花期低温对晚稻干物质积累及分配的影响*
2016-05-27白光志刘寿东余焰文杨再强殷建敏南京信息工程大学气象灾害预报预警与评估协同创新中心南京0044江西气候中心南昌330000
白光志,刘寿东,余焰文,杨再强**,殷建敏(. 南京信息工程大学气象灾害预报预警与评估协同创新中心,南京 0044;. 江西气候中心,南昌330000)
花期低温对晚稻干物质积累及分配的影响*
白光志1,刘寿东1,余焰文1,杨再强1**,殷建敏2
(1. 南京信息工程大学气象灾害预报预警与评估协同创新中心,南京 210044;2. 江西气候中心,南昌330000)
摘要:以“广两优香66”为试材, 于2014年8-10月在南京信息工程大学农业气象站进行人工控制实验, 研究花期低温对晚稻光合作用和后期灌浆过程的影响, 并分析低温胁迫下晚稻后期干物质积累和分配的变化。低温胁迫温度设定为22℃ /17℃(昼/夜), 处理时间分别为2、4、6和8d, 分别依次记为T2d、T4d、T6d和T8d, 以外界环境条件32℃ /25℃ (昼/夜)为对照(CK)。结果表明:(1)低温处理后, T2d处理最大光合速率(Pmax)变化较小, T4d和T6d分别比CK下降11.66%和21.47%, T8d的Pmax仅为17.25μmol·m−2·s−1, 比CK减少28.54%。低温处理后叶片光饱和点(LSP)降低, 而光补偿点(LCP)明显升高, 与CK相比, T2d、T4d、T6d、T8d的LSP分别降低6.68%、8.08%、8.63%、10.87%, LCP升高2、3、4、6倍。说明低温处理降低晚稻叶片的光合能力;(2)对不同处理的灌浆过程的拟合表明, 低温处理时间越长, 达到最大灌浆速率所需时间(T-GRmax)越长, 晚稻最大千粒重(A)越小, 灌浆活跃期的时间(T90)也越短, 导致整体平均灌浆速率(GR)呈不同程度下降, 说明低温会减缓晚稻的灌浆过程;(3)低温处理时间越长, 晚稻结实率越低, 不超过2d低温对结实率无显著影响, 但4d及以上低温处理能显著降低晚稻结实率, T8d处理的结实率比CK降低34.39%;(4)晚稻总干物质与花期低温处理时间呈负相关, 且低温处理6d后总干物质显著减少, T8d处理总干物质比CK减少28.05%, 其中叶片和穗部减少较明显, 低温处理4d后两者干物质均明显下降, 处理8d后比CK分别减少42.57%和38.70%。研究表明花期低温处理抑制晚稻光合作用, 4d及以上低温处理显著降低结实率, 减缓籽粒灌浆过程, 对产量形成不利。
关键词:花期低温;晚稻;光合作用;灌浆;干物质
白光志,刘寿东,余焰文,等.花期低温对晚稻干物质积累及分配的影响[J].中国农业气象,2016,37(2):206-212
水稻的低温冷害在许多国家均有发生,是全球性自然灾害。全球有1500万hm2以上的稻作面积受到低温威胁,尤以日本、朝鲜和中国为甚[1]。据Deng 等[2]研究报道,全球每年因低温伤害造成的农、林业损失高达数千亿,Bertrand等[3]指出随着全球气候变暖,极端气候尤其是低温将更频发,这一损失甚至还有可能增加。
Lyons[4]研究表明,水稻在温度低于10~12℃时即表现出冷害症状。关于冷害对植物造成危害的原因,目前较普遍接受的观点是0℃以上低温引起了生物膜系统的损害[5],自由基产生和清除机制失衡[6-8],叶绿体超微结构破坏[9-10],叶绿素合成受抑制,导致光合能力下降[11]。低温不仅降低在叶绿体基质中进行的光合作用暗反应活性[8],也引起类囊体膜介导的光反应活性降低[12],使类囊体膜上的PSⅡ的光能传递效率和光能转换效率降低[13],从而导致CO2同化能力降低。据Kaniuga等[14]报道,低温使叶绿体的希尔反应活性降低,使类囊体膜中PSⅡ的电子传递活性受抑,也会降低植物的光合能力。
