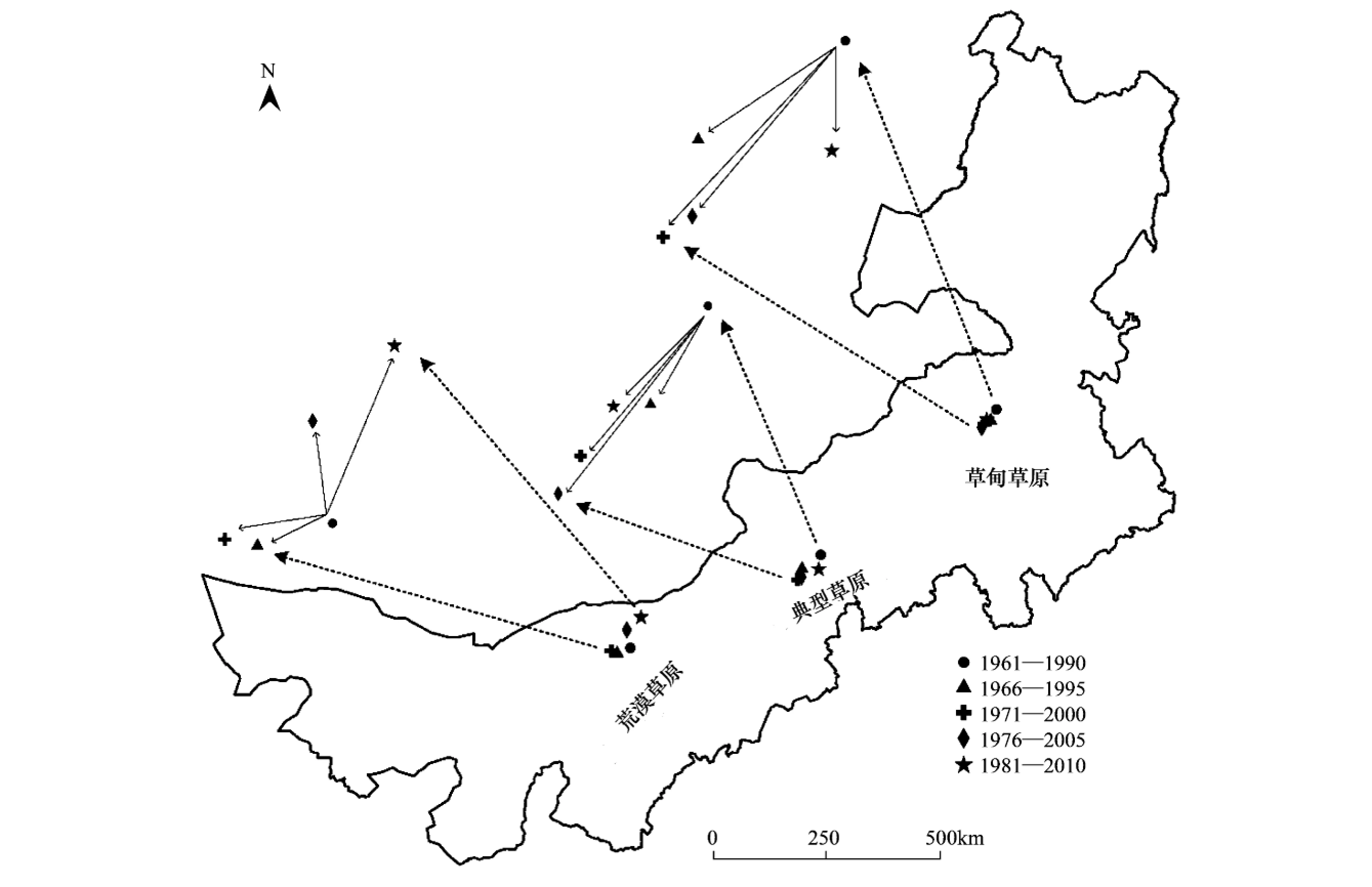
图4 1961—2010年内蒙古草原植被分布的重心变化Fig.4 Changes of gravity centers of steppe vegetation distribution in Inner Mongolia during 1961 to 2010

植被类型Vegetationtypes1966—19951971—20001976—20051981—2010距离/kmDistance方向Direction距离/kmDistance方向Direction距离/kmDistance方向Direction距离/kmDistance方向Direction草甸草原Meadowsteppe24西南42西南51西南29西南典型草原Typicalsteppe49西南74西南64西南30西南荒漠草原Desertsteppe30西南42南方40北方73东北
距离和方向的变化都是相对于1961—1990年
2.4内蒙古草原植被净第一性生产力动态
综合模型的模拟结果清晰地展现了内蒙古草原NPP在1961—2010年的变化趋势 (图5)。结果表明,内蒙古整个草原NPP的变化主要表现为3段不同的特征。1961—1970年呈下降趋势,1971—1990年转为上升趋势,1991—2010年则呈现下降趋势,这3个阶段NPP的年均值分别为 297、313、329g干重 m-2a-1。其中,草甸草原和典型草原的NPP与整个草原的变化趋势相同,而荒漠草原的NPP变化趋势相对平稳。1961—2010年内蒙古整个草原、草甸草原、典型草原和荒漠草原的NPP的年均值分别为:312、341、309、260 g干重m-2a-1。
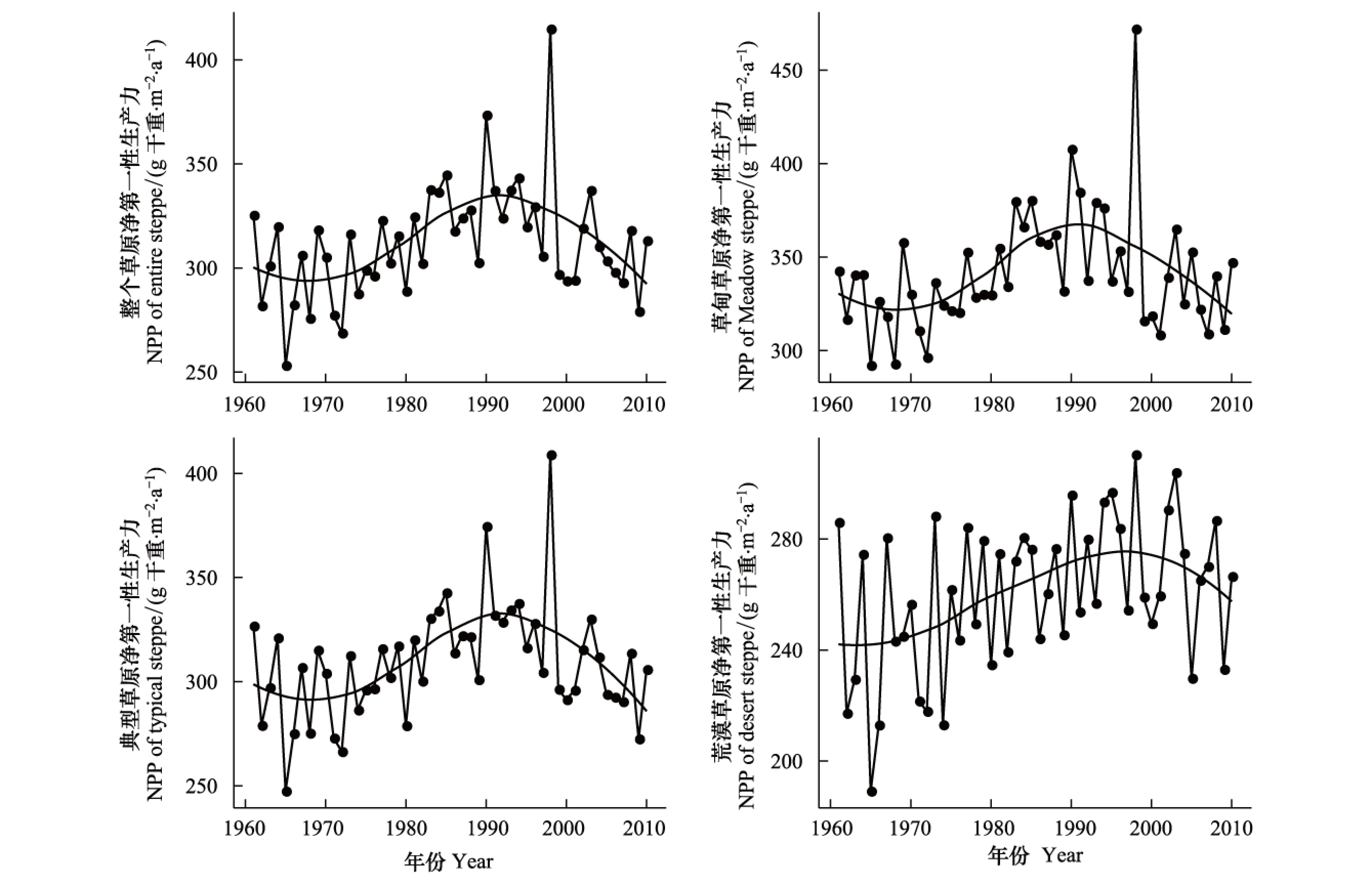
图5 1961—2010年内蒙古草原植被净第一性生产力动态Fig.5 Dynamic of NPP in Inner Mongolia during 1961 to 2010
