白粉病菌对不同抗性辣椒品种防御酶活性的影响及其互作超微结构分析
2016-07-25莫熙礼蒋选利武华文江厚成
莫熙礼++蒋选利++武华文++江厚成


摘要:为了探索白粉病菌对不同抗性辣椒品种防御酶活性的影响及其互作机制,采用酶标仪法测定病原菌对辣椒叶片防御酶活性的影响;用扫描电子显微镜和荧光显微镜对辣椒-白粉病菌互作过程进行超微结构观察。试验表明:接种白粉病菌后,抗性材料(花溪辣椒)的POD、PPO活性表现为峰值出现最早,感病材料(西安8819和遵义辣椒)表现为活性变化幅度大、峰值出现较晚;抗病材料和感病材料的PAL酶活性都在处理3 d到达峰值,抗病材料活性变化幅度大于感病材料;花溪辣椒(抗病)和遵义辣椒(感病)的几丁质酶和β-1,3-葡聚糖酶迅速上升,而西安8819(感病)的几丁质酶和β-1,3-葡聚糖酶活性上升速度较慢。接种白粉病菌后12 h,分生孢子萌发产生芽管和附着胞,24 h 形成菌丝,菌丝周边的蜡质组织开始被溶解,120 h菌丝从叶表面气孔伸出,形成为分生孢子梗,未见有从气孔侵入的迹象。从而可以得出结论:接种白粉病菌后,辣椒叶片防御酶活性均有不同程度的提高,不同辣椒品种的防御酶活性变化不同。
关键词:白粉病菌;辣椒;防御酶;超微结构
中图分类号: S436.418.1+9文献标志码: A文章编号:1002-1302(2016)06-0229-04
收稿日期:2016-01-04
基金项目:贵州省黔西南州科技局计划(编号:2015-05)。
作者简介:莫熙礼(1982—),男,广西梧州人,硕士,讲师,从事植物病虫害防治教学和科研工作。E-mail:moxili1982@163.com。辣椒白粉病是辣椒重要病害之一,病原菌的有性世代为子囊菌亚门内丝白粉菌属的鞑靼内丝白粉菌,无性世代为半知菌亚门拟粉孢霉真菌。辣椒白粉病主要危害辣椒叶片,影响光合作用,从而导致辣椒产量和品质降低。贵州是我国辣椒主产区之一,气候温暖湿润,非常适宜辣椒种植,且辣椒品种资源丰富。随着病菌多年的积累,辣椒白粉病在贵州各辣椒产地大面积发生,大大降低了辣椒的产量和品质,这给当地农业生产的发展和农民的脱贫致富造成了严重威胁。因此,对辣椒白粉病的系统研究迫在眉睫。目前,人们对辣椒白粉病方面的研究主要集中在病菌的鉴定[1]、分类以及化学药剂防治[2]等方面,而在辣椒与白粉病菌互作方面还不清楚。为此,本研究通过室内试验,测定白粉病菌侵染后辣椒POD等防御酶活性的变化以及利用电子显微镜和荧光显微镜进行白粉病菌-辣椒互作的超微结构观察,以期从生理生化和组织病理角度探讨白粉病菌-辣椒互作机制,为今后有效、绿色控制辣椒白粉病提供参考。
1材料与方法
1.1供试辣椒品种
供试辣椒品种为花溪辣椒、西安8819、遵义辣椒,其中花溪辣椒为抗病品种、西安8819和遵义辣椒为感病品种。消毒后,将种子播种在直径约6 cm、高约15 cm的塑料杯中,待幼苗长到8~10片真叶时,将植株移栽在直径约20 cm、高约 25 cm 的花盆中,保持每盆2~3株,置于温室培养。
1.2供试菌源
菌源为辣椒白粉病菌,孢子采自花溪区湖潮乡磊庄辣椒基地自然发病的植株,配制成孢子悬浮液(1×105个/mL)。
1.3接种方法
