3种室内观叶植物光合特性的比较研究
2016-07-29陈海霞
陈海霞,范 适
(1.湖南农业大学 园艺园林学院,湖南 长沙 410128; 2.湖南环境生物职业技术学院 园林学院,湖南 衡阳 421005)
3种室内观叶植物光合特性的比较研究
陈海霞1,范适2
(1.湖南农业大学 园艺园林学院,湖南 长沙 410128; 2.湖南环境生物职业技术学院 园林学院,湖南 衡阳 421005)
摘要:观叶植物是居室绿化的重要材料.为了合理进行居室植物设计,采用3种常见室内观叶植物豆瓣绿、吊兰、铁线蕨为试材,分别将其置于居室的东、南和西面,再利用LI-6400便携式光合测定仪测定这3种植物的光合特性.结果表明:3种植物的光补偿点分别为25.7、29.6和13.0 μmol·m-2·s-1;光饱和点分别约为1 300、1 400、1 200 μmol·m-2·s-1.由此可见,铁线蕨对光的适应性较强,是一种荫性植物,而吊兰和豆瓣绿则是半荫性植物,这为观叶植物的室内布置提供了科学依据.图6,表2,参10.
关键词:观叶植物;净光合速率;光饱和点;光补偿点
人类80%~90%的时间是在室内度过,室内环境相对独立和隔离,加之建筑材料的污染,据调查结果表明人们生活的室内环境的空气质量直接影响到人们的身体健康和生活质量[1].因此,室内绿化逐渐受到青睐,室内植物的应用越来越广泛,形式也多样化,已由过去的不注重室内植物的摆放,发展为室内绿化设计,更注重植物对室内环境生态质量的影响;室内植物种植设计也是缓解人类精神压力的一种有效途径[2].但是受到室内光照、温度和湿度等环境条件的限制,植物生长发育也受到影响,因此,根据居室不同方向的光照条件进行植物种植设计,是维持室内植物景观的前提条件.作者以3种常见的室内观叶植物为研究对象,分别置于室内的东、南和西面,对其在室内不同自然光照条件下的光合特性进行比较分析,旨在为居室植物的绿化设计和应用提供参考依据.
1材料与方法
1.1材料
实验于2014年11月在湖南农业大学园艺园林学院观赏园艺研究所进行.以长势一致的三年生豆瓣绿、吊兰和铁线蕨为试材,分别在居室的东、南和西面放置3种观叶植物各3盆,选取每盆植株中部的功能叶3片进行测定.
豆瓣绿(Peperomia tetraphylla)胡椒科椒草属多年生草本植物,叶片肥厚,光亮翠绿,四季常青,株型美观,给人以小巧玲珑之感,深受人们喜爱.
吊兰(Chlorophytum elatum)百合科吊兰属多年生草本植物,肉质根,叶细长,似兰花;其性喜温暖湿润、半阴的环境,为室内绿化常用植物.
铁线蕨(Adiantum capillus veneris)是多年生常绿草本植物,叶片轻盈秀美,畏干燥及强烈光照,在10~20 ℃温度下生长良好,冬季温度高于5 ℃,才能安全越冬.
1.2居室环境条件

表1 居室不同方向的环境条件
注:以上数据为测定时间段的环境参数
选取室内南、西、东3个方向作为试验环境,将供试植物置于以上环境30 d后进行各项指标的测定.
1.3测量指标
采用LI-6400XT便携式光合仪(LI-COR)测定光合日变化褐光响应曲线.
(1)光合日变化的测定.选择一大晴天,使用自然光源,CO2为环境浓度,流量设定为500 μmol s-1,从8∶00-18∶00分别对放置在南、西、东面的3种室内观叶植物进行测定,每2 h测定1次.