颖花分化期低温不仅显著降低早稻叶片气体交换能力,明显降低穗粒数、结实率、千粒重和最终产量[15]。抽穗开花期遭遇低温后,花粉育性受到影响,子房延长受到阻碍,空粒显著增加[16],可导致结实率降低10%~20%,严重时可导致晚稻大幅减产[17]。邱上深[18]通过实验证明,水稻花期和灌浆期的冷害机制主要是因低温影响同化物的形成和运输,进而影响劣势位粒胚乳的分化和灌浆,致使部分子粒停止灌浆变为秕谷。低温下,作物的多种生理活动均会有不同程度的下降,在宏观上表现为延缓生长发育,进而导致生育期延长,此外还表现为作物生长量降低,干物质积累减少,群体生产力下降。夜间不同时段低温处理明显降低作物植株地上部分干物质积累总量,并导致叶片中糖和淀粉的积累,夜低温处理使根中的干物质分配比率前期减少而后期增加,叶、茎的干物质分配比率增加,果实的干物质分配比率降低[19-20]。水稻产量形成的过程实质上是干物质积累与分配的过程,但关于晚稻干物质积累和分配对花期低温的响应研究报道并不多见,本文通过研究花期不同低温处理晚稻后期干物质积累和分配的特征,以期为晚稻后期低温灾害研究和指标制定提供一定科学依据。
1 材料与方法
1.1试验设计
试验在南京信息工程大学农业试验站进行,以南京晚稻品种“广两优香66”为试材,2014年4月30日播种,5月30日将秧苗(五叶)分别移栽至30个塑料盆中,盆上口为边长35cm 的正方形,盆底为边长30cm 的正方形,盆深40cm。每个处理6盆,3盆用于测定灌浆速率,3盆用于测定成熟期干物质分配情况。每盆栽3株秧苗,置于田间保持良好生长,待水稻开始进入抽穗扬花期(8月27日)挂标识牌定穗,并取其中的24盆置于人工气候箱(TPG-1260, Australian)中进行低温处理,参照气象行业标准《寒露风等级》[21]气候箱设定白天(6:00-18:00 )和夜间(18:00-6:00)温度分别为22℃和17℃,为避免临界时间对试验的影响,低温处理时间设定为2、4、6和8d(分别记为T2d、T4d、T6d和T8d),处理结束后即取出置于室外。剩余6盆一直置于室外自然环境中,作为对照(CK)处理,处理期间昼/夜平均温度为32℃/25℃。期间需关注室外温度情况,若温度低于处理温度则需将室外水稻移至室内,避免自然低温对试验结果的影响,并记录试验期间各处理晚稻从灌浆至成熟所经历的时间天数。
1.2测定项目
1.2.1光响应曲线测定
每个处理结束后的第一天9:00-11:00 测定水稻叶片光合速率,选取上3叶(剑叶、倒1叶和倒2叶)进行测定,采用便携式光合作用测定系统(Li-6400,USA)测定水稻叶片净光合速率(Pn)。设定参比室CO2浓度为400μL·L−1,叶室温度为25℃,按照由强到弱的顺序手动设定光辐射强度(PAR)分别为2000、1800、1600、1400、1200、1000、800、600、400、200、150、100、50、0μmol·m−2·s−1。每叶1次,共3次,即为3次重复。
1.2.2灌浆速率测定
每5d取一次挂标识牌定穗的稻穗,每次3穗,取下籽粒后,数其总粒数后放入铝盒,在恒温干燥箱中,60~80℃烘烤24h以上(幼小籽粒烘烤温度宜低),使用精度为0.001g电子天平称取籽粒干重,直至两次重量相差不超过0.01g,再转换成千粒重。
1.2.3干物质分配
将不同处理组的水稻按成熟先后依次收获并分解为根、茎、叶和穗洗净,分别在烘箱中烘干至恒重(前后两次相差不超过0.01g)后测定干物质量,然后计算植株各器官干物质分配比例,并统计各处理组的结实率。
1.3统计分析
试验数据运用DPS、SPSS进行相关统计分析,Excel作图,使用Photosynthesis Work Bench程序进行光响应曲线拟合,使用Matlab对晚稻灌浆速率进行拟合,拟合方程为Richard模型,其表达式为