3 讨论
尽管内蒙古草原植被的退化现状不能完全归因于气候变化,但气候变化毫无疑问加速了这一过程,并改变着草原植被的分布格局[7]。内蒙古草原处于干旱、半干旱区域,相对于其它自然生态系统,其结构和功能对气候变化最为敏感。研究基于MaxEnt模型和综合模型分析了1961—2010年内蒙古草原植被的分布格局和净第一性生产力变化, 研究结果对于理解气候变化对干旱区域草原植被的影响有重要作用。
净第一性生产力代表着从空气中进入植被的纯碳量, 是表征植被活动和陆地生态系统过程的关键参数, 对于理解地表碳循环过程具有重要的意义[41]。NPP的影响因素有很多,包括降水、营养物质、土壤属性,以及一系列生物间相互作用[42]。通常认为水分是影响内蒙古草原NPP最重要的因素, NPP和年降水与季节性降水都有显著的相关性[43- 46]。本研究表明,NPP和温度之间没有显著的相关性,降水(R2=0.68,P<0.01)是决定NPP变化的最主要因素,与已有研究结果相似。尽管本世纪初开展了退牧还草的生态工程建设,使原有的退化草地得到一定恢复,然而这些研究通常是基于样点的围封观测,难以排除气候因子的干扰[46]。本研究通过综合模型模拟了内蒙古草原植被在历史气候条件下潜在生产力的动态,表明区域降水的波动是决定植被潜在生产力的最主要因素。因此,发展草原节水和保水技术是维持草原生态系统功能稳定的关键。
MaxEnt模型本质上是生物气候包络模型(Bioclimatic Envelope Models,BEMs)的一种,BEMs利用具有地理参考的环境变量和物种分布信息之间的相关性推断物种的生态需求,模拟物种的地理分布[47],目前已被广泛应用于解决保护生物学、生物地理学、生态学和分类学中的问题[48]。但这些应用多数是在物种尺度上,因为BEMs构建的基础是生态位理论,将物种对环境的适应能力看作是环境变量的函数[17],假设物种当前分布范围的气候条件代表着物种所需的气候条件,所构建的生物气候包络代表着物种的实际生态位[49]。