云南拉沙山黑白仰鼻猴游走行为范围分析
2016-05-14王云李智宏
王云 李智宏


摘 要 经过2014年9月-2015年8月对拉沙山黑白仰鼻猴游走行为的研究显示:猴群夏季日移动距离最长
1 826.10±603.89 m,其次为秋季1 142.38±217.46 m、春季1 032.15±597.80 m和冬季839.36±643.81 m,并且夏季分别与春季P=0.007,秋季P=0.026和冬季P=0.000的差异性显著;不同季节的活动海拔秋季最高
3 478.13±53.17 m,夏季最低3 080.02±54.71 m,春季和冬季分别为3151.18±54.71 m和3 302.95±48.91 m;全年家域为9 km2,4季活动范围大小分别为:春季5.3 km2、夏季4.5 km2、秋季4.9 km2、冬季2.7 km2。春、夏、秋、冬猴群的移动时间比例(Chi-Square test x2=128.49 df=3 P=0.000)和取食(Chi-Square test x2=65.50 df=3 P=0.000)的时间比例都极显著。移动时间比与叶的可获得性显著正相关(r=0.691 n=12 P=0.013),不同月份猴群的月均日移动距离与叶的可获得性显著正相关(r=0.788 n=12 P=0.002)。春季和夏季猴群以“高成本-高收益”的策略在食物斑块间移动较远的距离来获取高质量食物;而冬季猴群则选择“低成本-低收益”的策略,增加取食时间,减少移动时间并减少移动距离。该区域常年人为干扰较强,主要的干扰方式烧炭占55%、砍豆杆占23%、盗伐占12%,其次是放牧和挖药材等占10%。
关键词 黑白仰鼻猴;日移动距离;活动范围;云南拉沙山
中图分类号:Q958 文献标志码:A 文章编号:1673-890X(2016)12-00-06
仰鼻猴属(Rhinopithecus)现存5个种:金仰鼻猴(Rhinopithecus roxellana)、黑白仰鼻猴(R. bieti)、灰仰鼻猴(R. brelichi)、黑仰鼻猴(R. strykeri)和越南仰鼻猴(R. avunculus)。除越南仰鼻猴外,其余在我国均有分布,且前三3种为我国特有物种。黑白仰鼻猴的分布南北跨越滇西北和藏东南6个县:西藏芒康,云南德钦、维西、丽江、兰坪和云龙[1],适宜生境约为
4 169 km2,现存数量有1 500~1 700只,约有13群(Xiao et al.,2003)。为了更好的保护这一濒危物种,前人主要从其分布(Long et al.,1994)、食性(Wu,1993;Yang & Zhao,2001)、生境(Xiao et al.,2003;Li et al.,2004)等方面做了研究。
灵长类游走行为变化主要受食物季节性变化的影响(Kaplin,2001)。由于温带森林中植物生产力和多样性波动强烈(Latham & Ricklefs,1993;Cramer et al.,1999),所以食物季节性变化在温带森林中比较明显。而温带灵长类会选择“高成本-高收益”和“低成本-低收益”策略来适应相应环境的不同季节温度、食物或营养的变化(Grueter & Li,2009)。“高成本-高收益”策略在温带灵长类中不常见,往往需要投入更多的能量用于移动来获取高质量食物并从中获取能量和营养(Sayers & Norconk,2008)。而“低成本-低收益”策略则常见于温带灵长类,表现为减少移动和其他活动以减少能量支出,并以取食低质量食物而获得能量(Ding & Zhao,2004)。云南省最南端富合山和整个黑白仰鼻猴分布最北部的西藏小昌都猴群当夏季喜食食物丰富的时候都以增加日移动距离和活动范围来获得高质量的食物,而冬季则采取缩短日移动距离,同时取食低质量食物的适应策略[2]。