式中,t为晚稻开花后天数(d),y为对应时间t的晚稻千粒重,B为初值参数,K为生长速率参数,A为生长终止时的最大千粒重(g),N为环境充分系数,决定曲线形状[22]。通过式(1)对t求导得到灌浆速率GR的表达式为

当t=0时,可求出初始灌浆速率GR0和初始相对灌浆速率即起始势RGR0= GR0/y0,式(2)再对t求导,可得到灌浆速率的变化率GR′,由此可再求出灌浆持续天数T90、平均灌浆速率GR等,详细参数说明及来源参考文献[23-24]。
2 结果与分析
2.1花期低温对晚稻叶片光响应曲线的影响
由图1可见,与CK相比,低温处理后晚稻叶片对光能的利用有不同程度的下降,相同光照强度下,随着低温处理时间的延长,晚稻叶片的净光合速率下降,且下降幅度与低温胁迫时间呈正相关。对各处理光响应曲线拟合后的特征值进行对比可见,CK的最大净光合速率Pmax相对最高,为24.15μmol·m−2·s−1,随着低温处理时间的增加,Pmax持续减小,T2d相对CK变化较小,T4d和T6d分别比CK下降11.66%和21.47%,T8d的Pmax仅为17.25μmol·m−2·s−1,比CK减少了28.54%。不同处理下晚稻叶片光饱和点LSP的变化与Pmax的变化趋势类似,低温处理8d的LSP最低,仅1280μmol·m−2·s−1,比CK的1436μmol·m−2·s−1下降了约10%。不同处理下晚稻光补偿点LCP的变化趋势则与LSP相反,以CK为最低,仅4μmol·m−2·s−1,低温处理后LCP有明显升高,T2d、T4d、T6d、T8d 的LSP分别较CK升高2、3、4、6倍。

图1 各处理叶片光响应曲线Fig. 1 Light response curves of rice leave under different treatments
2.2花期低温对晚稻灌浆速率的影响
对不同处理晚稻灌浆速率进行方程拟合,结果见表1。由表可见,各处理方程的决定系数均超过0.98,说明拟合结果较可靠。最大千粒重A随花期低温处理时间的增加呈不同程度下降,且低温处理时间越长下降幅度越大,CK的A值最大,为29.311g,低温处理2d后减小了16.89%。低温处理4d后A值为22.022g,低温处理6d和8d的A值有明显下降,分别仅为CK的64.59%和53.88%。
在确定方程参数基础上进一步确定灌浆过程的相关特征参数,结果见表2。由表可见,初始灌浆速率GR0随着低温处理时间的增加呈持续下降趋势,T2d、T4d、T6d和T8d各处理GR0相对CK分别减小了19.26%、47.31%、59.87%和69.01%,说明花期低温处理对晚稻初始灌浆速率有一定的抑制作用,且低温处理时间越长,抑制作用越明显。不同处理下晚稻的初始相对灌浆速率RGR0和最大灌浆速率GRmax则均呈先增加后减小的趋势,低温处理2d,两者相对CK均有一定上升,随着低温时间的增加,则又开始有不同程度的下降,其中GRmax的变化幅度相对较小,RGR0下降则更明显,T8d处理下RGR0比CK减少了近50%。最大灌浆速率对应的花后天数T-GRmax和此时的千粒重y-GRmax两者对花期低温响应的整体趋势大致相似,短期(2d)低温处理,两者有不同程度减小,低温处理时间超过2d,则均随低温处理时间增加而增加,且T-GRmax对低温的响应更明显,长时间花期低温处理后晚稻灌浆速率最大时的时间明显推迟,T8d处理相对CK更是晚了近12d,但此时千粒重减少却不多。此外,花期低温也会明显缩短晚稻的灌浆活跃期T90,各低温处理组灌浆活跃期相对CK缩短5~12d,且低温处理时间和T90的下降幅度呈正相关。