(2)光响应曲线的测定.选择晴天的上午8∶00~11∶30,使用LED红蓝光源,CO2为环境浓度,流量设定为500 μmol s-1,将光照强度(PAR)分别设定15个水平(2 000、1 700、1 400、1 200、1 000、800、600、400、200、150、100、80、50、20、0 μmol·m-2·s-1).采用自动测量系统Auto Prog,设定最小等待时间(120 s)、最大等待时间(200 s),ΔCO2绝对值低于20去匹配.最大净光合速率为光饱和时的净光合速率,以光强为横坐标,净光合速率为纵坐标,绘制光响应曲线;由Pn-PAR响应曲线的回归方程y=ax2+ bx+ c(x为光量子强度,y为净光合速率) 获得光补偿点及光饱和点的值.
1.4数据处理
采用Excel2007对获得数据进行整理和分析.
2结果与分析
2.13种观叶植物在不同环境下的光合日变化
光合作用是一个光生物化学反应,所以光合速率随着光照强度的增加而加快.净光合速率是光合系统功能的直接体现,也是植株光合系统工作正常与否的指标.植物的生态型影响其对光照的响应方式,在强光和弱光下其光合速率变化也均有差异[3-4].在自然条件下,植物光合速率的日变化曲线有“单峰和双峰”两种类型,植物在晴天午时前后光合速率达到顶点即能形成单峰曲线;如植物在中午光合速率下降(曲线的低谷),即出现“午休”现象,然后再上升则形成双峰曲线[5].
如图1所示,豆瓣绿B和C环境下净光合速率呈双峰变化曲线,两个峰值分别在12∶00以及16∶00出现,午后有“午休”现象,最大净光合速率为3.72 μmol·m-2·s-1;在A环境中,豆瓣绿净光合速率呈单峰变化曲线,且在14∶00达到最高峰值,最大净光合速率分别为1.20 μmol·m-2·s-1.
