利用单片段代换系测交群体定位玉米产量相关性状的杂种优势位点
2016-04-27薛亚东张向歌李慧敏孙高阳李卫华谢慧玲汤继华
彭 倩 薛亚东 张向歌 李慧敏 孙高阳 李卫华 谢慧玲 汤继华
省部共建小麦玉米作物学国家重点实验室/河南省粮食作物协同创新中心/河南农业大学农学院,河南郑州450002
利用单片段代换系测交群体定位玉米产量相关性状的杂种优势位点
彭倩**薛亚东**张向歌李慧敏孙高阳李卫华谢慧玲汤继华*
省部共建小麦玉米作物学国家重点实验室/河南省粮食作物协同创新中心/河南农业大学农学院,河南郑州450002
摘要:杂种优势利用是提高农作物产量与品质的一种重要途径,而明确杂种优势的遗传机制将促进优良玉米新品种的选育,但是截至目前其遗传机制仍不清楚。本研究以玉米自交系lx9801背景的昌7-2单片段代换系为基础材料,利用与自交系T7296的测交群体,对昌7-2和lx9801相应染色体片段与T7296之间存在差异的杂种优势位点进行了分析,共检测出64个不同穗部性状和产量的杂种优势位点(HL),其中23个在2个环境中同时被检测到,包括4个穗长的HL,4个穗粗的HL,4个穗行数的HL,7个行粒数的HL和4个产量的HL,并在多个染色体片段上鉴定出同时包含产量及其构成因子的杂种优势位点,该研究为进一步解析玉米产量杂种优势形成的遗传机制奠定了材料基础。
关键词:玉米;染色体片段代换系;产量;杂种优势;数量性状位点
本研究由国家自然科学基金项目(31271732)资助。
This study was supported by the National Natural Science Foundation of China.
第一作者联系方式:E-mail:18613706657@163.com,Tel:0371-63558377**同等贡献(Contributed equally to this work).
杂种优势是生物界一种广泛存在的遗传现象,并在农作物和畜牧业育种工作中得到广泛利用。从Shull[1]在上个世纪初提出杂种优势概念的一个多世纪以来,科研工作者对杂种优势的遗传机制进行了大量研究,提出了显性、超显性和上位性等著名假说来解释杂种优势形成的遗传机制[2-5]。随着分子生物学研究的不断深入,科研工作者从基因组学[6]、转录组学[7]、蛋白质组学[8]、miRNA调控[9]以及关键基因的遗传转化[10]等方面揭示了杂种优势形成的可能遗传机制。
由于杂合是杂种优势产生的遗传基础,前人曾利用F2:3群体[11]、RIL测交或回交群体[12]、三交群体[13-14]、“永久F2”群体[15]等不同的遗传群体,通过对不同物种多个性状的QTL效应值分析或杂种优势位点定位剖析了杂种优势的遗传机制。Xiao等[16]利用水稻籼粳交的F7重组自交系与双亲回交群体,通过分析QTL的效应值,认为显性效应是杂种优势产生的主要遗传机制。Lu等[17]通过对玉米随机交配群体产量性状的QTL分析,认为超显性在玉米产量杂种优势中具有重要作用。Hua等[15]利用水稻RIL群体随机组配的“永久F2”群体,发现单位点水平上的超显性效应以及两位点水平上的显×显互作是水稻优良杂交种籼优63产量杂种优势形成的重要遗传机制。由于上述分离群体的遗传组成较为复杂,为简化分离群体的遗传背景,近期不同学者利用单片段代换系的测交或者回交群体对番茄[18-19]、水稻[20]、拟南芥[21]、棉花[22]等作物的杂种优势遗传机制进行了研究。
玉米是世界上第一个成功利用杂种优势的作物,也是世界上利用杂种优势面积最大的作物,此外还是经典遗传学研究的模式生物。在长期的玉米育种实践中,育种家根据不同种质材料来源及其配合力的高低将玉米种质资源划分为不同的杂种优势类群,并总结我国常用的杂种优势模式[23],这些研究减少了玉米育种过程中的盲目性,有效地提高了育种效率[24]。唐四平头和Reid是我国黄淮海夏玉米区和东北春玉米区常用的种质类群和一对杂优模式,本研究以来源于我国地方种质唐四平头2个骨干系昌7-2 与lx9801的一套单片段代换系为基础材料,利用来自Reid类群的自交系T7296作为测验亲本,组配了一套测交群体对玉米产量与4个穗部性状的杂种优势位点进行了分析,以期鉴定出昌7-2和lx9801对应染色体片段与T7296之间存在差异的杂种优势位点,为进一步揭示玉米杂种优势形成的分子机制提供材料平台。
1 材料与方法
1.1试验材料
基础材料是我国地方优异种质唐四平头类群的2个优良自交系昌7-2 和lx9801,以昌7-2为供体亲本、lx9801为受体亲本从800对SSR引物中选择了225对在2个亲本间存在多态性的引物。从BC3F1世代开始用分子标记选择只有一段供体染色体的株系,经过4个世代回交和3个世代自交,构建了184个lx9801背景的昌7-2单片段代换系,代换片段总长1683.33 cM,平均长度9.25 cM,覆盖玉米基因组的35.5% (图1)[25]。由于构建的单片段代换系在225对SSR标记检测下与lx9801相比只有1段昌7-2供体片段,因此背景回复率根据供体片段的长短不同基本在95%~98%以上。2012年冬在海南将单片段代换系群体与自交系T7296测交,组配了184个CSSLs× T7296的测交群体。自交系T7296选自Reid类群,而T7296×lx9801杂交组合是河南省审定的优良玉米杂交种豫单811。