待供试辣椒长至25片真叶时,对每个辣椒品种的叶片进行喷菌接种处理,孢子浓度为1×105个/mL,每个材料接种20株,以清水喷雾为对照(CK)。接种后,马上取样进行防御酶活性测定,之后连续6 d同一时间取样检测。每个处理进行3次重复。
1.4粗酶液的提取
PPO和POD酶粗液的提取参照李靖等的方法[3];PAL酶粗提液的提取参考薛应龙等的方法[4];β-1,3-葡聚糖酶和几丁质酶粗提液的提取参考车海彦等的方法[5]。
1.5酶活性的测定
PPO和POD活性的测定参照李保聚等的方法[6];PAL 活性测定参照考薛应龙等的方法[4];β-1,3-葡聚糖酶活性测定参照车海彦等的方法[5];几丁质酶活性测定:胶态几丁质的制备采用陈鹏等的方法[7],几丁质酶活性的测定参照汤章城的方法[8]。
1.6白粉菌侵染超微结构分析
1.6.1材料辣椒品种:花溪辣椒。
1.6.2接种方法方法同“1.3”节,待辣椒长至25片真叶时,对辣椒叶片进行喷菌接种处理,孢子浓度为1×105个/mL,在处理后12 h、1 d、2 d、8 d剪取叶片进行以下观察。
1.6.3扫描电子显微镜和荧光显微镜观察(1)器材:扫描电子显微镜(S-3400N)。(2)方法:①取材:剪取辣椒叶片,长和宽1~2 cm;②清洗:用 5%碳酸钠冲洗辣椒叶片表面常细胞碎片、杂质。并粘在样品台上;③观察:将样品台放置冷冻台内,打开扫描电子显微镜观察、拍照。
1.6.4荧光显微镜观察(1)器材:荧光显微镜(Olympus 13X51T)、数码相机(佳能A640);(2)方法:采用Calcofluor荧光染色法进行染色观察。
2结果与分析
2.1白粉病菌对不同抗性辣椒品种防御酶活性的影响
2.1.1POD活性的变化由图1可见,花溪辣椒接种白粉病菌后,植株的POD活性明显增加,并在接种1 d达到高峰。西安8819接种白粉病菌后,植株的POD活性明显增加,并在接种5 d达到高峰。遵义辣椒接种白粉病菌后,接种植株的POD活性也迅速增加,并在接种4 d达到高峰。接种白粉病菌会促进辣椒植株POD酶活性的增强,抗病较强品种花溪辣椒的POD峰值出现较早,感病品种 (西安8819和遵义辣椒)的POD酶活性峰峰值出现较迟。由此推断出,辣椒抗病品种的POD酶对白粉病菌的敏感性较强。
2.1.2PPO活性的变化由图2可见,花溪辣椒接种白粉病菌后,PPO酶活性在接种2 d和接种4 d出现2个高峰期,分别比对照高58.6%和60.4%。西安8819辣椒植株在接种白粉病菌后,PPO酶活性在接种3 d出现高峰期,其值高于对照1003%。遵义辣椒仔接种白粉病菌后,PPO酶活性在接种 3 d 出现高峰期,也明显高于对照。感病品种挑战接种白粉菌后,PPO酶活性高于感病品种,但峰值出现的时间较晚。
2.1.3PAL活性的变化由图3可见,花溪辣椒接种白粉菌后,PAL酶活性在接种3 d达到最大,比CK高出23%。西安8819接种白粉病菌后,PAL酶活性均在处理后接种3 d达到高峰期,比CK高出35%。遵义辣椒接菌处理和CK的PAL酶活性在接种3 d和5 d出现2个高峰期,但差别不明显。由此推断出,抗病品种的PAL酶对白粉病菌敏感,接菌后活性迅速上升,而感病品种的PAL酶反应较慢,变化幅度不大。
