不同海拔高度对星油藤叶片特性、植株生长及种子成分的影响
2016-05-30焦冬英杨春蔡传涛蔡志全
焦冬英 杨春 蔡传涛 蔡志全


摘 要 为了探讨不同海拔高度上星油藤植株生长和产量构成的差异,在西双版纳雨季,对4个海拔梯度(560、900、1 200、1 490 m)上栽培的星油藤(Plukenetia volubilis)进行了叶片光合与解剖、植株生长与种子成分的观测。结果表明:不同海拔高度的星油藤叶片的最大净光合速率差异不显著;高海拔(≥1 200 m)叶片的气孔导度和呼吸速率值显著小于低海拔(<1 200 m),但水分利用效率较高。叶片上下表皮的气孔密度随海拔升高均显著减小,而叶片厚度和气孔大小的变化则不显著。单株的总生物量、果实生物量、地上部生物量、地下部生物量和叶面积指数均随海拔的升高而减小,比叶面积和比根长则随海拔的升高而增大。低海拔(560 m)星油藤具有较大的根茎叶非结构性碳水化合物(NSC)含量以及氮和NSC库,较高海拔(900~1490 m)植株则存在不同程度的碳源供应不足。不同海拔高度星油藤种子的NSC、脂肪酸、蛋白质和多酚含量差异不显著。试验结果表明,不同海拔高度星油藤生物量的差异主要由植株冠层碳源供应不足造成;雨季种子产量随海拔升高而降低,但种子的品质没有显著差异。
关键词 星油藤;海拔高度;生长;非结构性碳水化合物;种子成分
中图分类号 S727.3 文献标识码 A
Leaf Traits,Plant Growth and Seed Chemicals of Plukenetia
volubilis Cultivated Along an Altitude Gradient
JIAO Dongying1,2, YANG Chun1, CAI Chuantao1, CAI Zhiquan1 *
1 Key Laboratory of Tropical Plant Resources and Sustainable Use, Xishuangbanna Tropical Botanical Garden,
Chinese Academy of Sciences, Mengla, Yunnan 666303, China
2 Kunming Inca Biotechnology Co., Ltd. Kunming, Yunnan 650220, China
Abstract Leaf photosynthesis, anatomical structure, plant growth and seed chemical compositions in the rainy season were measured in Plukenetia volubilis cultivated along an altitude gradients(560, 900, 1 200 and 1 490 m)in Xishuangbanna, in order to unravel the differences of P. volubilis plant's growth and the yield component between different altitudes. The results showed that the maximum net photosynthetic rate had no difference between the four altitudes, while the stomatal conductance and respiratory rate were significantly lower, and water use efficiency were higher in high altitudes(≥1 200 m)than those in low altitudes(<1 200 m). With an increase of altitude, the stomatal density decreased significantly, while stomata size had no difference in both adaxial and abaxial leaves. The total biomass, fruit biomass, aboveground vegetative biomass, underground vegetative biomass and leaf area index of plants decreased, while the specific leaf area and specific root length increased with the increasing of elevation. In 560 m, non-structural carbohydrates(NSC)content, nitrogen pool and NSC pool in roots, stems and leaves were the highest, which indicated that the plants in high altitudes(900-1 490 m)showed the growth limitation due to carbon source deficiency. No significant differences were observed in the content of seed chemical compositions(NSC, fatty acids, protein and polyphenol)between the four elevations. It is concluded that biomass and yield difference of P. volubilis along an altitude gradient was mainly attributed to carbon source deficiency. Seed chemicals of P. volubilis are relatively stable in the rainy season.