1.2试验方法
2013年夏,将CSSLs×T7296测交群体、对照种豫单811 (T7296×lx9801)种植于河南长葛市试验田和鹤壁市农科院试验田(河南浚县),测交群体按完全随机区组设计,3个重复,单行区,行长4 m,每行15株,密度67 500株 hm-2,为提高杂种优势位点检测的准确性,每10个测交组合中添加1个对照。CSSLs群体(包含自交系lx9801、昌7-2和T7296)按照完全随机区组设计,3个重复,与测交群体分开种植在同一试验田中,每10个材料中同样添加1行lx9801作为对照。成熟后选择连续10株收获,自然晾干后分别考种,单穗调查穗长(cm)、穗粗(cm)、穗行数、行粒数和籽粒重,按照种植密度以籽粒重量折合产量(t hm-2)。
1.3数据处理与分析
由于本研究所利用的单片段代换系与受体亲本lx9801只存在一段供体染色体的差别,通过比较单个单片段代换系测交种与对照(lx9801×T7296)之间的差异,就可以鉴定出昌7-2供体片段与lx9801相应染色体片段和测验种T7296之间的杂种优势表现是否存在差异,即在2个自交系对应染色体片段上与T7296是否存在差异的杂种优势位点。
采用SPSS18.0统计软件,对两点试验材料的产量和穗部性状进行统计和相关性分析。以各试验点lx9801×T7296的观测值为对照,利用方差分析和t测验比较每个SSSL×T7296测验种单个性状与对照种之间的差异,在P≤0.05的显著水平下认为可能存在相应性状的杂种优势位点。在显著性检验的基础上,进一步利用多重比较对鉴定出的杂种优势位点进行分析,以剔除假阳性的杂种优势位点。杂种优势效应值用超标优势表示,超标优势(%) ={(SSSL× T7296)表型值-对照表型值}/对照表型值 × 100%。杂种优势位点以h+性状英文缩写+染色体序号+位点序号(如a,b,c,…)命名,如果一条染色体上仅有一个杂种优势位点则表示为h+性状英文缩写+染色体序号。
2 结果与分析
2.1测交群体产量与穗部性状的表型与杂种优势分析
CSSLs×T2796测交群体的产量和4个穗部性状在2个环境中均表现出较大的表型变异(表1)。穗长在长葛点和浚县点的平均值为16.78 cm和18.96 cm,变异范围为14.47~8.48 cm和16.95~20.36 cm,平均中亲优势值为 60.60%和59.13%,而对照种T7296×lx9801在2个环境中的平均穗长分别为16.78 cm和19.03 cm,中亲优势值为59.79%和58.75%。测交群体的行粒数在长葛点和浚县点的平均值分别为32.34和33.67,平均中亲优势值为55.16%和53.45%;而对照种T7296×lx9801的行粒数在长葛点和浚县点的平均值分别为32.13和33.78,中亲优势值为54.24%和57.62%。测交群体穗行数的平均值在长葛点和浚县点分别为14.72和14.05,中亲优势值为19.68% 和13.66%;对照种T7296×lx9801的穗行数在2个环境中的中亲优势值分别为19.43%和13.58%。测交群体的平均产量在长葛点和浚县点分别为8.44 t hm-2和10.05 t hm-2,平均中亲优势值分别为85.87%和81.78%,而对照种T7296×lx9801的产量在2个环境中的平均中亲优势值分别为74.89% 和79.56%。从群体的整体水平看,测交群体的4个穗部性状和产量的平均值与平均中亲优势值均与对照种相似,同时在玉米的4个穗部性状中穗长的平均中亲优势最强,其次分别是行粒数和穗行数,穗粗的中亲优势值最小,说明穗长和行粒数的杂种优势对产量的杂种优势贡献较大。

表1 CSSLs×T7296测交群体穗部性状与产量的表型与中亲优势表现Table 1 Performance and mid-parent heterosis of grain yield and ear traits in the CSSLs×T7296 test population
2.2测交群体产量与穗部性状的相关分析
CSSLs×T7296测交群体在2个环境中穗长与行粒数均呈显著正相关(表2),同时穗粗与穗行数在2个环境中也呈显著正相关,而穗部4个性状在2个环境中均与产量呈显著正相关。此外,穗长与穗粗在2个环境中均呈显著正相关,但是穗长与穗行数在浚县点呈显著负相关,在长葛点的相关不显著。
2.3玉米产量与穗部性状的杂种优势位点分析