2.1.4β-1,3-葡聚糖酶活性变化由图4可见,花溪辣椒接种白粉病菌后,植株的β-1,3-葡聚糖酶活性明显增加,高于CK处理。西安8819接种白粉病菌后,β-1,3-葡聚糖酶活性在接种3 d达到峰值,随后逐渐下降,在接种5 d与CK处于同一水平。遵义辣椒接菌处理和CK在接种后β-1,3-葡聚糖酶活性逐渐上升,接菌处理升高的幅度高于CK处理。
4 d时,接菌处理和CK的β-1,3-葡聚糖酶活性无明显差别。在整个检测期内,抗病品种(花溪辣椒)的β-1,3-葡聚糖酶活性低于感病品种。
2.1.5几丁质酶活性的变化由图5可知,接种白粉菌后,3个辣椒品种的几丁质酶活性均上升,只是增长的速度不同。花溪辣椒接种白粉病菌后,几丁质酶活性迅速升高,接种2 d 到达峰值,随后下降,接种5 d与CK无明显差别。西安8819接种白粉病菌后,几丁质酶活性在接种2 d和接种4 d出现2个峰值。遵义辣椒接种白粉病菌后,几丁质酶接种1 d就出现了峰值,随后逐渐下降。在整个检测期内,接菌处理的活性明显高于CK。
2.2辣椒与白粉菌互作超微结构分析
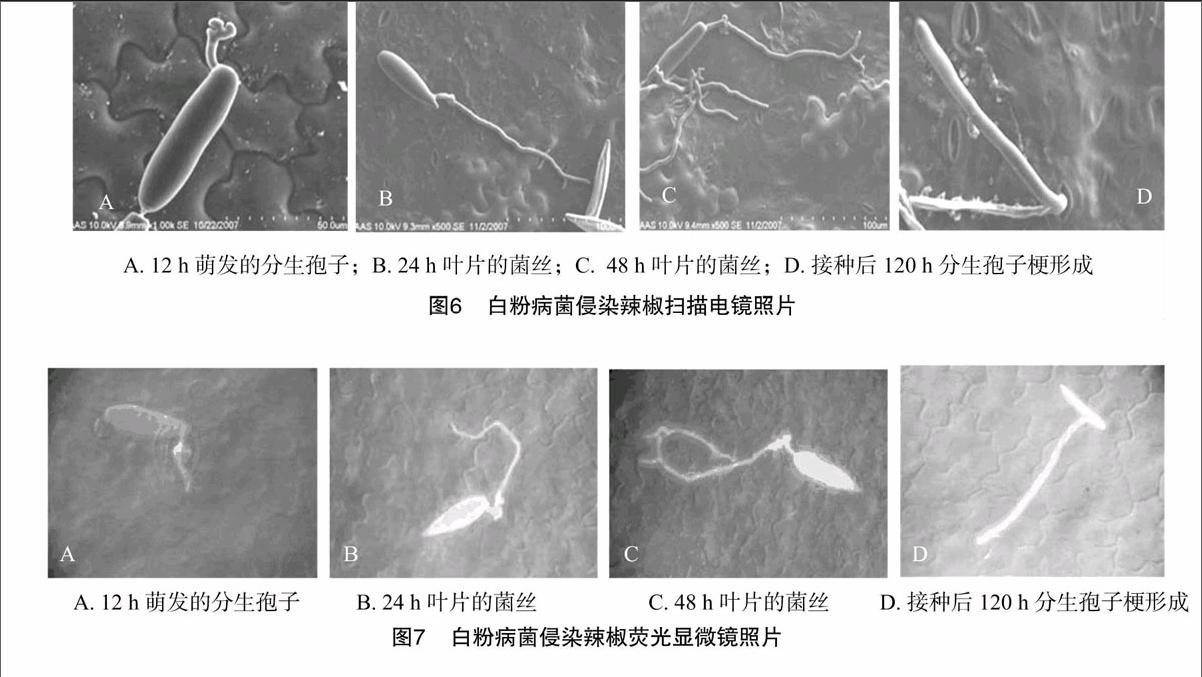
接种白粉病菌后12 h,辣椒叶片表面上的分生孢子萌发产生芽管,孢子萌发只产生1个芽管,可观察到芽管顶端形成圆形附着孢,并且附着孢开始产生菌丝,少数的孢子可以在侧面产生附着孢(图6-A、图7-A),主要起附着作用。接菌后24 h,菌丝形成后不立即侵入叶组织,而是在叶表面上扩展伸长,不断产生分枝(图6-B、图7-B)。从图6-B发现,菌丝周边的蜡质组织开始被溶解。接菌后48 h,扫描电镜观察发现,菌丝从气孔旁穿行而过,未见有从气孔侵入的迹象(图6-C、图7-C)。接菌后120 h,辣椒叶片表面产生许多褪绿斑,可发现叶片内菌丝从叶表面气孔伸出,形成为分生孢子梗,少部分还观察到分生孢子梗上产生孢子(图6-D、图7-D)。扫描电子显微镜和荧光显微镜观察,未发现菌丝从气孔侵入,且菌丝从气孔旁穿行而过,未见有从气孔侵入的迹象。
3讨论
国内外许多学者对防御酶活性与植物抗病性的关系进行了深入研究,寄主-病原物体系不同,得出的研究结果不尽相同[9-10]。以往研究认为:POD 参与形成对真菌孢子萌发具有直接抑制作用的物质——H2O2,且参与木质素和木栓质的合成;PPO酶可以催化植物产生对病原菌菌丝生长有毒性的物质(醌和单宁),参与催化酚类物质合成木质素的反应,促进细胞壁木质化以抵抗病原菌的侵害;PAL酶活性升高会促进木质素的积累、酚类物质和植保素的合成,增加细胞壁的厚度,形成抵抗病原入侵的机械屏障[11];几丁质酶和β-1,3-葡聚糖酶也是许多真菌细胞壁的主要组分之一,在植物的抗病性方面具有重要作用,能够降解真菌的细胞壁和抑制真菌的生长[11]。病原菌和化学因子都能诱导植物体内的POD、PPO、PAL、几丁质酶和β-1,3-葡聚糖酶酶活性的升高。
研究结果表明:与对照相比,接菌处理的辣椒叶片防御酶活性均有不同程度的提高,辣椒品种不同,防御酶活性的变化量不同。接种白粉病菌后,抗性材料花溪辣椒的POD、PPO活性表现为峰值出现最早,感病材料表现为活性变化量大、峰值出现较晚。由此认为,接种白粉病菌后,抗病材料的POD、PPO活性迅速到达峰值,形成抑菌物质和有毒物质,促进木质化抵抗白粉病菌的侵染,感病材料则反应较慢,没有迅速抵抗病菌的侵入。