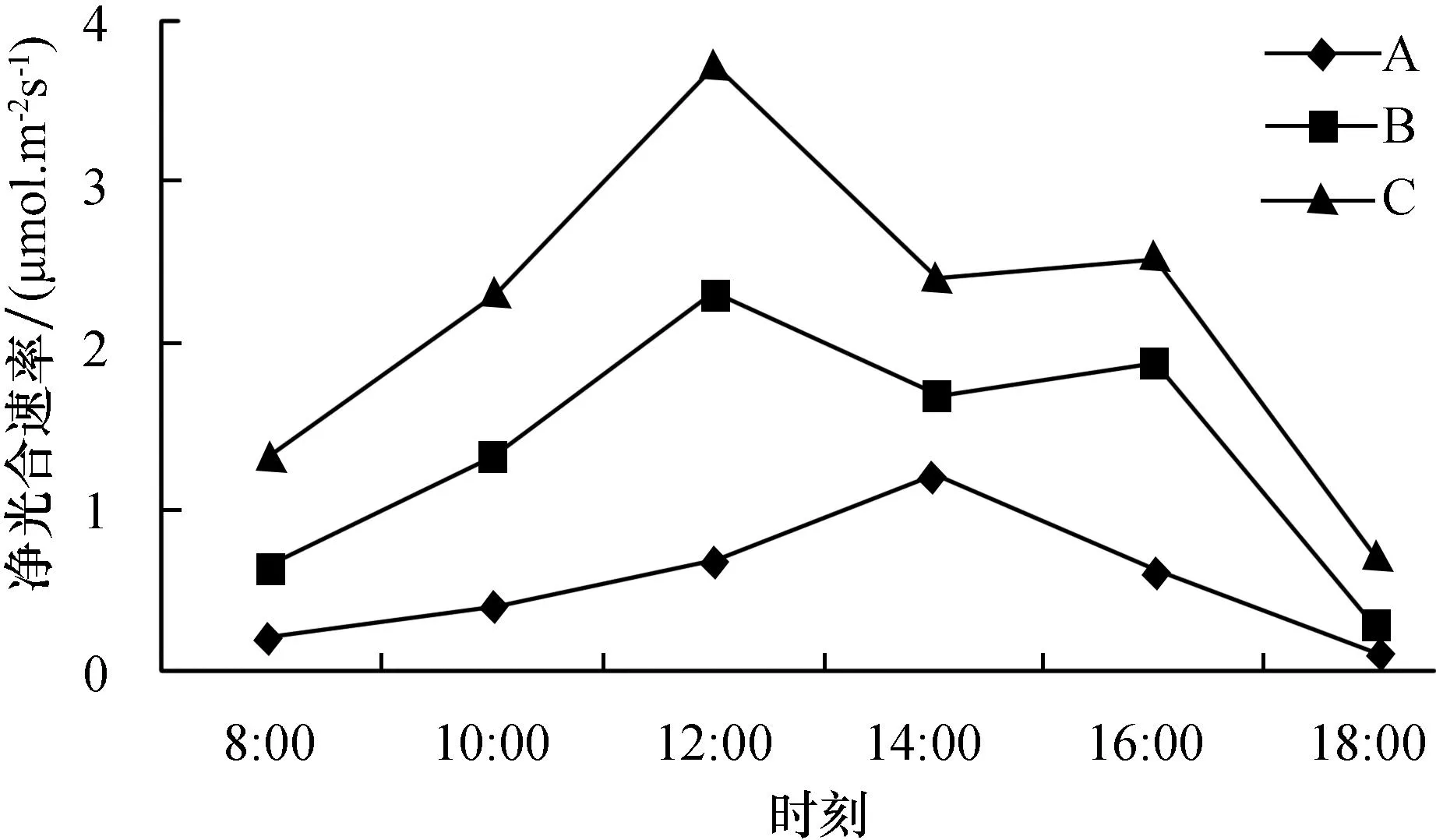
图1 豆瓣绿在不同环境下的净光合速率日变化Fig.1 Diurnal variation of net photosynthetic rate under different circumstances of peperomia tetraphylla
吊兰的净光合速率变化曲线如图2所示,在A和B环境下净光合速率的变化曲线呈双峰,分别在12∶00和16∶00出现,最大的净光合速率为5.11 μmol·m-2·s-1;在环境中,吊兰的净光合速率为单峰变化曲线,12∶00时达到峰值为1.4 μmol·m-2·s-1.

图2 吊兰在不同环境下的净光合速率日变化Fig.2 Diurnal variation of net photosynthetic rate under different circumstances of chlorophytum elatum
如图3所示,铁线蕨在B和C环境下净光合速率呈双峰变化曲线,分别在12∶00和16∶00出现峰值,其中在C环境下12∶00的净光合速率最大,其值为3.87 μmol·m-2·s-1;在A环境中,铁线蕨的净光合速率为单峰变化曲线,14∶00时达到峰值,为3.2 μmol·m-2·s-1.
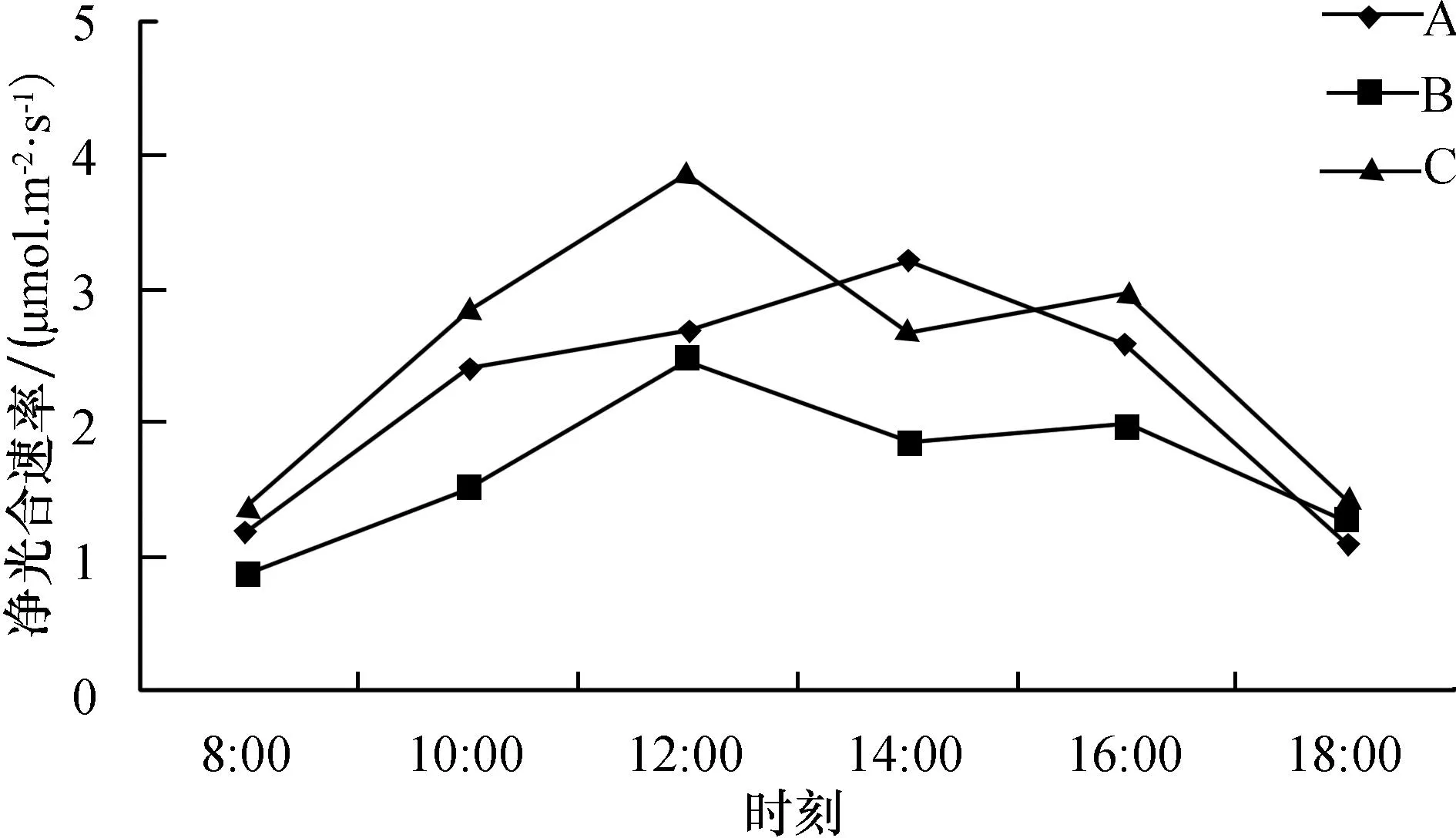
图3 铁线蕨在不同环境下的净光合速率日变化Fig.3 Diurnal variation of net photosynthetic rate under different circumstances of adiantum capillus veneris
2.23种观叶植物在不同环境下的光补偿点和光饱和点