表1 不同处理下Richard方程的参数结果Table 1 The simulated results of Richard equation parameters for rice grouting process under different treatments
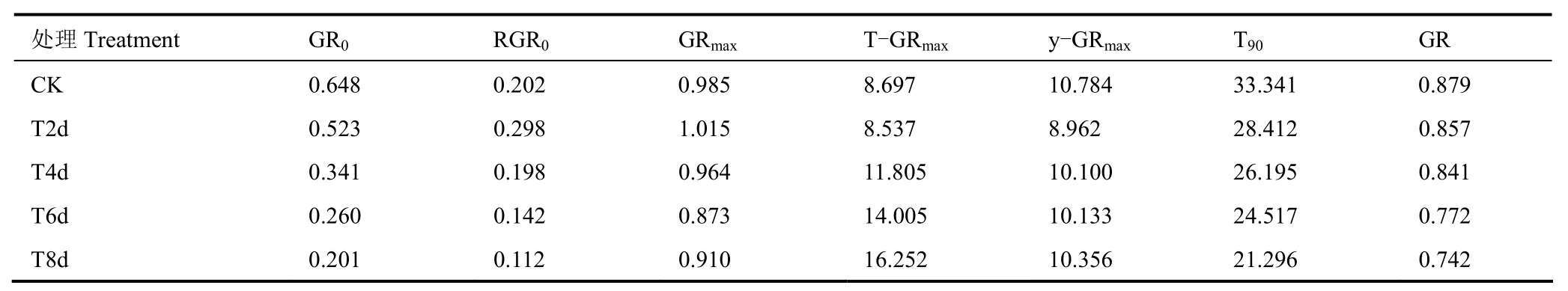
表2 不同处理晚稻灌浆速率的特征参数Table 2 Characteristic parameters of rice filling rate under different treatments
2.3花期低温对晚稻结实率和干物质分配的影响

表3 不同处理下晚稻的结实率(%)Table 3 Seed setting of rice under different treatments(%)
不同低温处理下晚稻的结实率见表3,由表可见,随着低温处理时间的延长,晚稻结实率有不同程度的降低,低温处理时间越长,结实率越低,CK结实率最高,为86.02%,低温处理8d后结实率最低,仅56.44%,相对CK减少了34.39%。各低温处理组间结实率均差异显著,但T2d和CK差异不显著。
由图2a可见,短期(不超过4d)低温处理晚稻干物质有不同幅度下降,但与CK相比并不显著,而低温胁迫时间超过(含)6d后则总干物质明显减少,T6d和T8d总干物质分别为170.43和147.13g/株,比CK分别减少16.65%和28.05%,说明较长时间的低温胁迫处理对晚稻干物质积累的影响显著。不同低温处理时间下晚稻成熟后根系的干重有所增加,但是各处理间差异均不显著,低温胁迫时间不超过8d处理下晚稻茎秆干重减少不明显,低温胁迫8d后则会显著减少。晚稻叶片和穗的干重对花期低温的响应相似,低温胁迫4d后叶和穗的干重显著降低,低温胁迫8d处理叶和穗的干重最小,分别仅为21.06 和54.96g,比CK分别减少42.57%和38.70%。
晚稻各器官干物质所占比例的变化见图2b,由图可见,各处理组间茎秆所占的比例无显著变化,花期低温胁迫不超过6d,晚稻叶片干物质占总干物质的比例无明显变化,而低温胁迫8d后叶片占干物质的比例明显减少,穗占干物质比例在各器官中最高,其变化规律与叶片基本一致。各处理间根系的干物质变化虽然不显著,但其所占总干物质的比例却有较明显的差异。随着低温胁迫时间的增加,根系干物质所占比例也增加,胁迫时间超过2d后与CK相比增加显著。

图3 不同处理晚稻成熟时单株各器官干物质重量(a)及其比例(b)Fig. 3 Organs dry weight(a) and its percentage(b) at rice maturity stage under different treatments

3 结论与讨论
光合作用是作物最重要的生理过程之一,是植物生长发育的基础,是植物合成有机物质和获得能量的根本来源,它可以反映植株生长势和抗性强弱[25],同时其受低温的影响也最明显。低温对作物光合作用的影响几乎贯穿整个光合过程的各个环节,包括光电子的吸收和转运、气体交换、碳同化过程等。杨猛等[26]以玉米为实验材料发现随着低温胁迫处理时间的延长和程度的加强,光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、PSⅡ最大光能转换效率(Fv/Fm)和光系统PSⅡ的潜在活性(Fv/Fo)呈下降趋势,说明低温胁迫使叶片光系统PSⅡ活性中心受损,光合作用效率也因此降低,从而表现出光合速率下降趋势。本实验也得出类似结论,随着低温胁迫时间的增加,水稻叶片最大光合速率和光饱和点下降,光补偿点和暗呼吸速率升高,说明低温对光合作用的抑制作用明显。李平等[27]研究发现,水稻后期受低温影响后表观量子效率和最大光合速率均降低,而FBPase活性也在低温影响下有一定的降低。