但这并不妨碍学者对这种模型进行全新的应用,用BEMs研究植物功能型(PFTs)和群落集合(相对于个体而言)的分布范围[50]。Pau等[51]采用Maxent模型对比了夏威夷群岛C3和C4植物功能型及其相关的物种集合(将152个物种划分为6类)的分布范围。南非国家生物多样性研究所(South African National Biodiversity Institute, SANBI)在评估南非生物群区对气候变化的脆弱性时就采用Maxent来模拟现在和未来生物群区的分布范围[52]。Werneck等[53]基于MaxEnt构建了生物群区分布模型(Biome Distribution Model, BDM),来模拟北美大陆季节性干热带森林生物群区的分布范围。Deblauwe等[54]训练了一个经验预测模型(MaxEnt)来预测干旱半干旱地区周期性植被的存在概率,探讨了植被的自组织现象。本研究尝试通过MaxEnt模型建立气候因子与草原植被类型的关系模型,模拟内蒙古草原植被地理分布对气候变化的响应,同样取得了较好的模拟效果。这种研究扩展了BEMs的应用范围,有助于理解生态系统和高级分类单元(个体和群落水平之上)的功能多样性[51]。
MaxEnt模型的一个优势是可以对环境变量的重要性进行评价。湿润指数和降水的贡献率最高,表明水分是决定内蒙古草原植被分布格局最重要的因素。气温年较差和5℃积温的贡献率最低,表明内蒙古草原对热量的波动并不敏感。
基于Maxent模型的模拟结果,1961—2010年整个草原分布面积减少了11%,尤其是荒漠草原退化面积较大,与近年来内蒙古草原的实地调查结果相符[55]。草甸草原面积的减少主要是典型草原向其扩张导致的。典型草原被草甸草原和荒漠草原取代的面积要小于其向后两种草原扩张的面积,荒漠草原被典型草原取代的面积仅占荒漠草原总面积的15%,表明部分典型草原和荒漠草原的分布区域不再适宜草原植被生存,整个草原面积的减少预示着草原向着荒漠化的方向发展。尽管草原植被分布对温度的变化并不敏感,但温度升高会间接地加速草原退化,因为温度升高使潜在蒸散增加,即使降水不发生改变,植被也会遭受水分亏缺[56],进而影响到草原植被分布。