光合作用是非常复杂的光生物化学反应,其光合速率会随着光照强度的变化而变化,光响应曲线则反映了这种变化规律.光饱和点则反映了植物利用强光的能力,不同植物的光饱和点受到叶片特征的影响.在自然条件下,测定在不同光合有效辐射下所对应的净光合速率,做出散点图,得到其光响应曲线图及回归方程,再从曲线上判断和计算植物的光补偿点和光饱和点[6-8].根据光饱和点和光补偿点的值判断植物对光照条件的需求,以及植物喜阳和耐阴的性能.
由表2和图4~6可知,这3种植物对光照的适应范围较大,在弱光情况下也能维持一定的净光合速率;在光照强度较高时,净光合速率也有较高水平.当光合有效辐照为0 μmol·m-2·s-1时,3种植物的净光合速率均为负值;当光合有效辐照达到50 μmol·m-2·s-1时,铁线莲和豆瓣绿开始进行光合作用,而吊兰的净光合速率还为负值;当光照强度小于700 μmol·m-2·s-1时,3种植物的净光合速率都随光照强度的增强而显著增加;当光照强度继续增强时,净光合速率增加的速度逐渐缓慢;当达到一定值时,3种植物的净光合速率均不再增加,而是维持在一定水平,即达到光饱和点.从图中可见,豆瓣绿的光补偿点和光饱和点分别为25.7 μmol·m-2·s-1,1 100 μmol·m-2·s-1;吊兰的光补偿点和光饱和点分别为29.6 μmol·m-2·s-1,1 400 μmol·m-2·s-1;铁线蕨的光补偿点和光饱和点分别为13.0 μmol·m-2·s-1,1 150 μmol·m-2·s-1.比较3种植物的光补偿点和光饱和点可知,铁线蕨的光补偿点最低,其利用弱光的能力最强,其耐阴性也最好;吊兰光补偿点最高,利用弱光的能力相对较低,能耐较强光,光饱和点最高,说明其适应光照的范围最大.