通过CSSLs×T7296群体中每一个测交组合单个性状与对照之间的显著性分析和多重比较,在0.05%显著水平上共检测出64个产量与4个穗部性状的杂种优势位点(HL),其中23个在2个环境中同时被检测到(表3和表4)。在长葛点和浚县点分别检测到9个和8个穗长的HL,其中4个HL在2个环境中同时被检测到(表3和图1)。在第1染色体上1.08 bin上的hEL1b在长葛点和浚县点的超标优势分别为-9.25%和-8.29%。位于第3染色体上3.08 bin上的hEL3c在长葛点和浚县点的超标优势分别为10.15%和6.98%;而位于第7染色体上的hEL7a在长葛点和浚县点的超标优势分别为-7.66%和-8.23%。第4个在2个环境中共同被检测到的穗长杂种优势位点是hEL9,其在长葛点和浚县点的超标优势分别为5.26%和5.51%。
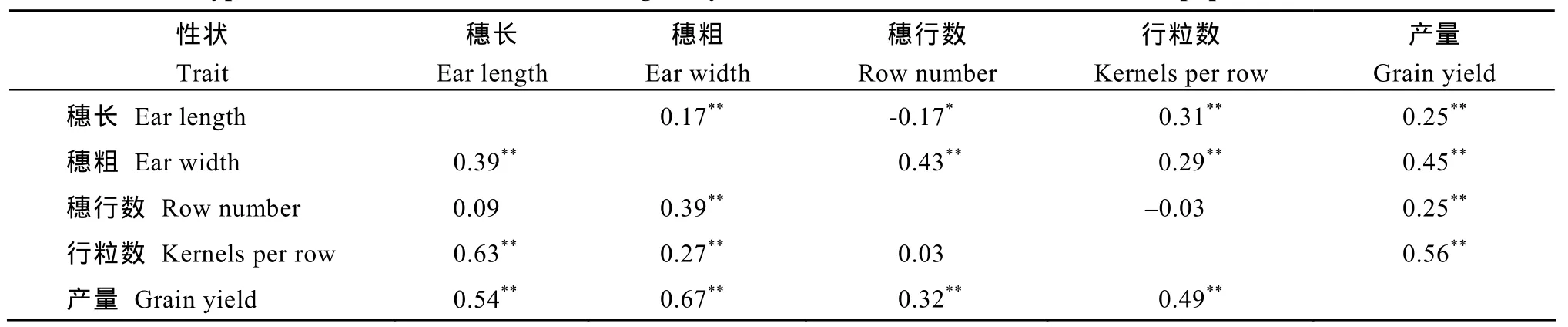
表2 CSSLs×T7296测交群体产量及穗部性状间的表型相关系数Table 2 Phenotypic correlation coefficients between grain yield and ear traits of the CSSLs×T7296 population in two environments
在长葛点和浚县点分别检测到8个和9个穗粗HL,其中4个在2个环境中同时被检测到(表3和图1)。位于第1染色体上的hED1a,在长葛点和浚县点超标优势分别为-6.71%和-4.45%;位于第5染色体上标记phi048-bnlg1306染色体区段上的qED5b,在长葛点和浚县点的超标优势分别为9.94%和6.43%。第6染色体上hED6c与对照相比在长葛点和浚县点可以使穗粗分别减少6.71%和6.57%。在第9染色体上也检测到1个共同控制穗粗的杂种优势位点hED9a,在长葛点和浚县点与对照相比穗粗分别增加8.42%和5.41%。
在长葛点和浚县点分别检测到6个和9个穗行数HL,其中4个在2个环境中同时被检测到。位于第1染色体1.08 bin上的点hRN1与对照相比在长葛点和浚县点可以使穗行数分别增加6.79% 和7.44%。位于第4染色体上的hRN4,与对照相比在长葛点和浚县点可以使穗行数减少9.50%和9.18%;另外2个在2个环境中同时检测到的穗行数杂种优势位点是hRN5b和hRN8,超标优势分别为-6.11%和-4.43%,-10.41%和-8.86%。
在长葛与浚县点各检测到13个行粒数HL,其中7个在2个环境中同时被检测到。在第1染色体上检测到2个共同的HL,hKPR1a和hKPR1d,在长葛点和浚县点与对照相比可以使穗行数分别增加12.89%和11.014%,15.72%和16.64%。在第2染色体上检测到1个共同的HL hKPR2b,在长葛点和浚县点的超标优势分别为17.24%和10.60%。在第3和第4染色体上同时检测到2个共同的HL hKPR3a和hKPR4b,在长葛点和浚县点的超标优势分别为14.89%和10.82%,-13.67%和-9.93%。另外2个同时被检测到的行粒数杂种优势位点分别是hKPR7a和hKPR10,其在长葛点和浚县点的超标优势分别为-14.45%和-13.58%,-10.94%和-9.93%。
在2个环境中共检测到13个产量杂种优势位点,其中4个HL在2个环境中同时被检测到(表4和图1)。位于第1染色体上的hGY1b在长葛点和浚县点的超标优势分别为9.41%和15.60%;位于第3染色体上的hGY3a在长葛点和浚县点的超标优势分别为14.62% 和9.93%。第6染色体上的hGY6与对照相比在长葛点和浚县点可以使产量分别减少16.35%和8.98%;而第7染色体上的hGY7a比对照相比在2个环境中可以使产量分别减少10.27%和21.99%。
3 讨论
前人研究结果与生产实践均证明增加密度是世界范围内提高玉米产量的一种重要因素。尽管随着播种密度的增加,杂种优势在玉米产量中的相对贡献率在逐渐下降,但是杂种优势在玉米产量贡献中的绝对量并没有发生明显的改变[25],杂种优势依然是保证优良杂交种产量的一个重要遗传因素,因此定位玉米杂种优势基因,剖析杂种优势的遗传机制仍将对玉米新品种的选育具有重要的促进作用。在玉米杂种优势遗传机制研究方面,Stuber等[22]利用(Mo17×B73)F3家系与双亲回交的2个分离群体,发现绝大多数QTL杂合子的表型值均高于任何纯合子的表型值,认为超显性是杂种优势产生的主要遗传基础。Tang等[27]利用豫玉22的一套“永久F2”群体对玉米株高的杂种优势位点进行了定位。近期Wei 等[28]利用一套许178背景上的综3单片段代换系与轮回亲本的回交群体,定位了玉米株型相关性状的杂种优势位点,发现超显性效应可能是杂种优势形成的重要遗传机制。Guo等[10]将来源于先锋种质杂种优势群的2个ARGOS1 (ZAR1)等位基因分别进行了遗传转化,发现转基因植株表现出不同杂种优势效应,说明不同杂种优势位点的等位基因之间的效应存在一定差异,从而为优异杂种优势等位基因的筛选与利用提供了理论依据。本研究利用我国生产上广泛利用的地方优异种质唐四平头的骨干自交系昌7-2与lx9801构建的单片段代换系群体,通过与Reid种质的代表性自交系T7296组配的测交群体,在单片段水平上分析了自交系昌7-2与lx9801相应染色体片段和T7296杂种优势的表现,在2个环境中同时鉴定出23个产量与4个穗部性状的对应染色体片段,该研究为等位基因之间存在不同的杂种优势效应提供了理论依据。