由于本研究没有考虑土地利用和过度放牧等因素,因此研究结果是区域尺度上基于气候适宜性的草原植被潜在变化,而结合地形、土壤类型、人为活动等因素的研究有待进一步开展。本研究模拟的是历史气候条件下植被分布状况,20世纪`80年代之后遥感技术的飞速发展为研究植被覆盖提供了新的手段,如何结合遥感数据进一步验证模拟的准确性,以及将该方法应用于未来气候情景预测植被结构和功能对未来气候变化的响应,有待进一步研究。
4 结论
本研究基于MaxEnt模型和综合模型模拟了1961—2010年内蒙古草原植被的地理分布和净第一性生产力变化。研究表明,湿润指数(MI)、年降水量(P)、最暖月平均温度(Tw)和最冷月平均温度(Tc)是影响草原植被地理分布的主导气候因子。MaxEnt模型能够很好地模拟内蒙古草原植被的地理分布,Kappa总体一致性值为0.76,草甸草原、典型草原和荒漠草原的Kappa一致性值分别为0.95、0.70和0.57。1961—2010年内蒙古草甸草原、典型草原和荒漠草原分布面积分别减少了5%、1% 和62%,草原面积整体减少了11%,预示着草原向着荒漠化的方向发展。降水是决定内蒙古草原NPP波动的最重要因素。该研究有助于理解内蒙古草原生态系统结构和功能对气候变化的响应,为进一步预测未来气候变化对草原植被的不利影响提供有价值的参考。
[1]IPCC. Summary for Policymakers // Stocker T F, D Qin, G K. Plattner, M Tignor, S K Allen, J Boschung, A Nauels, Y Xia, V Bex, P M Midgley, eds. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge, United Kingdom and New York, NY, USA: Cambridge University Press, 2013.