Key words Plukenetia volubilis; Altitude; Growth; Non-structural carbohydrates; Seed chemical composition
doi 10.3969/j.issn.1000-2561.2016.02.024
星油藤(Plukenetia volubilis Linneo)为大戟科(Euphorbiaceae)攀援状多年生木质藤本,原生长在海拔80~1 700 m的南美洲安第斯山脉热带雨林中,2006年引种至版纳植物园并获成功。星油藤种子富含油脂、蛋白质、氨基酸,还有多种维生素和生育酚[1-3]。油脂部分主要由多种不饱和脂肪酸组成,其多不饱和脂肪酸含量明显高于其它油料植物[1-2]。
目前国外对星油藤的研究较少,主要集中在种子油脂方面,关于栽培方面的研究则更少。国内对星油藤的研究也仅处于起步阶段[2-4]。星油藤在西双版纳引种和推广多年以来,长势良好,产量可观。将不同海拔高度的特定环境作为一个有机整体,研究环境综合效应对星油藤的影响,不仅可充分利用云南大量的荒山资源,也可改变单一橡胶种植现状,因此,对星油藤山地栽培开展研究对促进热带地区的经济发展和完善星油藤的产业化开发具有重要意义。
植物组织的非结构性碳水化合物(NSC),即淀粉与可溶性糖的总和,是植物碳吸收(光合同化)与碳消耗(生长与呼吸消耗)关系的一种量度,反映了可供植物生长利用的物质水平;NSC在很大程度上影响着植物的生长发育[5]。一般认为,随海拔升高NSC的积累或不足都会引起植物生长受阻,并表现为植株生物量减小。随着海拔升高土壤温度的降低导致根系生长减慢,细根作为土壤中碳库的主要来源,吸收水分和养分的能力更多地取决于比根长[6]。高海拔植株细根的主要形态指标往往大于低海拔,因此具有较强的吸收能力[7]。随海拔升高,植株生物量减小,并增加了向地下部分的生物量分配[8],导致植株地上部分的生产能力减小,最终引起了产量的下降。本研究旨在弄清星油藤在不同海拔高度的生理形态变化、产量和品质的差异以及高海拔星油藤生长受限的原因。
1 材料与方法
1.1 研究地概况
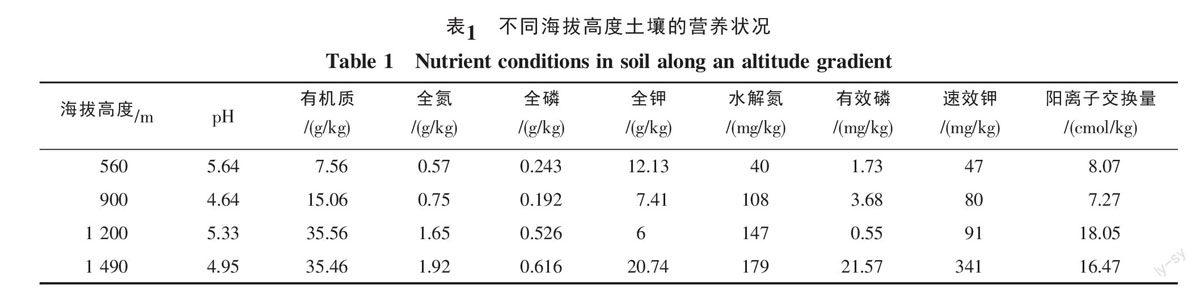
试验设置在西双版纳地区的4个海拔点:(1)中科院西双版纳热带植物园(21°56′N,101°15′E),海拔560 m;(2)勐腊县小勐养四家寨(22°11′N,100°55′E),海拔900 m;(3)勐腊县红冒树(21°48′N,101°19′E),海拔1 200 m;(4)勐腊县象明曼供村(22°13′N,101°20′E),海拔1 490 m。试验区为西南季风控制区,一年中有明显的雨季(5~10月)和干季(11~4月)之分。试验地土壤营养状况见下表(表1),土壤呈酸性,随海拔升高,土壤有机质、有效N、P、K增加(除1 200 m的P较低外),高海拔(≥1 200 m)土壤阳离子交换量也较高,高海拔的土壤环境优于低海拔。
1.2 方法
1.2.1 试验设计 选取中科院西双版纳热带植物园引种栽培的星油藤植株所结成熟完整种子,于2009年6月在温室沙床中萌发育苗。于2009年8月将生长一致的约20 cm高的星油藤实生苗定植于以上4个海拔点,栽种密度为2 m×2 m,种植面积大于667 m2。每株每年施复合肥150 g(N ∶ P2O5 ∶ K2O=1 ∶ 1 ∶ 1),以植株为中心开环状沟,将肥料均匀撒入沟内并覆土,于实验开始时一次性施入。根据植株生长需要及时除草除虫,用水泥桩和铁丝网搭建攀援架支撑星油藤生长。