表3 在CSSLs×T7296群体中鉴定出的玉米穗部性状杂种优势位点Table 3 Heterotic loci for ear traits detected in the CSSLs×T7296 population in maize

(续表3 )

表4 在CSSLs × T7296测交群体中鉴定出的玉米产量杂种优势位点Table 4 Heterotic loci for grain yield detected in the CSSLs × T7296 population in maize

图1 玉米产量及穗部性状的杂种优势位点在染色体上的位置Fig.1 Chromosomal location of heterotic loci (HL) for grain yield and its components长葛:▽穗长HL,○穗粗HL,◇穗行数HL,□行粒数HL,☆产量HL;浚县:▼穗长HL,●穗粗HL,◆穗行数HL,■行粒数HL,★产量HL。Changge location:▽Ear length HL,○ Ear width HL,◇ Row number HL,□ Kernels per row HL,☆ Grain yield HL;Xunxian location:▼ Ear length HL,● Ear width HL,◆ Row number HL,■ Kernels per row HL,★ Grain yield HL.
比较本研究中鉴定的64个产量与穗部性状的杂种优势位点与Tang 等[29]的研究结果,发现在2个分离群体中只有1个穗长杂种优势位点位于相同的染色体片段上(el7,标记区间bnlg1805-umc1888;hEL7b,染色体片段bnlg2271-umc1112-bnlg1805);同时本研究在该染色体片段上还各检测到1个行粒数和产量的HL (hKPR7b,hGY7b),说明单片段代换系的测交群体对杂种优势位点具有更高的检测效率。此外,在本研究所检测的玉米产量与穗部性状的杂种优势位点中,一些染色体片段上同时检测到多个性状的杂种优势位点(图1),如在第3染色体上的phi374118-umc2258-bnlg1447的片段上同时检测到穗粗、穗行数和产量的HL (hED3a、hRN3和hGY3a),在第6染色体上的bnlg1732-umc1424-umc1296片段上同时检测到了穗粗、行粒数和产量的HL (hED6c、hKPR6c和hGY6),在第7染色体上的bnlg1792-umc1929-umc1585和bnlg2271-umc1112-bnlg1805片段上同时检测到控制穗长和行粒数的HL (hEL7a和hKPR7a,hEL7b和hKPR7b)等等,这些在一个染色体片段上同时检测到的共同HL性状往往是高度相关的性状,如穗长、行粒数与产量,穗粗与穗行数等,说明在玉米产量与穗部性状高度相关的性状之间可能存在相同的杂种优势遗传机制。
由于玉米优良组合选配的效率较低,而且具有极大的盲目性,为提高育种效率,玉米育种家根据长期的育种经验与配合力高低总结出不同种质类群之间的杂种优势利用模式,杂种优势模式间的选系组配出优良杂交组合的概率相对较高,已经成为玉米育种家普遍采用的一种方法[30]。尽管分子标记的出现特别是高密度SNP标记的应用为准确划分不同的杂种优势类群提供了有效的工具[24,31],但是并不是杂种优势利用模式内的任何自交系之间都能组配出优良的杂交组合,其制约因素就在于人们对杂优模式内的杂种优势位点及其效应仍然不清楚,导致在杂种优势模式内的育种工作中仍然存在较大的盲目性。本研究利用唐四平头的代表性自交系lx9801和昌7-2以及来源于Reid的自交系T7296对产量与穗部性状的杂种优势位点进行了鉴定,该研究可以为唐四平头与Reid杂种优势模式之间优良杂交组合的组配提供一定的理论依据。
4 结论
共检测出64个玉米产量与穗部性状的不同杂种优势位点,其中23个在2个环境中同时被检测到,包括穗长的4个,穗粗的4个,穗行数的4个,行粒数的7个以及产量的4个。在一些染色体片段上同时检测到控制玉米产量与穗部性状的杂种优势位点,说明产量与穗部高度相关的性状之间可能存在相同的杂种优势遗传机制。
References
[1]Shull G H.The composition of a field of maize.J Heredity,1908,4:296-301
[2]Bruce A B.The Mendelian theory of heredity and the augmentation of vigor.Science,1910,32:627-628