[2]Weltzin J F, Loik M E, Schwinning S, Williams D G, Fay P A, Haddad B M, Harte J, Huxman T E, Knapp A K, Lin G H, Pockman W T, Shaw M R, Small E E, Smith M D, Smith S D, Tissue D T, Zak J C. Assessing the response of terrestrial ecosystems to potential changes in precipitation. Bioscience, 2003, 53(10): 941- 952.
[3]Nemani R R, Keeling C D, Hashimoto H, Jolly W M, Piper S C, Tucker C J, Myneni R B, Running S W. Climate-driven increases in global terrestrial net primary production from 1982 to 1999. Science, 2003, 300(5625): 1560- 1563.
[4]Sala O E, Chapin F S, Armesto J J, Berlow E, Bloomfield J, Dirzo R, Huber-Sanwald E, Huenneke L F, Jackson R B, Kinzig A, Leemans R, Lodge D M, Mooney H A, Oesterheld M, Poff N L, Sykes M T, Walker B H, Walker M, Wall D H. Biodiversity-Global biodiversity scenarios for the year 2100. Science, 2000, 287(5459): 1770- 1774.
[5]Vitousek P M, Mooney H A, Lubchenco J, Melillo J M. Human domination of Earth′s ecosystems. Science, 1997, 277(5325): 494- 499.
[6]Field C B, Chapin F S, Matson P A, Mooney H A. Responses of terrestrial ecosystems to the changing atmosphere-a resource-based approach. Annual Review of Ecology and Systematics, 1992, 23(1): 201- 235.
[7]Zhang G G, Kang Y M, Han G D, Sakurai K. Effect of climate change over the past half century on the distribution, extent and NPP of ecosystems of Inner Mongolia. Global Change Biology, 2011, 17(1): 377- 389.
[8]Wang X M, Chen F H, Dong Z B. The relative role of climatic and human factors in desertification in semiarid China. Global Environmental Change, 2006, 16(1): 48- 57.
[9]Zhou G, Wang Y, Wang S. Responses of grassland ecosystems to precipitation and land use along the Northeast China Transect. Journal of Vegetation Science, 2002, 13(3): 361- 368.
[10]马瑞芳, 李茂松, 马秀枝. 气候变化对内蒙古草原退化的影响. 内蒙古气象, 2011, (2): 30- 39.
[11]Meyer N. Desertification and restoration of grasslands in Inner Mongolia. Journal of Forestry, 2006, 104(6): 328- 331.
[12]叶笃正, 丑纪范, 刘纪远, 张增祥, 王一谋, 周自江, 鞠洪波, 黄签. 关于我国华北沙尘天气的成因与治理对策. 地理学报, 2000, 55(5): 513- 521.
[13]尹燕亭, 侯向阳, 运向军. 气候变化对内蒙古草原生态系统影响的研究进展. 草业科学, 2011, 28(6): 1132- 1139.
[14]苏吉安, 朱幼军, 哈斯. 内蒙古草地生态问题及其对策探讨. 中国草地, 2003, 25(6): 68- 71.
[15]周广胜, 王玉辉. 全球生态学. 北京: 气象出版社, 2003.
[16]Roy P S, Joshi P K, Singh S, Agarwal S, Yadav D, Jegannathan C. Biome mapping in India using vegetation type map derived using temporal satellite data and environmental parameters. Ecological Modelling, 2006, 197(1/2): 148- 158.
[17]Phillips S J, Anderson R P, Schapire R E. Maximum entropy modeling of species geographic distributions. Ecological Modelling, 2006, 190(3/4): 231- 259.
[18]杨志香, 周广胜, 殷晓洁, 贾丙瑞. 中国兴安落叶松天然林地理分布及其气候适宜性. 生态学杂志, 2014, 33(6): 1429- 1436.
[19]殷晓洁, 周广胜, 隋兴华, 何奇瑾, 李荣平. 蒙古栎地理分布的主导气候因子及其阈值. 生态学报, 2013, 33(1): 103- 109.