1.2.2 测定方法 a.星油藤叶片光合、结剖特征的测定:选择植株顶端成熟同龄的完整叶片,于2011年8月份(雨季)晴天上午8 : 30~11 : 30,用Li-6400光合测定系统测定不同海拔梯度上星油藤的光合参数。测定时通过系统来控制空气流量为500 μmol/s,叶室温度为25 ℃~27 ℃,叶室CO2浓度(400±10)μmol/mol,相对湿度75%~80%,采用Li-6400人工光源,测定叶片在2 000 μmol/m2·s光强下的净光合速率(Amax)、气孔导度(gs)和黑暗条件下的呼吸速率(Rd),计算得水分利用效率(WUE=Amax/gs),每一海拔重复6~8株。
与光合测定同步进行叶片的显微结构观察,徒手切片制成水装片,在40×10倍双筒显微镜(Olympus,Japan)下测量上层成熟叶片的总厚度及各组织层厚度。用无色指甲油涂于叶片上下表面取印记制成装片,20×10倍显微镜下分成10个观测点,计数网格尺内的气孔数,将平均值换算成单位面积的气孔数,即得气孔密度;在40×10倍显微镜下测量保卫细胞长度。
b.整株形态特征的测定:于2010年8月中下旬,每一海拔梯度随机选取6株个体,挖回根系完整的全植株。自来水下冲洗掉根部泥土,将植株的粗根(直径≥1 mm)、细根(直径<1 mm)、茎、叶、花(包括雄花和雌花)和果实分开。用Li-3000叶面积仪(Li-Cor,USA)测定叶面积(LA,cm2),再将其置于70 ℃恒温干燥箱中干燥48 h至恒重并称重(LW,g),求取比叶面积(SLA=LA/LW,cm2/g)和;将细根进一步切成4~5 cm长的小段,在400 dpi的分辨率下进行扫描(根系相互不重叠),扫描得到的图片用DT-Scan(WINR HIZO software,Regent Instruments,Quebec,Canada)软件计算细根的根长(RL,m),并求得扫描细根的干重(RW,g),计算比根长(SRL= RL/RW,m/g);将植株其他叶部分、茎、根、花及果实在70 ℃下烘干至恒重并称重,求得各部分生物量。计算求得总生物量、地上部生物量、地下部生物量及叶面积指数(LAI=SLA×ML/A,其中ML为叶生物量,A为植株在地面的垂直投射面积)。
c.种子及营养器官化学成分的测定:采收成熟饱满种子,去壳粉碎;根、茎和叶粉碎过60目筛,每个海拔5个重复。样品送至农业部农产品质量监督检验测试中心(昆明)进行检测,测定项目有:种子粗脂肪、蛋白质及多酚含量,植株叶片、茎、根及种子的N、淀粉、可溶性糖和非结构性碳水化合物(NSC=淀粉+可溶性糖)含量。各营养成分的检测方法分别为:粗脂肪(GB/T 5512-2008,同食品中粗脂肪测定);N/蛋白质(GB/T 5009.5-2010,凯氏定氮法);脂肪酸(GB/T 14489.3-1993,索氏提取法),测试结果为总脂肪酸的相对含量;可溶性糖(NY/T 1278-2007,铜还原碘量法);淀粉(GBT 5009.9-2008,酶水解法);多酚(GB/T5009.82-2003,高效液相色谱法)。
1.3 数据处理与统计分析
采用单因素方差分析(One-way ANOVA)比较不同海拔植株不同参数之间的差异显著性,不同海拔间多重比较选择LSD法,显著水平α=0.05。所有数据采用SPSS17.0进行分析,所有数据在分析前进行方差的齐性和正态性检验,方差不齐时要进行对数或平方根转换。
2 结果与分析
2.1 海拔高度对星油藤叶片光合特性和解剖特征的影响
海拔高度对雨季星油藤最大光合速率(Amax)的影响不显著,说明雨季高海拔植株叶片也具有较强的光合生产能力(表2)。海拔对叶片气孔导度(gs)的影响显著,高海拔(≥1 200 m)植株的gs较低而水分利用效率(WUE)较高。叶片gs和WUE具有极显著的负相关关系(r=-0.938,p<0.001),海拔1 200 m植株具有最小的gs和最大的WUE。叶片呼吸速率(Rd)受海拔影响显著,高海拔(≥1 200 m)植株Rd显著低于低海拔(≤900 m)植株。植株叶片的氮素利用效率PNUE在不同海拔高度差异显著,在最低海拔(900 m)和最高海拔(1 490 m)处较高,与另外2个海拔高度的叶片含氮量关系较大。