[3]Jones D F.Dominance of linked factors as a means of accounting for heterosis.Proc Natl Acad Sci USA,1917,3:310-312
然而,19世纪末期,女权主义运动开始在英国兴起,“新女性”开始出现。“新女性”一词由作家亨利·詹姆斯创造,用以形容当时欧洲和美国急剧增加的受过教育、经济独立、有强烈自我意识的年轻女性。她们不愿再做主流社会期待的“家庭天使”,而是要求男女平等,并大胆表露感情和性欲。这种全新大胆的女性形象毋庸置疑给当时的男权社会造成了极大冲击。学者坎宁安认为,使“新女性”成为危险形象的关键因素是性。
[4]East E M.Heterosis.Genetics,1936,21:375-397
[5]Yu S B,Li J X,Xu C G,Yan Y F,Gao Y J.Importance of epistasis as the genetic basis of the heterosis in an elite rice hybrid.Proc Natl Acad Sci USA,1997,94:9226-9231
[6]Song R T,Messing J.Gene expression of a gene family in maize based on noncollinear haplotypes.Proc Natl Acad Sci USA,2003,100:9055-9060
[7]Hoecker N,Keller B,Muthreich N,Chollet D,Descombes P,Piepho H P,Hochholdinger F.Comparison of maize (Zea mays L.) F1-hybrid and parental inbred line primary root transcription suggests organ-specific patterns of nonadditive gene expression and conserved expression trends.Genetics,2008,179:1275-1283
[8]Fu Z Y,Jin X N,Ding D,Li Y L,Fu Z J,Tang J H.Proteomic analysis of heterosis during maize seed germination.Proteomics,2011,11:1462-1472
[9]Ding D,Wang Y J,Han M S,Fu Z Y,Li W H,Liu Z H,Hu Y M,Tang J H.MicroRNA transcriptomic analysis of heterosis during maize seed germination.PLoS One,2012,7(6):e39578
[10]Guo M,Rupe M A,Wei J,Winkler C,Goncalves-Butruille M,Weers B P,Cerwick S F,Dieter J A,Duncan K E,Howard R J,Hou Z,Löffler C M,Cooper M,Simmons C R.Maize ARGOS1 (ZAR1) transgenic alleles increase hybrid maize yield.J Exp Bot,2014,65:249-260
[11]严建兵,汤华,黄益勤,石永刚,李建生,郑用琏.不同发育时期玉米株高QTL的动态分析.科学通报,2003,48:1959-1964 Yan J B,Tang H,Huang Y Q,Si Y G,Li J S,Zheng Y L.Dynamic QTL analysis for plant height in different developing stages in maize.Chin Sci Bull,2003,48:1959-1964 (in Chinese with English abstract)
[12]Li Z K,Luo L J,Mei H W,Wang D L,Shu Q Y,Tabien R,Zhong D B,Ying C S,Stansel J W,Khush G S,Paterson A H.Overdominance epistatic loci are the primary genetic basis of inbreeding depression and heterosis in rice:I.Biomass and grain yield.Genetics,2001,158:1737-1753