[20]段居琦, 周广胜. 我国双季稻种植分布的年代际动态. 科学通报, 2013, 58(13): 1213- 1220.
[21]何奇瑾, 周广胜. 我国玉米种植区分布的气候适宜性. 科学通报, 2012, 57(4): 267- 275.
[22]孙敬松, 周广胜. 利用最大熵法(MaxEnt)模拟中国冬小麦分布区的年代际动态变化. 中国农业气象, 2012, 33(4): 481- 487.
[23]Roy J, Saugier B, Mooney H A. Terrestrial Global Productivity. San Diego: Academic Press, 2001.
[24]Odum E P. Fundamentals of Ecology. Philadelphia: W. B. Saunders, 1976.
[25]Leith H, R. H. W. Modeling the primary productivity of the world // Primary Productivity of the Biosphere. New York: Springer, 1975: 237- 263.
[26]Uchijima Z, Seino H. Agroclimatic evaluation of net primary productivity of natural vegetation (1) Chikugo model for evaluating net primary productivity. Journal of Agricultural Meteorological, 1985, 40(4): 343- 352.
[27]周广胜, 张新时. 自然植被净第一性生产力模型初探. 植物生态学报, 1995, 19(3): 193- 200.
[28]Gang C C, Zhou W, Li J L, Chen Y Z, Mu S J, Ren J Z, Chen J M, Groisman P Y. Assessing the Spatiotemporal Variation in Distribution, Extent and NPP of Terrestrial Ecosystems in Response to Climate Change from 1911 to 2000. Plos One, 2013, 8(11).
[29]Zhou G S, Wang Y H, Jiang Y L, Yang Z Y. Estimating biomass and net primary production from forest inventory data: a case study of China′sLarixforests. Forest Ecology and Management, 2002, 169(1/2): 149- 157.
[30]普宗朝, 张山清, 王胜兰. 近47年天山山区自然植被净初级生产力对气候变化的响应. 中国农业气象, 2009, 30(3): 283- 288.
[31]李镇清, 刘振国, 陈佐忠, 杨宗贵. 中国典型草原区气候变化及其对生产力的影响. 草业学报, 2003, 12(1): 4- 10.
[32]Akiyama T, Kawamura K. Grassland degradation in China: methods of monitoring, management and restoration. Grassland Science, 2007, 53(1): 1- 17.
[33]Wullschleger S D, Epstein H E, Box E O, Euskirchen E S, Goswami S, Iversen C M, Kattge J, Norby R J, van Bodegom P M, Xu X F. Plant functional types in Earth system models: past experiences and future directions for application of dynamic vegetation models in high-latitude ecosystems. Annals of Botany, 2014, 114(1): 1- 16.
[34]陈新美, 雷渊才, 张雄清, 贾宏炎. 样本量对MaxEnt模型预测物种分布精度和稳定性的影响. 林业科学, 2012, 48(1): 53- 59.
[35]Thornton P E, Running S W, White M A. Generating surfaces of daily meteorological variables over large regions of complex terrain. Journal of Hydrology, 1997, 190(3/4): 214- 251.
[36]Thornton P E, Running S W. An improved algorithm for estimating incident daily solar radiation from measurements of temperature, humidity, and precipitation. Agricultural and Forest Meteorology, 1999, 93(4): 211- 228.
[37]Woodward F I. Climate and plant distribution. Cambridge: Cambridge University Press, 1987.
[38]翁恩生, 周广胜. 用于全球变化研究的中国植物功能型划分. 植物生态学报, 2005, 29(1): 81- 97.
[39]张新时. 植被的PE(可能蒸散)指标与植被-气候分类(一)——几种主要方法与PEP程序介绍. 植物生态学与地植物学学报, 1989, 13(1): 1- 9.
[40]Phillips S J, Dudik M. Modeling of species distributions with Maxent: new extensions and a comprehensive evaluation. Ecography, 2008, 31(2): 161- 175.