海拔高度对星油藤叶片厚度的影响较小(图1-A),高海拔(≥1 200 m)叶片栅栏组织厚度小于低海拔;叶片的上、下表皮及海绵组织厚度变化较小(图1-B)。星油藤叶片的气孔主要分布于下表皮,上、下表皮气孔密度受海拔的影响均显著并随海拔的升高而减小(图1-C、E);叶片上下表皮气孔密度总和在海拔间差异显著(p<0.001。叶片上表皮保卫细胞的长度大于下表皮保卫细胞长度,但海拔对上、下表皮气孔大小的影响不显著(p>0.05)(图1-D、F)。上表皮气孔密度和保卫细胞大小、下表皮气孔密度与保卫细胞大小间均无显著的相关关系(p>0.05)。
2.2 海拔对星油藤非结构性碳水化合物的影响
海拔对星油藤茎叶氮和淀粉的含量以及根茎叶可溶性糖的含量影响显著。最低海拔(560 m)植株有最小的叶氮含量和最大的根淀粉含量,而900~1 490 m的海拔范围内叶氮含量和根淀粉含量的变化则较小(图2-A、B)。叶片的氮含量以及根的淀粉、可溶性糖和NSC含量在海拔梯度上有显著的负相关关系(r=-0.690~-0.737,p<0.001)。根茎叶的可溶性糖与NSC含量具有极显著的正相关关系(r=0.982~0.998,p<0.001),NSC含量在不同海拔间的差异主要由存储形式的可溶性糖造成。低海拔(560 m)植株具有较大的根茎叶可溶性糖含量;海拔1 200 m处,植株根叶可溶性糖含量显著低于其他海拔(图2-C),与该海拔处较低的Amax相一致;海拔1 490 m处,植株具有较高的根叶可溶性糖含量。在最低海拔560 m处,植株各器官中NSC含量均较高;与最低海拔相比,海拔900 m的根、海拔1 200 m的根茎叶及海拔1 490 m的根茎中NSC含量显著降低。在900~1 490 m的海拔范围内,随海拔升高根NSC含量增加,说明植株向根的碳分配增加(图2-A)。海拔对星油藤根茎叶N和NSC库的大小有显著影响,最低海拔560 m处根茎叶的N、NSC库均较大,与之相比,海拔1 200 m的茎叶N库和1 490 m的根茎叶N库显著减小,海拔900 m的根NSC库、海拔1 200 m和1 490 m的根茎叶NSC库也显著减小(图3-A、B)。
2.3 海拔对星油藤植株形态和生长的影响
星油藤的总生物量、果实生物量、地上部生物量、地下部生物量及叶面积指数(LAI)随着海拔的升高显著减小(p<0.05),而比叶面积(SLA)则显著增大(p=0.028);随着海拔升高,比根长(SRL)的增大不显著(表3)。雨季海拔梯度上的星油藤具有较大的养分捕获能力(高的SLA和SRL),与植株高速生长的生理需求相一致(Amax较高)。用果实生物量预测海拔梯度上星油藤的收获指数(果实生物量/总生物量)可以得出,高海拔(1 200 m为0.40;1 490 m为0.40)比低海拔(560 m为0.31;900 m为0.22)的收获指数高,高海拔更利于总生物量向经济产量的转化。
2.4 海拔对星油藤种子化学成分的影响
种子中可溶性总糖随海拔升高而降低,但淀粉、非结构性碳水化合物、脂肪酸、蛋白质和抗氧化活性物质多酚含量在各海拔间差异不显著(表4)。
3 讨论与结论