Liu(2004)认为,云南符合黑白仰鼻猴群春冬季选择“低成本-低收益”策略,在低海拔地区及过夜地附近活动,以取食树叶为主,并缩短日移动距离。Von Hippel(1998)认为,黑白仰鼻猴的游走趋向短时间移动和在离过夜地较近的区域活动来获得更多更高质量的食物;同时,可以降低能量消耗和捕食风险。尽管以上研究有一定的相似性,但并不统一。对于分布于云南省次南端的拉沙山黑白仰鼻猴群不同季节到底选取“高成本-高收益”策略还是“低成本-低收益”策略,甚至是其他策略有待进一步验证。
黑白仰鼻猴生存环境的恶劣程度从北到南逐渐减弱。栖息地海拔从北到南逐渐降低,生境多样性逐渐增加,这暗示生境质量从南到北逐渐下降,单位面积生境能够承载的量也逐渐下降[3]。相应的,食物种类、分布模式(均匀分布vs斑块分布)、食物斑块属性(丛集度、斑块间距离及其可通行状况)从北到南也有渐变过程;陈永春[4]也认为,由于黑白仰鼻猴南北分布区的生境差异可能导致南北猴群的生态适应机制的不同。进而了解拉沙山黑白仰鼻猴的生态适应机制及其适应可塑性,为物种保护管理措施的制定和实施提供科学依据。
1 研究方法
1.1 研究地点、对象和时间
研究地(见图1)位于云南省兰坪县云岭省级自然保护区拉沙山 (北纬26°20′, 东经99°15′)。拉沙山行政区划隶属兰坪县啦井镇期井村,最高峰海拔 3 854 m。目前,拉沙山猴群家域被低海拔(<2 800 m)村庄和耕地以及>3 600 m 的高山牧场包围,猴群被限制在海拔2 900~3 600 m的林带内活动[5]。猴群活动区植被随海拔从低到高呈现带状分布,依次为针叶林、针阔混交林、落叶阔叶林和暗针叶林,林下主要是杜鹃灌丛和箭竹林 (Fargesia spp.)(Huang,2009)。拉沙山地区月均温4.4(2月)~17.5 ℃ (7月)。5-10月为雨季,降水量占全年的79%;11月-翌年4月降雨稀少(Huang,2009)。
研究对象为分布于云南省黑白仰鼻猴分布区次南部的拉沙山猴群,最南端为龙马山猴群。该研究区域猴群由11个OMU和1个AMU 组成, 种群大小约130只[5]。根据该地区的气候划分春(3-5月)、夏(6-8月)、秋(9-11月)、冬(12-2月)4个季节,且从2014年9月至2015年8月,对该猴群进行了为期12个月的连续跟踪观察。
1.2 数据收集
1.2.1 日移动距离及活动海拔
本研究从2014年9月-2015年8月,历时12个月(除大雾、大雪、大雨天气外)。选取猴群活动区域对面视野较为开阔的至高点(如山脊、岩石、牧场等),借助单筒望远镜(Leica,T77)对猴群进行全天(从猴群07:00-09:00出过夜地开始,到18:00-20:00进入过夜地结束)连续跟踪观察,每隔半小时在1∶50 000的地形图上标注猴群中心位置(当猴群跟丢时,根据声音、树枝摇晃以及粪便等痕迹判断),把猴子最多的树或其他标志物记为猴群中心点。
1.2.2 活动时间分配
为满足取样的均衡性,每10 min采取(从左到右,然后从右到左的顺序)一次瞬时取样(Instantaneous Sampling)记录猴群活动状态(觅食、移动、休息、梳理、玩耍及其他行为)。分别用扫描到的觅食、移动、休息、梳理、玩耍及其他行为猴子的个体数除以扫描到的猴子总数来获得猴群的活动时间分配。
1.2.3 物候监测