[41]方精云. 全球生态学: 气候变化与生态响应. 北京: 高等教育出版社, 2000.
[42]Bai Y F, Han X G, Wu J G, Chen Z Z, Li L H. Ecosystem stability and compensatory effects in the Inner Mongolia grassland. Nature, 2004, 431(7005): 181- 184.
[43]Lauenroth W K, Sala O E. Long-term forage production of North American shortgrass steppe. Ecological Applications, 1992, 2(4): 397- 403.
[44]Knapp A K, Smith M D. Variation among biomes in temporal dynamics of aboveground primary production. Science, 2001, 291(5503): 481- 484.
[45]毛飞, 张艳红, 侯英雨, 唐世浩, 卢志光, 张佳华. 藏北那曲地区草地退化动态评价. 应用生态学报, 2008, 19(2): 278- 284.
[46]Li S, Verburg P H, Lv S, Wu J, Li X. Spatial analysis of the driving factors of grassland degradation under conditions of climate change and intensive use in Inner Mongolia, China. Regional Environmental Change, 2012, 12(3): 461- 474.
[47]Franklin J, Davis F W, Ikegami M, Syphard A D, Flint L E, Flint A L, Hannah L. Modeling plant species distributions under future climates: how fine scale do climate projections need to be?. Global Change Biology, 2013, 19(2): 473- 483.
[48]Guisan A, Thuiller W. Predicting species distribution: offering more than simple habitat models. Ecology Letters, 2005, 8(9): 993- 1009.
[49]Pearson R G, Dawson T P. Predicting the impacts of climate change on the distribution of species: are bioclimate envelope models useful? Global Ecology and Biogeography, 2003, 12(5): 361- 371.
[50]Chapman D S, Purse B V. Community versus single-species distribution models for British plants. Journal of Biogeography, 2011, 38(8): 1524- 1535.
[51]Pau S, Edwards E J, Still C J. Improving our understanding of environmental controls on the distribution of C3and C4grasses. Global Change Biology, 2013, 19(1): 184- 196.
[52]Driver A, Sink K J, Nel J N, Holness S, Van Niekerk L, Daniels F, Madjiet P A, Jonas Z, Maze K. National Biodiversity Assessment 2011: An assessment of South Africa′s biodiversity and ecosystems. Synthesis Report. Pretoria: South African National Biodiversity Institute and Department of Environmental Affairs, 2012.
[53]Särkinen T, Iganci J R, Linares-Palomino R, Simon M F, Prado D E. Forgotten forests-issues and prospects in biome mapping using Seasonally Dry Tropical Forests as a case study. BMC Ecology, 2011, 11(1): 27.
[54]Deblauwe V, Barbier N, Couteron P, Lejeune O, Bogaert J. The global biogeography of semi-arid periodic vegetation patterns. Global Ecology and Biogeography, 2008, 17(6): 715- 723.
[55]邢旗. 内蒙古草业研究. 呼和浩特: 内蒙古人民出版社, 2004.
[56]Hickler T, Vohland K, Feehan J, Miller P A, Smith B, Costa L, Giesecke T, Fronzek S, Carter T R, Cramer W, Kuhn I, Sykes M T. Projecting the future distribution of European potential natural vegetation zones with a generalized, tree species-based dynamic vegetation model. Global Ecology and Biogeography, 2012, 21(1): 50- 63.