叶片作为光合作用的重要器官,对环境变化敏感,可塑性大,其结构特征最能体现环境因子的影响和植物对环境的适应,因此叶片的形态解剖特征及功能与环境的关系较为密切[9-10]。叶片厚度、栅栏组织厚度、气孔密度和气孔大小受海拔环境中水分和光照的影响较大[11]。在西双版纳雨季,随海拔升高,降雨量增加而蒸发量减小,空气相对湿度升高[12]。受水分和光辐射的共同作用,高海拔(≥1 200 m)叶片栅栏组织变薄而叶片厚度没有明显变化;上、下表皮气孔密度随着海拔梯度上水分和湿度的增加、空气CO2浓度的降低而减小,与相关研究结果一致[9]。叶片的SLA主要与空气的蒸汽压/潜在蒸散呈正相关关系,干旱环境中植物的SLA较小,SLA随海拔升高递增的趋势可能与具有相同变化趋势的降水量有关[13-14]。低海拔实验地的土壤养分含量并不优于中、高海拔,但高海拔星油藤叶片光合获取碳的能力并不比低海拔植株低。随海拔升高根系SRL的增加有利于提高低温下植株对地下营养和水分的捕获能力。高海拔星油藤具有较强的地下资源捕获能力(SRL较大)和较高的组织营养物质含量(氮含量),因此其叶片的光合能力和根系功能不是高海拔星油藤生长受限的主要原因。
“碳源限制”假说认为环境胁迫使植物的碳吸收与碳消耗关系失调,碳供应不足导致高海拔植物生长受阻;而“生长抑制”假说认为植物碳供应充足,但低温导致充足的碳不能被生长所利用,从而使生长受限[15-16]。植物组织中NSC还可以帮助植物抵御荫蔽、干旱、低温等逆境环境[17-18]。有研究认为植物保持一定的NSC含量而不会因库的需求将其消耗殆尽,NSC不是油橄榄产量的决定因素而是一种生存策略[19]。本试验中,与最低海拔560 m相比,海拔900~1 200 m的植株存在不同程度的NSC供应不足。雨季海拔梯度上温度较高,随海拔升高温度的减低不足以对植株造成逆境伤害,NSC可能不是植物响应环境逆境而产生的信号物质。在高海拔处星油藤植株的根茎叶NSC含量较低,说明冠层碳的获取能力不能满足植株营养生长的需求,雨季星油藤的生长受到碳源供应不足的限制。海拔梯度上环境因子的差异(如温度、降雨等)没有对星油藤的叶片光合能力产生显著影响。但是随着海拔升高,叶面积的减小使得植株冠层碳获取能力减小,最终导致星油藤生长受限。
环境条件直接影响着光合同化产物向种子中不同储藏物(油脂、蛋白质等)的转化和分配;不同油料作物的种子成分与环境的水分、温度及营养条件等因素密切相关。温度对油料种子的影响因种而异,油菜、大豆的种子含油量和环境温度之间无相关性,而向日葵种子的油含量与温度间存在极显著的负相关关系[20]。海拔作为影响植物生长的综合性环境因子,对不同的油料作物含油量的影响不同。高海拔地区的油菜和油橄榄的种子含油量比低海拔的含油率要高[21-22]。沙棘果在一定的海拔高度下含油量较高,过高或过低的海拔均会影响其含油量[23]。然而,当海拔梯度上环境条件的差异没有对油料植物的种子形成产生限制时,油料植物种子的化学成分差异会较小,如Lesquerella fendleri的种子油含量在300~1 200 m的海拔梯度上相似,表明温度引起的海拔环境变化对油含量的影响较小[24]。雨季星油藤种子的化学成分在海拔间差异不显著,可能与雨季海拔梯度上充足的水分和土壤养分条件以及热带地区较高的温度等适宜生长的条件有关。
参考文献
[1] Hamaker B R, Valles C, Gilman R, et al. Amino acid and fatty acid profiles of the Inchi peanut(Plukenetia volubilis L.)[J]. Cereal Chemistry, 1992, 69: 461-463.
[2] 蔡志全, 杨 清, 唐寿贤, 等. 木本油料作物南美油藤种子营养价值的评价[J]. 营养学报, 2011, 33(2): 193-195.
[3] 蔡志全. 特种木本油料作物星油藤的研究进展[J]. 中国油脂, 2011, 36(10): 1-6.