[13]Kusterer B,Muminovic J,Utz H F,Piepho H P,Barth S,Heckenberger M,Meyer R C,Altmann T,Melchinger A E.Analysis of a triple testcross design with recombinant inbred lines reveals a significant role of epistasis in heterosis for biomass-related traits in Arabidopsis.Genetics,2007,175:2009-2017
[14]Kusterer B,Piepho H P,Utz H F,Muminovic J,Meyer R C,Altmann T,Melchinger A E.Heterosis for biomass related traits in Arabidopsis investigated by a novel QTL analysis of the triple testcross design with recombinant inbred lines.Genetics,2007,177:1839-1850
[15]Hua J,Xing Y,Wu W,Xu C,Sun X,Yu S B,Zhang Q F.Single-locus heterotic effects and dominance by dominance interactions can adequately explain the genetic basis of heterosis in an elite rice hybrid.Proc Natl Acad Sci USA,2003,100:2574-2579
[16]Xiao J H,Li J M,Yuan L P,Tanksley S D.Dominance is the major genetic basis of the heterosis in rice as revealed by QTL analysis using molecular markers.Genetics,1995,140:745-754
[17]Lu H,Romero-Severson J,Bernarbo R.Genetic basis of heterosis explored by simple sequence repeat markers in a random-mated maize population.Theor Appl Genet,2003,107:494-502
[18]Semel Y,Nissenbaum J,Menda N,Zinder M,Krieger U,Issman N,Pleban T,Lippman Z,Gur A,Zamir D.Overdominant quantitative trait loci for yield and fitness in tomato.Proc Natl Acad Sci USA,2006,103:12981-12986
[19]Krieger U,Lippman Z B,Zamir D.The flowering gene single flower truss drives heterosis for yield in tomato.Nat Genet,2010,42:459-463
[20]Wang Z Q,Yu C Y,Liu X,Liu S J,Yin C B,Liu L L,Lei J G,Jiang L,Yang C,Chen L M,Zhai H Q,Wan J M.Identification of indica rice chromosome segments for the improvement of japonica inbreds and hybrids.Theor Appl Genet,2012,124:1351-1364
[21]Meyer R C,Kusterer B,Lisec J,Steinfath M,Becher M,Scharr H,Melchinger A E,Selbig J,Schurr U,Willmitzer L,Altmann T.QTL analysis of early stage heterosis for biomass in Arabidopsis.Theor Appl Genet,2010,120:227-237
[22]Guo X,Guo Y,Ma J,Wang F,Sun M,Gui L J,Zhou J J,Song X L,Sun X Z,Zhang T Z.Mapping heterotic loci for yield and agronomic traits using chromosome segment introgression lines in cotton.J Integr Plant Biol,2013,55:759-774