FBPase对光合作用的运转和光合产物转化以及运输有非常重要的作用,低温作用下水稻光合效率和FBPase活性均下降,光合产物运输受阻,导致源库失衡,这可能是导致水稻减产的主要原因之一。虽然低温处理时间的不同导致各处理间测定叶片的叶龄不完全一致,但测定时仍处于晚稻抽穗扬花期,叶龄差异相对低温对光合作用的影响相对较小。结实率是水稻产量重要的要素之一,许多研究对后期低温处理后水稻结实率降低已形成共识[28-29],主要原因是低温使花粉发育延迟,花药开裂不畅,花粉萌发率下降,影响水稻受精结实[16,30],这也是低温降低水稻产量的重要原因之一。
灌浆过程是光合产物向籽粒运输和积累的过程,水稻籽粒灌浆状况最终决定了籽粒的产量和稻米的质量。本文通过Richard方程对水稻的灌浆过程进行拟合,决定系数均超过0.98,说明拟合的结果可信度较高,再通过拟合方程的参数,根据灌浆过程的生物学意义得出灌浆过程的相关参数,结果表明,低温作用下晚稻的初始灌浆速率和灌浆活跃期均随着低温胁迫时间增加有明显下降,平均灌浆速率也呈现相同规律,而灌浆速率最大时对应的花后天数对低温胁迫时间的响应则表现为增加的趋势。初始相对灌浆速率对低温胁迫时间的响应则表现出先增大后减小的趋势,这可能是晚稻对低温逆境适应性的一种表现。说明低温胁迫影响下,晚稻整体的灌浆速率和灌浆时间均有所缩小,最终导致千粒重下降,产量降低。徐田军等[31]的研究也得出类似结论。
光合产物向各个器官的运输与分配直接关系到植物体的生长和产量的形成,分析晚稻后期的干物质分配特点可以更好地研究光合产物运输过程的源库平衡关系。试验发现,晚稻的总干重随着低温胁迫时间的增加呈不同程度的减小,且低温胁迫6d后减小显著,茎、叶和穗对低温的响应也表现出类似规律,叶和穗的干重在低温胁迫4d后明显下降,而茎秆的干重则在低温胁迫8d后显著减少,根系的干重随着低温胁迫时间的增加有所增加,但是并不显著。综合分析,本研究认为,短期低温处理下(不超过2d),低温主要影响水稻光合作用,但是恢复正常温度后光合作用能达到正常水平,这是T2d与CK相比,最终各器官干物质无显著差异的原因。低温处理超过4d则会减缓花粉发育,使花药不能正常开裂,影响水稻受精,导致结实率降低,此外还会引起库源失衡,籽粒饱满程度降低,最大千粒重减小,灌浆活跃期缩短,减缓籽粒的灌浆过程,这可能是花期低温导致水稻产量降低的主要原因。
本研究旨在分析花期低温对晚稻干物质积累和分配的影响,为晚稻的低温灾害研究提供一定理论依据,也可为水稻的低温防御提供理论支撑,但供试品种仅选用一个,其结果对其它水稻品种是否试用有待更多的试验考究。
参考文献References
[1]钟克友,唐湘如,胡飞,等.水稻剑叶气孔性状与孕穗期耐冷性的关系研究[J].西北植物学报,2010, 30(1):98-103.
Zhong K Y,Tang X R,Hu F,et al.Correlations between the cold tolerance during the booting stage and the stomatal traits of flag leaf surface in rice[J].Acta Bot.Boreal.-Occident.Sin, 2010,30(1):98-103.(in Chinese)
[2]Deng J.Advances of studies on plant freezing tolerance mechanism:freezing tolerance gene expression and its function[J].Chinese Bulletin of Botany,2001,18(5):521-530.