接种白粉病菌后,抗病材料和感病材料的PAL酶活性都在3 d时到达峰值,抗病材料活性变化幅度大于感病材料,这可能是抗病材料PAL酶活性上升幅度大,大量积累了木质素,并迅速合成酚类物质和植保素,增加细胞壁的厚度,形成病原入侵的机械屏障,从而表现出来抗性,而感病材料PAL活性上升速度慢,没有迅速形成机械屏障阻碍病原菌的侵入表现为感病。接种白粉病菌后,花溪辣椒(抗病)和遵义辣椒(感病)的几丁质酶和β-1,3-葡聚糖酶活性迅速上升,而西安8819(感病)的几丁质酶和β-1,3-葡聚糖酶活性上升速度较慢,这可能是由于辣椒品种不同而出现的差异,似乎这2种酶活性与辣椒的抗病性无相关性,与许多学者的研究结果不一样,其原因需要进一步研究。
组织病理学研究发现,辣椒叶片接种白粉病菌后24 h,菌丝周边的蜡质层被溶解,笔者认为这是菌丝分泌角质酶迅速降解辣椒叶片的角质,降低其防御能力,以达到侵害的目的。扫描电子显微镜和荧光显微镜观察均未发现菌丝从气孔侵入,且菌丝从气孔旁穿行而过,未见有从气孔侵入的迹象。其侵入方式有2种可能性,一是孢子产生附着孢后,附着孢产生吸器或侵染菌丝直接侵入植物组织;二是孢子产生附着孢后,并不产生吸器或侵染菌丝,而是附着孢在组织表面产生菌丝,以菌丝的侧枝侵入到植物组织里面。这需要以后的透射电镜的观察确定。
参考文献:
[1]白滨,胡冠芳. 甘肃省辣椒病害新记录——辣椒白粉病[J]. 甘肃农业科技,2002(7):46.
[2]莫熙礼,蒋选利,彭赫. 10%世高水分散粒剂防治辣椒白粉病药效试验[J]. 山地农业生物学报,2007,26(1):39-42.
[3]李靖,利容千,袁文静. 黄瓜感染霜霉病菌叶片中一些酶活性的变化[J]. 植物病理学报,1991,21(4):278-283.
[4]薛应龙,欧阳光察,澳绍根. 植物苯丙氨酸解氨酶的研究Ⅳ. 水稻幼苗中PAL活性的动态变化[J]. 植物生理学报,1983,9(3):301-305.
[5]车海彦,吴云锋,杨英,等. 植物源病毒抑制剂WCT-Ⅱ控制烟草花叶病毒(TMV)的作用机理初探[J]. 西北农业学报,2004,13(4):45-49.
[6]李保聚,李凤云. 黄瓜不同抗性品种感染黑星病菌后过氧化物酶和多酚氧化酶的变化[J]. 中国农业科学,1998,31(1):86-88.
[7]陈鹏,李振岐. BTH对小麦叶片防卫相关酶的系统诱导作用[J]. 西北植物学报,2006,26(12):2468-2472.
[8]汤章城. 现代植物生理学实验指南[M]. 北京:科学出版社,1999.
[9]刘会宁,姚晓雯. 葡萄霜霉病抗性与叶片生理生化指标的关系[J]. 江苏农业科学,2015,43(11):180-182.
[10]王雅雅,付瑞敏,于烽,等. 植物土传疾病拮抗菌的筛选及其生防潜力评价[J]. 江苏农业科学,2015,43(7):102-106.
[11]Sekizawa Y,Harayawa T,Kano M,et al. Dependence on ethylene of induction of peroxidase and lipoxygenase activity in rice leaf infecled with blast fungus[J]. Agric Bio Chem,1990,54:471-478.