Changes in distribution and productivity of steppe vegetation in Inner Mongolia during 1961 to 2010: Analysis based on MaxEnt model and synthetic model
YE Yongchang1,3, ZHOU Guangsheng1,2,*, YIN Xiaojie1,3
1StateKeyLaboratoryofVegetationandEnvironmentalChange,InstituteofBotany,ChineseAcademyofSciences,Beijing100093,China2ChineseAcademyofMeteorologicalSciences,Beijing100081,China3UniversityofChineseAcademyofSciences,Beijing100049,China
Global temperature and precipitation are undergoing unprecedented change, which has seriously affected the structure and function of steppe ecosystems in Inner Mongolia. The vegetation of this region has been suffering from severe salinization, desertification, and decline in productivity, resulting in severely ecological problems and economic losses. Although numerous studies indicate that desertification is intensifying currently, but the history of vegetation cover and geographical distribution patterns of vegetation are poorly understood. This research attempts to simulate the temporal and spatial changes in the geographical distribution of steppe vegetation based on MaxEnt and changes of net primary productivity (NPP) based on synthetic model in Inner Mongolia during 1961 to 2010. The aim of this research is to provide a valuable method for predicting the potential impact of climate change on vegetation in the future. The results show that moisture index (MI), annual precipitation (P), mean temperature of the warmest month (Tw), and mean temperature of the coldest month (Tc) are the dominant climatic factors determining the geographical distribution of steppes. The MaxEnt model performs well in simulating the geographical distribution of steppe vegetation in Inner Mongolia. The AUC of meadow steppe, typical steppe, and desert steppe are about 0.90 (excellent), 0.85 (very good), and 0.95 (excellent), respectively. The kappa consistencies of meadow steppe, typical steppe, and desert steppe are about 0.95 (excellent), 0.70 (very good), and 0.57 (very good), respectively. The overall kappa value of the entire steppe vegetation is about 0.76 (very good). The areas characterized by steppe vegetation in Inner Mongolia have shrunk during 1961 to 2010. The distribution area decreased to about 5% for meadow steppe, 1% for typical steppe, 62% for desert steppe, and 11% for the entire steppe. The gravity centers of both meadow steppe and typical steppe have moved towards southwest, while the desert steppe moved towards southwest on the initial stage and to the northeast later. Due to these shifts the entire range of steppe are confined within narrow distribution boundary. The area converted from meadow steppe to typical steppe (2.38 × 104km2) is less than the area of transformation of the latter to the former (3.57 × 104km2). The area transformed from desert steppe to typical steppe accounts for 15% of the total area of desert steppe. The change in areas shows that the Inner Mongolia steppe is undergoing desertification. The NPP of the entire steppe, meadow steppe, typical steppe, and desert steppe were 312 gDWm-2a-1, 341 gDWm-2a-1, 309 gDWm-2a-1, and 260 gDWm-2a-1in Inner Mongolia during 1961 to 2010. The change in NPP of the entire steppe followed a decline-increase-decline trend during 1961 to 2010. Regional precipitation is the primary determining factor for the NPP of steppes in Inner Mongolia. The MaxEnt model is one of the bioclimatic envelope models (BEMs) based on the niche theory. BEMs are often used to solve problems in conservation biology, biogeography, ecology, and taxonomy at species level. We have successfully used the MaxEnt model to simulate the geographical distribution of the different steppe vegetation types. This study extends the range of application of BEMs, which has facilitated the understanding of the functional diversities of ecological systems and advanced classification units (above the individual and community level).
Inner Mongolia Steppe; MaxEnt model; geographical distribution of steppe vegetation; synthetic model; net primary productivity
国家重点基础研究发展计划(973计划)项目(2010CB951303);中国科学院战略性先导科技专项-应对气候变化的碳收支认证及相关问题(XDA05050408)
2014- 12- 30;
2015- 12- 08
Corresponding author.E-mail: gszhou@ibcas.ac.cn
10.5846/stxb201412302599
叶永昌,周广胜,殷晓洁.1961—2010年内蒙古草原植被分布和生产力变化——基于MaxEnt模型和综合模型的模拟分析.生态学报,2016,36(15):4718- 4728.
Ye Y C, Zhou G S, Yin X J.Changes in distribution and productivity of steppe vegetation in Inner Mongolia during 1961 to 2010: Analysis based on MaxEnt model and synthetic model.Acta Ecologica Sinica,2016,36(15):4718- 4728.