表2 3种室内观叶植物的光强响应值
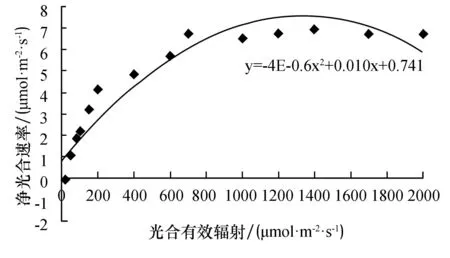
图4 豆瓣绿的光响应曲线Fig.4 The light response curve of peperomia tetraphylla
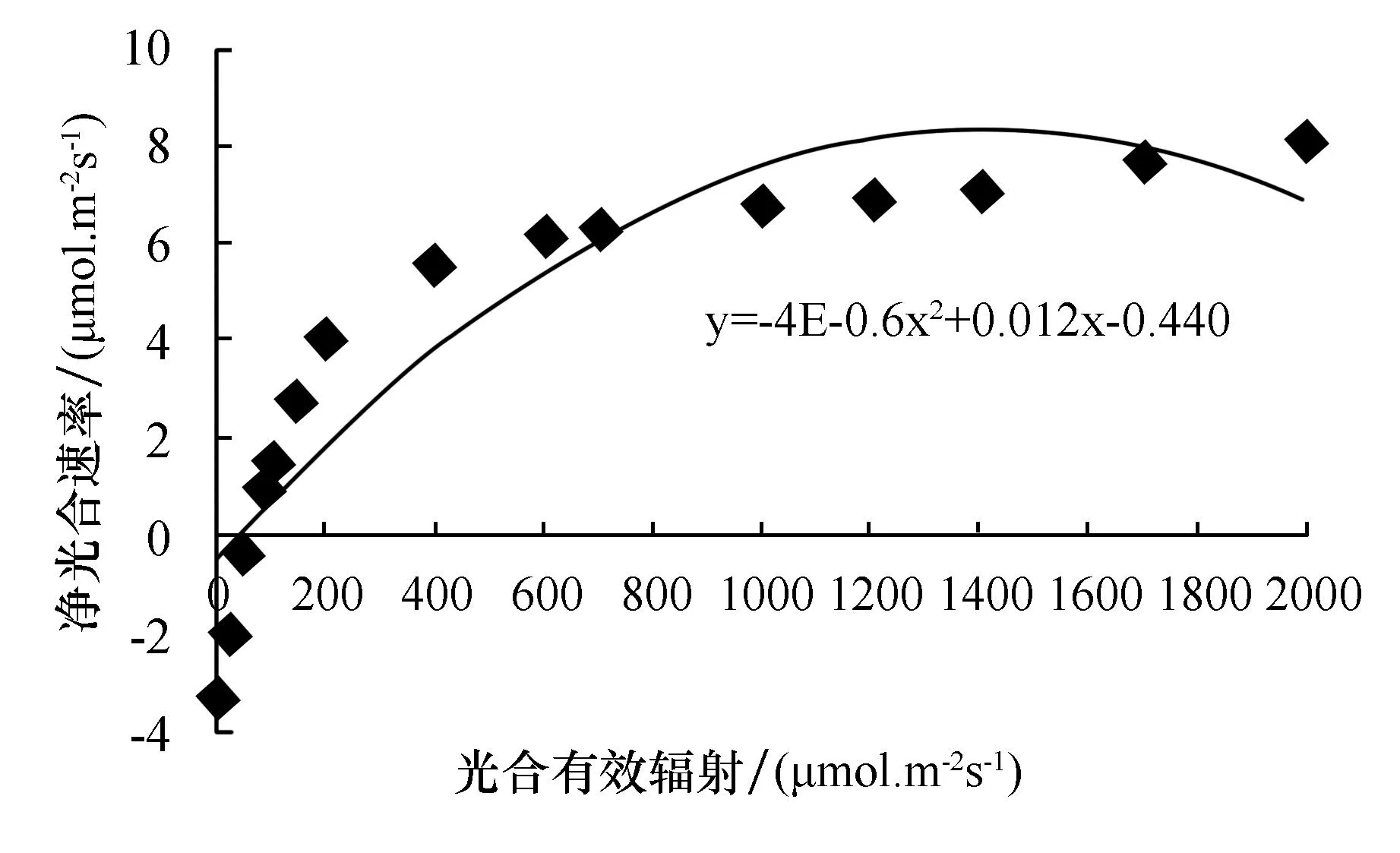
图5 吊兰的光响应曲线Fig.5 The light response curve of chlorophytum elatum
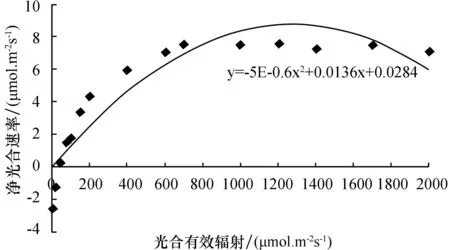
图6 铁线蕨的光响应曲线Fig.6 The light response curve of adiantum capillus veneris
3讨论
植物光合作用是十分复杂的生理过程,其自身叶绿体含量、叶片特性及环境因子都会影响叶片净光合速率[9].一天之中,由于外界光照强度、气温、空气中CO2浓度以及植物体内水分、光合中间产物及气孔的开放度等不断变化,植物的净光合速率也有明显的日变化.本试验结果表明,相同植物放置在室内不同的方向,由于光照强度的不同,叶片净光合速率的日变化呈现不同的峰,其中豆瓣绿和铁线蕨有相同的变化规律,在东面和西面的净光合速率日变化呈双峰变化曲线.
通过分析光合响应曲线,可以判断植物光合作用的光饱和点、光补偿点等重要数值,这两组数值很好地反映了植物对光照条件的要求,和阳性植物相比耐阴植物的光饱和点和光补偿点均较低[10].因此,不同光强下的净光合速率以及对不同光强的反应可以作为鉴定植物耐荫性能的生理指标.本试验中,由3种室内观叶植物的光响应曲线可知,3种植物在弱光条件下的光合作用均能正常进行;随着光照强度的不断增加,净光合速率都呈现增加趋势,并在1 000 μmol·m-2·s-1左右达到饱和点,说明3种植物对光的适应性都较强.其中铁线蕨的光补偿点和光饱和点均较低,豆瓣绿次之,因此这两种植物利用弱光的能力比吊兰强,耐阴性较好,在半阴状态下的生长状况更好,因此能很好地适应居室环境.
参考文献:
[1] 李磊,王逢瑚,安小雷.室内自然景观对室内环境调节净化的作用[J].东北林业大学学报,2006,34(2):65-69.
Li Lei,Wang Feng-hu,An Xiao-lei.Effect of regulation and purification of indoor natural and scape on indoor environment[J].Journal of Northeast Forestry University,2006,34(2): 65-69.
[2] 苏雪痕.植物造景[M].北京:中国林业出版社,1994.
Su Xue-hen.Plant landscape[M].Beijing: China Forestry Publishing House,1994.
[3] 朱万泽,王金锡,薛建辉.台湾桤木引种的光合生理特性研究[J].西北植物学报,2004,24(11):2012-2019.
Zhu Wan-ze,Wang Jin-xi,Xue Jian-hui.Study on the photosynthetic characteristics of introduced Alnus Formosana[J].Acta Bot.Boreal-Occident.Sin.2004,24(11):2012-2019.
[4] 贺安娜,伍贤进,李胜华,等.虎耳草春叶和秋叶光合特性及有效成分含量比较[J].江苏农业科学,2016,44(1):224-227.