借助瞬时扫描法记录不同季节、不同年龄-性别个体采食植物的种类和部位(叶、花、果实、树皮和种子等),根据觅食时间多少依序确定猴群主食种类。在确定不同季节主食资源分布模式(位置、范围和面积)的基础上,结合地形和实际的可行性设置5条样线,然后根据植被分布块特征及其主食分布状况(均匀vs聚集分布)确定样方数量(27个)和位置。沿样线每隔海拔50 m(根据海拔梯度和食物分布情况而定)设置一个样方(10 m×20 m),并在4个角和中间各设置1个小样方(1 m×1 m)。记录每个样方中的主食种类、数量及其他特征(如是乔木,记录取食部位、胸径、高度、冠幅、树冠高度和盖度;灌木则记录其取食部位、均高、盖度;草本则记录其种类、部位和盖度;竹笋记录颗数、高度)。
1.2.4 人为干扰
首先,根据人类活动干扰的方式及其对植被的破坏强度主要划分为烧炭、盗伐、砍豆杆和其他(包括放牧、找菌子、挖药材及采蜂蜜)4类。由于研究站点位于村民进山的主要通道口,所以能够较为准确、真实地直接记录人类活动干扰的情况。我们主要通过直接记录得到数据。但由于跟踪猴群的时间限制,无法保证数据的长期持续性。小部分数据只能通过向导帮助记录或者询问当地村民(主要是护林员)得到。首先,需要记录进山人数、马匹和牛羊等牲畜数量,人们的活动方式(砍豆杆、砍伐树木或者烧炭、放牧等)。其次,记录活动方向及位置以及所使用的工具。为了确保数据的真实性,有时通过向导或其他村民得到的数据还需结合研究者本人所听到的人们活动的声音和实地活动留下的痕迹(烧火)进行二次判断。
1.3 数据分析
首先,借助ArcGIS校正地形图并数字化,以获取地形图中数据点信息(如经纬度、海拔、坡向和坡度等);然后利用最小凸边形法(Minimum Convex Polygon: MCP)(Kaplin,2001)计算年家域面积和不同季节猴群的活动范围大小,利用ArcGIS要素转线法计算日移动距离。采用SPSS16.0通过单因子方差分析(one-way ANOVA)判断不同季节日移动距离的差异性。通过卡方检验分析不同季节猴群活动时间分配的差异性;同时,借助SPSS16.0相关性分析法分别对不同月份的月均日移动距离和活动时间分配与食物的可获得性进行相关性分析。其次,人为干扰强度分别以直接记录的人数除以总人数获得。
2 结果
2.1 日移动距离
在借助ArcGIS计算出不同季节的日均移动距离的基础上,通过one-way ANOVA 分析发现,不同季节的日均移动距离差异显著(F=6.905 df=3 P=0.001),分别为:春季1 032.15±597.80 m、夏季1 826.10±603.89 m、秋季1 142.38±217.46 m、冬季839.36±643.81 m。通过Post-hoc两两配对多重比较分析后,春季和夏季(P=0.007)、夏季和秋季(P=0.026)、夏季和冬季(P=0.000)的日移动距离较为显著(见图1)。全年日移动距离冬季最短(168 m),而夏季最长(3 084 m)。
2.2 活动时间分配
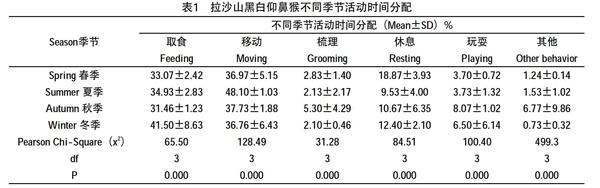
研究期间,将猴子行为主要划分为取食、移动、梳理、休息、玩耍和其他6类。猴群春、夏、秋、冬四季的取食变化极显著(Chi-Square test x2=65.50 df=3 P=0.000),冬季取食时间明显高于其他季节41.33±8.51%;不同季节移动极显著(Chi-Square test x2=128.49 df=3 P=0.000),夏季食物丰富移动时间最长48.00±10.54%,在食物相对匮乏的冬季,为节省能量开支移动时间最短37.00±6.56%;不同季节的梳理行为存在显著差异(Chi-Square test x2=31.28 df=3 P=0.000),梳理最长的时间是在高质量食物(果实)丰富和可获得性较高的秋季5.33±4.51%。同样不同季节的休息存在极显著差异(Chi-Square test x2=84.51 df=3 P=0.000),其中春季的休息时间最长18.67±3.79%,而夏季的休息时间最短9.67±4.04%;同理,该猴群不同季节的玩耍(Chi-Square test x2=100.401 df=3 P=0.000)以及其他(Chi-Square test x2=499.3 df=3 P=0.000)行为的活动时间分配都表现出显著差异。猴群不同季节的活动时间分配详见表1。