[3]Bertrand A,Castonguay Y.Plant adaptations to overwintering stresses and implications of climate change[J].Canadian Journal of Botanical,2003,81:1145-1152.
[4]Lyons J M.Chilling injury in plants[J].Ann Rev Plant Physiol,1973,24(6):445- 451.
[5]Levitt J.Responses of plants to environmental stresses[S].New York:Academic Press,1980: 23-64.
[6]曾乃燕,何军贤,赵文,等.低温胁迫期间水稻光合膜色素与蛋白水平的变化[J].西北植物学报,2000,20(1): 8-14.
Zeng N Y, He J X, Zhao W, et al.Changes in components of pigments and proteins in rice photosynthetic membrane during chilling stress[J].Acta Bot.Boreal.-Occident. Sin,2000, 20(1):8-14.(in Chinese)
[7]陈善娜,邹晓菊,梁斌.水稻不同抗冷品种幼苗叶细胞膜系统的电镜观察(简报)[J].植物生理学通讯,1997,33(3):191-194.
Chen S N,Zou X J,Liang B.Electronmicroscope observation on membrane system of leave cells of some varieties of rice seedlings with different cold-resistance[J].Plant Physiology Communications,1997,33(3):191-194.(in Chinese)
[8]Weeden N F,Buchanan B B.Leaf cytosolic fructose-1, 6-biphosphatase-A potential targets in low temperature stress[J].Plant Physiol,1983,2(1):259-261.
[9]王静,张成军,陈国祥,等.低温对灌浆期水稻剑叶光合色素和类囊体膜脂肪酸的影响[J].中国水稻科学,2006,20(2):177-182.
Wang J,Zhang C J,Chen G X,et al.Effect of low temperature on photosynthetic pigments and thylakoid membrane fatty acid in flag leaves of rice at the milky stage[J].Chinese J Rice Sci,2006,20(2):177-182.(in Chinese)
[10]Kratsch H A,Wise R R.The ultrastructure of chilling stress[J].Plant Cell and Environment,2000,23(4): 337-350.
[11]王国莉,郭振飞.低温对水稻不同耐性品种幼苗光和速率和叶绿素荧光参数的影响[J].中国水稻科学,2005,19(4):381-383.
Wang G L,Guo Z F.Effects of chilling stress on photosynthetic rate and the parameters of chlorophyll fluorescence in two rice varieties differing in sensitivity[J]. Chinese J Rice Sci,2005,19(4):381-383.(in Chinese)
[12]李平,刘鸿先,王以柔,等.低温对杂交水稻及其亲本三系始穗期旗叶光合作用的影响[J].植物学报,1990,32(6):456-464.
Li P,Liu H X,Wang Y R,et al.Effect of chilling on photosynthesis in flag leaves at primary heading stage of F1 hybrid rice cultivars and their parental lines(Three lines)[J]. Acta Botanica Sinica,1990,32(6):456-464.(in Chinese)
[13]李平,陈贻竹,李晓萍,等.籼稻的耐冷性与亲本的关系[J].植物学报,1994,37(7):544-551.
Li P,Chen Y Z,Li X P,et al.The cold tolerance of INDICA type rice in relation to its parents[J].Acta Botanica Sinica,1994,37(7):544-551.(in Chinese)
[14]Kaniuga Z,Zabek J,Sochanoeicz B.Photosynthetic apparatus in chilling-sensitive plants Ⅲ:contribution of loosely bound manganese to the mechanism of reversible in activation of Hill reaction activity following cold and dark storage and illumination of leaves[J].Planta,1979,144:490-502.
[15]张金恩,聂秋生,李迎春,等.颖花分化期低温处理对早稻叶片光合能力和产量的影响[J].中国农业气象,2014,35(4): 410-416.
Zhang J E,Nie Q S,Li Y C,et al.Effects of low temperature stress on the photosynthetic capacity and yield components of early rice at the spikelet differentiation stage[J].Chinese Journal of Agrometeorology,2014,35(4):410-416.(in Chinese)
[16]邓化冰,车芳璐,肖应辉,等.开花期低温胁迫对水稻花粉性状及剑叶理化特性的影响[J].应用生态学报,2011,22 (1):66-72.