He An-na,Wu Xian-jin,Li Sheng-hua,et al.comparative study on photosynthetic characteristics and active ingredient content of spring leaves and autumn leaves of Saxifraga stolonifera Curt.[J].Journal of Jiangsu Agricultural Sciences,2016,44(1):224-227.
[5] 邓明净,孙振委,张洪燕,等.红富士苹果盆景叶片净光合速率的研究[J].现代农业科技,2007,13:15-16.
Deng Ming-jing,Sun Zhen-wei,Zhang Hong-yan,et al.Study on net photosynthetic rate of bonsai leaves of Red Fuji apple[J].Journal of Modern Agricultural Science and Technology,2007,13:15-16.
[6] 尤扬,贾文庆,周建,等.黄栌叶片光合特性[J].东北林业大学学报,2009,37(7):25-26.
You Yang,Jia Wen-qing,Zhou Jian,et al.Photosynthetic characteristics of cotinus coggygria leaves[J].Journal of Northeast Forestry University,2009,37(7):25-26.
[7] 蒋高明,何维明.一种在野外自然光照条件下快速测定光合作用光响应曲线的新方法[J].植物学通报,1999,16(6):712-718.
Jiang Gao-ming,He Wei-ming.A quick new method for determining light response curves of photosynthesis under field light conditions[J].Chinese Bulletin of Botany,1999,16(6): 712-718.
[8] 张宝玉.葡萄光合作用光响应曲线的研究[D].杨凌:西北农林科技大学,2009
Zhang Bao-yu.Study on photosynthetic response curves of grape[D].Yangling:Northwest Agriculture and Forestry University,2009.
[9] 张治安,杨福,陈展宇,等.菰叶片净光合速率日变化及其与环境因子的相互关系[J].中国农业科学,2006,39(3):502-509.
Zhang Zhi-an,Yang Fu,Chen Zhan-yu,et al.Relationship between diurnal changes of net photosynthetic rate and environmental factors in leaves of Zizania latifolia[J].Scientia Agricultura Sinica,2006,39(3):502-509.
[10] 刘颖娇,李国防,王志博,等.中午适度遮阴对苹果叶片光合生理特性的影响[J].西北农业学报,2015,24(2):124-125.
Liu Ying-jiao,Li Guo-fang,Wang Zhi-bo,et al.Effect of moderately shade on photosynthetic characteristic in Malus domestica Borkh.Leaves during midday[J].Acta Agriculturae Boreali-occidentalis Sinica,2015,24(2):124-125.
Biography:CHEN Hai-xia,female,born in 1976,Ph.D.,assistant professor,research directions: Ornamental plant molecular biology
收稿日期:2016-03-12
基金项目:国家自然科学基金资助(编号:31201656);湖南省教育厅青年基金资助(编号:14B093);湖南省林业厅科技计划项目资助(编号:XLK201338)
作者简介:陈海霞(1976-),女,湖南张家界人,博士,副教授,研究方向:观赏植物分子生物学.
文章编号:2095-7300(2016)02-016-05
中图分类号:Q945.11
文献标识码:A
A Comparison of Characteristics of Photosynthetic of Three Indoor Ornamental Foliage Plants
CHEN Hai-xia1,FAN Shi2
(1.College of Horticulture and Landscape,Hunan Agriculture University,Changsha 410128,China; 2.Landscape Department, Hunan Polytechnic of Environment and Biological,Hengyang 421005,China)
Abstract:In this paper,three kinds of indoor foliage plants Peperomia te traphylla,Chlorophytum elatum and Adiantum capillus-veneris are for materials,LI-6400 portable photosynthesis analyzer of photosynthetic characteristics in the three light environment (east,west,south).The results indicated that light compensation point is respectively 25.7,29.6,13.0 μmol/(m2·s);Light saturation point is respectively 1 200,1 400,1 200 μmol/(m2·s).Results showed that Adiantum capillus-veneris had wider adaptability for light,but Peperomia tetraphylla and Adiantum capillus-veneris need much more light.6figs.,2tabs.,10refs.
Keywords:ornamental foliage plants;photosynthetic rate;light saturation point;light compensation point