[23]Stuber C W,Lincoln S E,Wolff D W,Helentjaris T,Lander E S.Identification of genetic factors contributing to heterosis in a hybrid from two elite maize inbred lines using molecular markers.Genetics,1992,132:823-839
[24]王懿波,王振华,王永普,张新,陆利行.中国玉米主要种质杂交优势利用模式研究,中国农业科学,1997,30(4) :16-24 Wang Y B,Wang Z H,Wang Y P,Zhang X,Lu L X.Studies on the heterosis utilizing models of main maize germplasms in China.Sci Agric Sin,1997,30(4):16-24 (in Chinese with English abstract)
[25]滕文涛,曹靖生,陈彦惠,刘向辉,景希强,张发军,李建生.十年来中国玉米杂种优势群及其模式变化的分析.中国农业科学,2004,37:1804-1811 Teng W T,Cao J S,Chen Y H,Liu X H,Jing X Q,Zhang F J,Li J S.Analysis of maize heterotic groups and patterns during pastdecade in China.Sci Agric Sin,2004,37:1804-1811 (in Chinese with English abstract)
[26]袁亮,丁冬,李卫华,谢惠玲,汤继华,付志远.玉米优良自交系单片段代换系的构建.玉米科学,2012,20(2):52-55 Yuan L,Ding D,Li W H,Xie H L,Tang J H,Fu Z Y.Construction of single segment substitution lines (SSSLs) of the elite inbred lines in maize.J Maize Sci,2012,20(2):52-55 (in Chinese with English abstract)
[27]Duvick D N.Biotechnology in the 1930s:the development of hybrid maize.Nature,2001,2:69-74
[28]Tang J H,Ma X Q,Teng W T,Yan J B,Wu W R,Dai J R,Li J S.Detection of quantitative trait loci and heterosis for plant height in maize in ‘‘immortalized F2’’ (IF2) population.Chin Sci Bull,2006,51:2864-2869
[29]Wei X Y,Wang B,Peng Q,Wei F,Mao K J,Zhang X G,Sun P,Liu Z H,Tang J H.Heterotic loci for various morphological traits of maize detected using a single segment substitution lines test-cross population.Mol Breed,2015,35(3):1-13
[30]Tang J H,Yan J B,Ma X Q,Teng W T,Dai J R,Dhillon B S,Melchinger A E.Dissection of the genetic basis of heterosis in an elite maize hybrid by QTL mapping in an “immortalized F2”population.Theor Appl Genet,2010,120:333-340
[31]王懿波,王振华,陆利行,王永普,张新,田曾元.中国玉米种质基础、杂种优势群划分与杂种优势模式研究.玉米科学,1998,6(1):9-13 Wang Y B,Wang Z H,Lu L H,Wang Y P,Zhang X,Tian Z Y.Studies on maize germplasm base,division of heterosis groups and utilizing models of heterosis in China.J Maize Sci,1998,6(1):9-13 (in Chinese with English abstract)
[32]吴金凤,宋伟,王蕊,田红丽,李雪,王凤格,赵久然,蔚荣海.利用SNP标记对51份玉米自交系进行类群划分.玉米科学,2014,22(5):29-34 Wu J F,Song W,Wang R,Tian H L,Li X,Wang F G,Zhao J R,Wei R H.Heterotic grouping of 51 maize inbred lines by SNP markers.J Maize Sci,2014,22(5):29-34 (in Chinese with English abstract)