[4] Cai Z Q. Shade delayed flowering and decreased photosynthesis, growth and yield of Sacha Inchi(Plukenetia volubilis)plants[J]. Industrial Crops and Products, 2011, 34: 1 235-1 237.
[5] Li M H, Xiao W F, Wang S G. Mobile carbohydrates in Himalayan treeline trees I. Evidence for carbon gain limitation but not for growth limitation[J]. Tree Physiology, 2008, 28(8): 1 287-1 296.
[6] Eissenstat D M. Costs and benefits of construction roots of small diameter[J]. Journal of Plant Nutrition, 1992, 15(6-7): 763-782.
[7] 权 伟, 徐 侠, 王 丰, 等. 武夷山不同海拔高度植被细根生物量及形态特征[J]. 生态学杂志, 2008, 27(7): 1 095-1 103.
[8] Moser G, Leuschner C, Hertel D, et al. Elevation effects on the carbon budget of tropical mountain forests(Ecuador S): The role of the belowground compartment[J]. Global Change Biology, 2011, 17(6): 2 211-2 226.
[9] 李芳兰, 包维楷. 植物叶片形态解剖结构对环境变化的响应与适应[J]. 植物学通报, 2005, 22(增刊): 118-127.
[10] 胡 营, 楚海家, 李建强. 4个花苜蓿居群叶片解剖结构特征及其可塑性对不同水分处理的响应[J]. 植物科学学报, 2011, 29(2): 218-221.
[11] 尤 毅, 孙映波, 吕复兵, 等. 干旱胁迫对文心兰生长及光合特性的影响[J]. 热带作物学报, 2011, 32(7): 1 245-1 248.
[12] 郑 征, 李佑荣, 张树斌, 等. 西双版纳海拔变化对水湿状况的影响[J]. 山地学报, 2007, 25(1): 33-38.
[13] Wright I J, Reich P B, Cornelissen J H C, et al. Modulation of leaf economic traits and trait relationships by climate[J]. Global Ecology and Biogeography, 2005, 14: 411-421.
[14] Meziane D, Shipley B. Direct and indirect relationships between specific leaf area, leaf nitrogen and leaf gas exchange. Effects of irradiance and nutrient supply[J]. Annuals of Botany, 2001, 88(5): 915-927.
[15] Hoch G, Popp M, Korner P C. Atitudinal increase of mobile carbon pools in pinus cembra suggests sink limitation of growth at the Swiss treeline[J]. Oikos, 2002, 98(3): 361-374.
[16] Korner C. Alpine plant life: functional plant ecology of high mountain ecosystems[M]. 2nd ed. New York: Springer Berlin Heidel-berg, 2003, 337.
[17] Li M H, Hoch G, Korner C H. Source/sink removal affects mobile carbohydrates in Pinus cembra at the Swiss treeline[J]. Trees, 2002, 16(4-5): 331-337.
[18] 潘红丽, 李迈和, 蔡小虎, 等. 海拔梯度上的植物生长与生理生态特性[J]. 生态环境学报, 2009, 18(2): 722-730.
[19] Bustan A, Avni A, Lavee S, et al. Role of carbohydrate reserves in yield production of intensively cultivated oil olive(Olea europaea L.)trees[J]. Tree physiology, 2011, 31(5): 519-530.
[20] 宁海龙, 宋秀吉, 王雪依, 等. 氮磷钾肥对大豆脂肪含量的效应[J]. 中国油料作物学报, 2007, 29(3): 302-307.
[21] 付三雄, 伍晓明, 李成磊, 等. 不同地理位置对甘蓝型油菜含油量的效应研究[J]. 江苏农业学报, 2009, 25(2): 247-252.
[22] Dabbou S, Sifi S, Rjiba I, et al. Effect of pedoclimatic conditions on the chemical composition of the sigoise olive cultivar[J]. Chemistry & Biodiversity, 2010, 7(4): 898-908.
[23] 阮成江, 李代琼. 沙棘含油量及影响因子研究综述[J]. 西北植物学报, 2001, 21(2): 207-214.
[24] Dieri D A, Adama N R, Mackey B E, et al. Temperature and elevation effects on plant growth, development, and seed production of two Lesquerella species[J]. Industrial Crops and Products, 2006, 24: 17-25.