2.3 物候监测
拉沙山黑白仰鼻猴活动区域的食物物候见图2。芽期为3-5月,嫩叶期集中于4-6月,而果食则在8月17日-9月17日达到峰值,直至11月下旬落叶落果结束。通过对活动时间分配与主食物候的相关性分析,我们得到取食时间比例与芽、叶、花、果的可获得性没有达到统计学上的标准(芽:r=-0.097 n=12 P=0.763;叶:r=-0.380 n=12 P=0.223;花:r=-0.022 n=12 P=0.946;果:r=-0.521 n=12 P=0.082)。同样,移动时间比例与花的可获得性之间的相关性不明显,而与叶的可获得性之间呈正相关,并且显著正相关(叶:r=0.691 n=12 P=0.013;花:r=0.499 n=12 P=0.099),但是,移动时间比例同样与芽、果的可获得性相关性也不明显(芽:r=-0.253 n=12 P=0.428;果:r=-0.015 n=12 P=0.962)。取食与芽、果的可获得性的相关性不明显可能与芽和果的短时期性有关。全年不同月份猴群的月均日移动距离与芽和果的可获得性相关性不明显(芽:r=-0.207 n=12 P=0.518;果:r=0.242 n=12 P=0.449);与叶的可获得性呈极显著正相关(r=0.788 n=12 P=0.002);与花的可获得性呈显著正相关(r=0.668 n=12 P=0.018)。
2.4 活动海拔与活动范围大小
拉沙山黑白仰鼻猴的季节性活动海拔见表2,该猴群不同季节的活动海拔呈现极显著的差异性(one-way ANOVA F=99.13 P=0.000)。秋季活动海拔最高3 478.13±53.17 m;夏季最低3 080.02±54.71 m,这可能与食物分布有关;冬季仅次于秋季3 302.95±48.91 m,这可能是受到光照的影响,猴群为了更早、更长时间的接受到光照,以此弥补冬季低温和食物匮乏条件下调节体温的压力。而春季(3 151.18±54.71 m)则介于夏冬之间;但是可能由于食物等因素的季节性变化影响,全年活动海拔最低(2 909.65 m)在春季,最高在秋季(3 629.53 m)。
拉沙山黑白仰鼻猴全年活动范围见图3。
由于受到人为干扰的影响,拉沙山黑白仰鼻猴2014年9月-2015年8月,年家域仅为9 km2,但实际家域面积已经大于9km2,该猴群有向北扩展的趋势。利用ArcGIS里的最小凸多边形法计算得到不同季节该猴群活动范围大小分别为:春季最大5.3 km2,夏季4.5 km2,秋季4.9 km2,而冬季(2.7 km2)最小。猴群春季活动范围最大主要受到食物物候的影响,为了获得高质量的嫩芽及花猴群会在特定的时期从一个食物斑块移动到另一个特定的食物斑块。而冬季则由于低温和食物短缺的压力,猴群为了节省能量支出选择在小范围内活动并取食低质量的食物。
2.5 人为干扰