URL:http://www.cnki.net/kcms/detail/11.1809.S.20160126.1559.004.html
Identification of Heterotic Loci for Yield and Ear Traits Using CSSL Test Population in Maize
PENG Qian**,XUE Ya-Dong**,ZHANG Xiang-Ge,LI Hui-Min,SUN Gao-Yang,LI Wei-Hua,XIE Hui-Ling,and TANG Ji-Hua*
Key Laboratory of Wheat and Maize Crops Science/Collaborative Innovation Center of Henan Grain Crops/College of Agronomy,Henan Agricultural University,Zhengzhou 450002,China
Abstract:Heterosis plays an important role in enhancing crop yield and quality.Dissecting the genetic basis of heterosis can promote hybrid maize selection,however it is unclear up to now.In this study,a set of chromosome segment substitution lines (CSSLs) population,which was constructed using the inbred line lx9801 as the receptor parent and the inbred line Chang 7-2 as the donor parent,was crossed with the inbred line T7296 to construct the corresponding test population.The test population was used to identify the heterotic loci (HL) for grain yield and ear traits in maize,which showed significant difference in heterosis between the corresponding chromosomal region of the inbred line Chang 7-2 and lx9801 as well as the test inbred line T7296.A total of 64 HL were identified for gain yield and ear traits,and among them 23 HL were identified at the two environments simultaneously,including 4 HL for ear length,4 HL for ear width,4 HL for row number,7 HL for kernels per row,and 4 HL for grain yield.Additionally,the HL for both grain yield and its components simultaneously were found on many chromosomal regions.This study could offer a basic material for thoroughly dissecting the genetic basis of heterosis for grain yield and its components in maize.
Keywords:Maize;Chromosome segment substitution lines;Grain yield;Heterosis;Quantitative trait loci
收稿日期Received():2015-07-04;Accepted(接受日期):2016-01-11;Published online(网络出版日期):2016-01-26.
*通讯作者(
Corresponding author):汤继华,E-mail:tangjihua1@163.com,Tel:0371-63558377
DOI:10.3724/SP.J.1006.2016.00482