Deng H B,Che F L,Xiao Y H,et al.Effects of low temperature stress during flowering period on pollen characters and flag leaf physiological and biochemical characteristics of rice[J].Chinese Journal of Applied Ecology,2011,22(1):66-72. (in Chinese)
[17]黄宇,王华.寒露风对水稻生产的影响[J].湖南农业,2008, (9):12.
Huang Y,Wang H.Effect of autumn low temperature on rice production[J].Hunan Agricuture,2008,(9):12.(in Chinese)
[18]邱上深.水稻冷害机制探讨[J].中国农业气象,1994,15(5): 41-45.
Qiu S S.Investigation of the mechanism of rice cold damage[J].Chinese Journal of Agrometeorology,1994,15(5): 41-45.(in Chinese)
[19]王丽娟,李天来,崔娜.夜间不同时段低温对番茄光合产物积累与分配的影响[J].沈阳农业大学学报,2006,37(6): 811-815.
Wang L J,Li T L,Cui N.Effects of different Time Lowtemperature at night on dry matter accumulation and distribution in tomato[J].Journal of Shenyang Agricultural University,2006,37(6):811-815.(in Chinese)
[20]王丽娟,李天来,齐红岩,等.长期夜间亚低温对番茄生长发育及光合产物分配的影响[J].沈阳农业大学学报,2006, 37(3):300-303.
Wang L J,Li T L,Qi H Y,et al.Effects of Long-term Sub-low Temperature on growth and development and dry matter distribution in Tomato[J].Journal of Shenyang Agricultural University,2006,37(3):300-303.(in Chinese)
[21]王保生,杜筱玲,刘文英,等.QX/T94-2008寒露风等级[S].北京:气象出版社, 2008.
Wang B S,Du X L,Liu W Y,et al.QX/T94-2008 grade of autumn low temperature[S].Beijing:China Meteorological Press,2008.(in Chinese)
[22]薛香,吴玉娥,陈荣江,等.小麦籽粒灌浆过程的不同数学模型模拟比较[J].麦类作物学报,2006,26(6):169-171.
Xue X,Wu Y E,Chen R J,et al.Comparison of different mathematical equations for simulating the grain filling process of wheat[J].Journal of Triticeae Crops,2006,26(6): 169-171.(in Chinese)
[23]朱庆森,曹显祖,骆亦其.水稻籽粒灌浆的生长分析[J].作物学报,1988,14(3):182-192.
Zhu Q S,Cao X Z,Luo Y Q.Growth analysis on the process of grain filling in rice[J].Acta Agronomica Sinica,1988, 14(3):182-192.(in Chinese)
[24]顾世梁,朱庆森,杨建昌,等.不同水稻材料籽粒灌浆特性的分析[J].作物学报,2001,27(1):7-14.
Gu S L,Zhu Q S,Yang J C,et al.Analysis on grain fil1ing characteristics for different rice types[J].Acta Agronomica Sinica,2001,27(1):7-14.(in Chinese)
[25]Sharp R E,Poroyko V,Hejlek L G.Root growth maintenance during water deficits:physiology to functional genomies[J].J Exp Bot,2004,55:2343-2351.
[26]杨猛,魏玲,胡萌,等.低温胁迫对玉米幼苗光合特性的影响[J].东北农业大学学报,2012,43(1):66-70.
Yang M,Wei L,Hu M,et al.Effect of low temperature stress on photosynthetic characteristics of maize seedling[J]. Journal of Northeast Agricultural University,2012,43(1): 66-70.(in Chinese)
[27]李平,王以柔,陈贻竹,等.低温对杂交水稻乳熟期剑叶光合作用和光合产物运输的影响[J].植物学报,1994,36(1): 45-52.
Li P,Wang Y R,Chen Y Z.Effects of chilling temperature on photosynthesis and photosynthate transport in flag leaves of hybrid rice at milky stage[J].Acta Bot anica Sinica,1994, 36(1):45-52.(in Chinese)
[28]王连敏,王立志,张国民.寒地水稻耐冷基础的研究Ⅲ:花期低温对水稻结实的影响[J].中国农业气象,1997,18(5):9-11.
Wang L M,Wang L Z,Zhang G M.Effect of cooling temperature at flowering stage on spikelet fertility of rice plant[J].Chinese Journal of Agrometeorology,1997,18(5): 9-11.(in Chinese)
[29]朱海霞,王秋京,闫平,等.孕穗抽穗期低温处理对黑龙江省主栽水稻品种结实率的影响[J].中国农业气象,2012,33(2): 304-309.