根据直接记录,拉沙山人为活动干扰情况见表3,全年干扰强度及主要方式见图4。除了6月、7月和8月降雨较多没有直接记录到人为活动外其余月份均有记录。但该时段经常听到山里有人吼叫,且直观看到有烧火的烟雾和林内被挖过的土坑,经过与向导和其他村民的核实,可判断为过来挖药材的傈僳族人。重楼、竹泥和菖蒲等药材的一直以来都是他们的重要经济来源,他们的活动海拔集中在2 800~3 600 m的整个猴群活动区。该地区人们的活动方式主要包括烧炭、盗伐、砍豆杆、放牧、挖药材和采菌子,偶有采蜂蜜者。烧炭活动主要是2014年9-12月,期间基本固定有6人和7匹马专门在分布有较多栎树的区域进行烧炭活动,同时他们还进入森林到达少数分布有落叶松(Larix gmelinii)、五角枫(Acer mono)及红豆杉(Taxus chinensis)的区域盗伐,活动海拔为2 800~3 600 m。另外,该地区1-5月的人为干扰最强且对植被破坏力最强的方式是砍豆杆(用于当地主要经济作物白云豆的爬杆)。主要的砍伐对象是分布于2 800~3 300 m的空心箭竹(Fargesia edulis)。而该地区放牧活动常年都有,少数几户人家大数目牛羊群(>200头)主要在高处牧场(3 300~3 800 m),大部分人家少数牛羊群(2~50头(只))则活动较散,遍及2 800~3 800 m的整个猴群活动区域。
3 讨论
不同取食策略影响灵长类游走行为的季节性差异。通常,食物的分布决定了猴群低能耗的取食策略,猴群往往在某种或者多种丰富食物的区域内取食而减少移动(Clutton-Brock & Harvey,1977)。而非洲加蓬马坎黑疣猴在果实丰富的季节日移动距离较长,在果实匮乏的季节日移动距离短(Fleury & Gauiter,1999)。同样,在西藏小昌都猴群的日移动与食物可获得性正相关相关,即猴群在夏季高温、食物可获得性高的时候增加日移动距离(向左甫,2005)。与敏长臂猿和分布在Chunati的西白眉长臂猿就没有发现它们的移动距离与食物密度和可利用性间存在相关性(Ashan,1994)相似,虽然拉沙山黑白仰鼻猴的日移动距离与高质量的芽和果实的相关性不明显,但是夏季该猴群的日移动距离与春季、秋季和冬季的差异性显著,夏季日移动距离最长。在果食匮乏的季节,东白眉长臂猿倾向于选择低质量的食物,此时日移动距离降到最低[6],对于拉沙山黑白仰鼻猴,在冬季高质量食物匮乏时,日移动距离也最低。
春夏季喜食食物丰富的时候黑白仰鼻猴日移动距离较长且活动范围较大(Kirkpatrick,1998;Liu,2004)。同样,拉沙山黑白仰鼻猴春夏季食物丰富的时候日移动距离较长且活动范围较大,其中春季的活动范围全年最大,夏季的日均移动距离全年最长。当某种果实集中于短时间内成熟时,与云南萨马阁黑白仰鼻猴总是在一个较小的范围中往返移动,反复使用果实成熟时间不同的生境(食物)斑块,结果增加了移动距离但并没有扩大活动范围(Grueter,2008)的异同在于,拉沙山黑白仰鼻猴在夏秋季果实成熟的时期,日移动距离(夏季:1 826.10±603.89 m,秋季:1 142.38±217.46 m)与活动范围(夏季4.5 km2,秋季4.9 km2)都较大。广西弄岗黑叶猴在喜食食物缺乏的季节,会扩大家域面积、增加漫游距离来获取充足的食物[7],而拉沙山黑白仰鼻猴在冬季高质量食物匮乏时日移动距离短,活动范围最小,用于移动的时间最短(36.76%±6.43%)且取食的时间最长(41.50%±8.63%),所以冬季该猴群主要选择“低成本-低收益”的适应策略。
拉沙山黑白仰鼻猴不同季节的移动时间比例从多到少分别为夏季、秋季、春季和冬季,而且该猴群的日移动距离夏季明显比春季、秋季和冬季要长,所以夏季该猴群主要选择“高成本-高收益”的适应策略通过增加日移动时间并增加日移动距离,投入更多的能量来获得高质量食物(嫩叶、果实和竹笋)。其次,秋季该猴群用于移动的时间比例仅次于夏季这可能是秋季猴群的日均移动距离小于夏季的原因之一。同样,冬季猴群用于移动的时间最少,而用于取食的时间最长,所以冬季该猴群的日均移动距离明显低于夏季,而导致夏冬两季比较明显的反差则可能是由于夏季食物丰富,冬季食物匮乏,夏季高温且光照时间长而冬季低温光照时间短等原因。虽然全年日移动距离最短的是冬季(168 m),最长的是夏季(3 084 m),但是,可能是居于拉沙山猴群食物分布的斑块化,春季(574~2 288 m)的日移动距离波动比较大且活动范围最大,猴群会在冬末春初移动较长的距离到达特定食物斑块取食特定的物种[山鸡椒(Litsea cubeba)],并且5月17日,猴群短时间移动较长的距离(2 288 m)从整个猴群活动区域的北边到达东边(经过当天的人为活动迹象排除了人为干扰的因素,并且实地调查发现猴群所到区域竹笋分布较多,发现有明显取食痕迹的竹笋)。同时,春季猴群用于休息的时间最长(18.87%±3.93%),所以,春季猴群会在短时期内以短时间长距离的移动,采取“高成本-高收益”的适应策略来获取高质量食物。同理,全年猴群冬季的移动距离最短,分配于移动的时间比例最少且活动范围最小,可能与冬季高质量食物的缺乏而以低质量的松萝为主,没有进行斑块间的长距离移动的原因。早期的研究表明疣猴主要以树叶为主食(Culton-Brock,1975),Oates(1977)研究发现,树叶在东非疣猴的食物中占81%,Huang(2003)的研究认为果实在黑白仰鼻猴的食物谱中也仅占7%~20%。因此,这些研究与本研究中拉沙山黑白仰鼻猴的日移动距离与果实和芽的可获得性相关性不明显,却与叶的可获得性显著正相关趋同。笔者认为,这应该与芽和果实的短时期性有关,而对于拉沙山黑白仰鼻猴秋季日移动距离的波动性相对较小(707~1 561 m)的原因一方面可能由于果实的短时期性,另一方面则由该地区食物的斑块化分布决定。猴群为了在短时期内对高质量果实的最高效利用而进行食物斑块间的移动。
Rui-Chang Quan和Guopeng Ren(2011)通过研究发现,黑白仰鼻猴冬季喜欢在高海拔的区域活动是受光照的影响,猴群在冬季低温时,由于低食物质量和体温调节的压力而选择光照时间最早、持续时间最长的高海拔区域活动。 同样,本研究发现拉沙山猴群的活动海拔冬季仅低于秋季,且猴群喜欢在南坡活动。此外,全年猴群的活动海拔春季并非最低而夏季最低,笔者认为,这与该区域常年人为活动的干扰存在密切的相关性。夏季人们集中于2 800~3 600 m的海拔带挖药材,并且高出牧场的牧草生长良好,牲畜往高海拔迁移。而春季当地村民则集中于2 800~3 300 m的海拔带中的几个空心箭竹分布区活动,所以春季人们将猴群往高海拔挤,而夏季则将猴群往低海拔压。常年对猴群的生存生境的破坏较强并加速了该区域生境的破碎片段化。
参考文献
[1]龙勇诚,钟泰,肖李.1996年滇金丝猴地理分布 种群数量与相关生态学的研究[J].动物学研究,1996,17(4):437-441.
[2]向左甫.西藏红拉雪山自然保护区黑白仰鼻猴(Rhinopithecus bieti)的生态与行为研究[D].昆明:中国科学院昆明动物研究所,2005.
[3]肖文,霍晟,向左甫,等.黑白仰鼻猴种群生存力初步分析[J].动物学研究,2005(1):9-16
[4]陈永春,肖林.滇金丝猴的生境适应对策[J].林业调查规划,2012(2):33-36.
[5]王双金.云南拉沙山黑白仰鼻猴(Rhinopithecus bieti)繁殖行为和食性[D].昆明:西南林学院,2012.
[6]吴建普,周伟,周杰珑,等.高黎贡山赧亢白眉长臂猿食性及日取食量[J].动物学研究,2009,30(5):539-544.
[7]黄中豪,周岐海,黄乘明,等.广西弄岗黑叶猴的家域和日漫游距离[J].兽类学报,2011(1):46-54.
(责任编辑:赵中正)