Zhu H X,Wang Q J,Yan P,et al.Effect of low temperature at heading stage on seed setting rate of major rice varieties in Heilongjiang province[J].Chinese Journal of Agrometeorology, 2012, 33(2):304-309.(in Chinese)
[30]Satake T,Shibata M.Male sterility caused by cooling treatment at the young micro spore stage in rice plants X X XI:four components participating in fertilization[J].Japanese Journal of Crop Science,1992,61:454-462.
[31]徐田军,王荣焕,赵久然,等.不同播期条件下‘京单68’和‘郑单958’的籽粒灌浆特性研究[J].农学学报,2013,3(10): 1-5.
Xu T J,Wang R H,Zhao J R,et al.The study on the grain filling characteristics of ‘Jingdan68’ and ‘Zhengdan958’under different sowing dates[J].Journal of Agriculture,2013, 3(10):1-5.(in Chinese)
Effect of Low Temperature at Flowering Stage on Late Rice Dry Matter Accumulation and Distribution
BAI Guang-zhi1, LIU Shou-dong1, YU Yan-wen1, YANG Zai-qiang1, YIN Jian-min2
(1.Collaborative Innovation Center on Forecast and Evaluation of Meteorological Disasters, Nanjing University of Information Science & Technology, Nanjing 210044,China; 2.Jiangxi Province Climate Center, Nanchang 330000)
Abstract:To study the impact of low temperature during flowering stage to photosynthesis, grain filling process, and dry matter accumulation and distribution, we took Guangliangyouxiang 66as a material in artificial control test in Nanjing University of Information Science & Technology from August to October in 2014. The low stress temperature was set as 22℃/17℃(day/night), and processing time was 2d, 4d, 6d and 8d (denoted as T2d, T4d, T6d and T8d), and 32℃/25℃ (day/night) as the control (CK). The results showed that: (1)after low temperature treatment, maximum photosynthetic rate (Pmax) of T2d changed smaller, and that of T4d and T6d were respectively 11.66% and 11.66% lower than CK. Pmaxof T8d was only 17.25μmol·m−2·s−1, which decreased by 28.54% than CK. In addition, the light saturation point (LSP) reduced after low temperature treatment, and the light compensation point (LCP) increased significantly. Compared with CK, T2d, T4d, T6d, T8d LSP was reduced by 6.68%, 8.08%, 8.63%, 10.87%, but LCP increased 2, 3, 4, 6 times. This means low temperature treatment can reduce thephotosynthetic capacity of late rice leaf. (2)When modeling grain fitting process of different treatments, we found that with more treatment time, the time required to reach the maximum filling rate(T-GRmax) was more, the largest grain weight(A) and time of the active period (T90) were less, which resulting a decline of average filling rate (GR). And this indicated that the low temperature will slow the process of rice grain filling. (3)The longer processing time, the lower the seed setting of rate rice. Treatment no more than 2d, the seed setting would not change significantly, but treatment more than 4d would result seed setting significant decline. Compared to CK, seed setting of T8d reduced by 34.39%. (4)The total dry matter was negatively correlated with low temperature treatment time, and the total dry matter would significant reduced if low temperature treatment after 6d. That of T8d showed a 28.05% decline compared to CK, which leaf and spike contributed obviously. And after 4d treatment, dry matter of leaf and spike would significant decline. Compared to CK, dry matter of leaf and spike of T8d were reduced by 42.57% and 38.07%. Result showed that low temperature at flowering stage could inhibit late rice photosynthesis. More than 4d treatment could significantly reduce the seed setting, slow down the process of grain filling, and those were adverse to the yield formation.
Key words:Low temperature at flowering stage; Late rice; Photosynthesis; Grain filling; Dry matter
doi:10.3969/j.issn.1000-6362.2016.02.0103
* 收稿日期:2015-07-21**通讯作者。E-mail:yzq@nuist.edu.cn
基金项目:国家科技部农业科技成果转化资金项目(2012GB24160606)
作者简介:白光志(1971-),硕士生, 主要从事农业气象灾害研究。E-mail: 00687@nuist.edu.cn
