高温对蓝莓叶片气孔特征和气体交换参数的影响
2016-04-09党承华王贺新姜国斌李根柱张自川郑云普大连大学生命科学与技术学院大连1166大连大学现代农业研究院大连1166大连民族大学环境与资源学院大连116600河北工程大学水电学院邯郸05608河北省水生态文明及社会治理研究中心邯郸05608
朱 玉,黄 磊,党承华,王贺新,姜国斌,李根柱,张自川,娄 鑫,郑云普,5※(1.大连大学生命科学与技术学院,大连1166;.大连大学现代农业研究院,大连1166;.大连民族大学环境与资源学院,大连116600;.河北工程大学水电学院,邯郸05608;5.河北省水生态文明及社会治理研究中心,邯郸05608)
高温对蓝莓叶片气孔特征和气体交换参数的影响
朱玉1,2,黄磊3,党承华4,王贺新2,姜国斌3,李根柱2,张自川2,娄鑫2,郑云普4,5※
(1.大连大学生命科学与技术学院,大连116622;2.大连大学现代农业研究院,大连116622;3.大连民族大学环境与资源学院,大连116600;4.河北工程大学水电学院,邯郸056038;5.河北省水生态文明及社会治理研究中心,邯郸056038)
摘要:为探讨不同强度高温对南高丛蓝莓叶片气孔特征及其气体交换参数的影响,利用人工气候箱设置4个温度处理:对照(25℃)、轻度高温(30℃)、中度高温(35℃)和重度高温(40℃)对两年生南高丛蓝莓(Vaccinium corymbosum L.)幼苗(海岸、奥尼尔及蓝脊)进行为期90 d的光照培养实验。研究结果表明:高温增加海岸和蓝脊叶片的气孔密度,但对奥尼尔的气孔密度无影响。中度高温增大奥尼尔和蓝脊叶片气孔的长度、宽度和面积,但海岸的气孔长度比轻度高温减小23.5% (P<0.05)。高温使奥尼尔的气孔空间分布更加规则,而对海岸和蓝脊的影响不大。3个品种的叶片净光合反应速率(Pn)、气孔导度(Gs)和蒸腾速率(Tr)均随高温强度先升高后降低,但其最大值随品种发生变化。研究结果表明,南高丛蓝莓具有调整气孔结构特征和优化气孔空间分布格局提高其气体交换效率的功能,但在品种间存在较大的差异,最终导致气体交换参数对高温产生不同的响应,尤其表现在抵抗极端高温能力方面。结果有助于从叶片气孔特征变化角度深入理解不同高温强度对南高丛蓝莓气体交换产生影响的潜在机理,为蓝莓耐高温胁迫选育及引种栽培工作提供理论支持。
关键词:温度;胁迫;优化;蓝莓;高温胁迫;气孔结构和功能;气孔分布格局;气体交换
朱玉,黄磊,党承华,王贺新,姜国斌,李根柱,张自川,娄鑫,郑云普.高温对蓝莓叶片气孔特征和气体交换参数的影响[J].农业工程学报,2016,32(01):218-225.doi:10.11975/j.issn.1002-6819.2016.01.031 http://www.tcsae.org Zhu Yu, Huang Lei, Dang Chenghua, Wang Hexin, Jiang Guobin, Li Genzhu, Zhang Zichuan, Lou Xin, Zheng Yunpu.Effects of high temperature on leaf stomatal traits and gas exchange parameters of blueberry[J].Transactions of the Chinese Society of Agricultural Engineering(Transactions of the CSAE), 2016, 32(01): 218-225.(in Chinese with English abstract)doi:10.11975/j.issn.1002-6819.2016.01.031 http://www.tcsae.org
0 引言
气孔是植物与大气之间进行气体交换的重要器官[1-2],对调节生态系统碳、水循环过程起着极其重要的作用[3-5]。植物一般通过调整叶片气孔的开度、大小、频度及其空间格局来优化气体交换的效率。然而,目前有关高温对叶片气孔频度及大小等方面特征的影响并没有一致的结论。例如,一些研究的结果发现,高温没有改变叶片的气孔密度和气孔指数[6-8],但有些研究却发现高温能够减少或增加气孔密度和气孔指数[9-12]。此外,高温还能改变单个气孔的大小和形状[13-14]及其在叶片上的空间分布格局[15-16]。同时,叶片的光合作用过程对高温的反应也最为敏感,在其他胁迫症状出现前被完全抑制[17]。然而,以往的研究结果关于高温对叶片光合速率的影响还存在较大的争议[6,18]。有研究报道高温对叶片光合速率没有影响[19],而另一些研究却发现高温降低了叶片的光合速率[18,20-23],甚至还有的研究认为高温会增加叶片的光合速率[11,24-25]。对以往不同的研究结果进行比较是非常困难的,这主要是由于以往研究中的高温强度存在较大差异,且光合作用的温度敏感性和最适温度在不同物种和生态型之间也不尽相同[26]。通常而言,当高等植物生存在极端高温或者低温环境下时,植物光合作用的反应速率均比较低;而只有植物生长在某一个最适温度下才能表现出最大的光合反应速率[11,27]。
蓝莓(Vaccinium corymbosum L.)属杜鹃花科(Ericaceae)越桔属(Vaccinium)植物,其果实富含的花青素类物质(Anthocyanins)具有较强的抗氧化活性[28-31]。南高丛蓝莓是美国人工杂交选育的栽培品种,冬季冷温需要量低于500 h,比北高丛蓝莓更适宜在高温气候区种植。然而,据报道我国的气候呈现逐渐变暖的趋势,特别是华北和东北地区的升温幅度尤为明显[32-33]。随着全球变暖的加剧,近年来极端高温气候事件的发生频率也明显增加[34],导致局部区域气温较长时间持续达到40℃以上[35-36]。此外,相关的研究结果还发现高温通过改变植物的气体交换过程(如光合作用)直接影响植物的生长和发育进程[24,37-42]。然而,高温是否会影响南高丛蓝莓叶片气孔特征及其气体交换过程,最终影响蓝莓的生长状况这一关键科学问题至今未得到较好的回答。本研究探讨高温对南高丛蓝莓叶片气孔特征(气孔大小、气孔密度和气孔空间分布格局)及其气体交换过程的影响,重点从气孔的角度深入剖析和理解高温对南高丛蓝莓叶片气体交换过程产生影响的潜在机理,为南高丛蓝莓耐高温的选育及引种提供理论依据和数据支持。
1 材料与方法
1.1试验材料及处理
以南高丛蓝莓奥尼尔、海岸和蓝脊为试验材料,每个品种随机挑选长势和大小一致的2年生幼苗20株,平均分配到4个人工气候箱中进行30 d的适应性预培养。所有的人工气候箱均设置温度25℃(昼)/ 20℃(夜),光照强度1 000 μmol/m2·s,光照周期8:00~20:00,相对湿度60%~75%。每周浇水和Hoagland营养液各1次。将上述4个人工气候箱的温度分别设定为25℃(对照)、30℃(轻度高温)、35℃(中度高温)和40℃(重度高温),其它环境参数完全相同,对幼苗进行90 d的高温处理培养,且每周对植株浇水和Hoagland营养液各1次。为避免气候箱自身差异对研究结果产生影响,培养期内每周随机调换气候箱的温度和植株。
1.2印迹法取样及光学显微镜观察
每株选取3个叶片,用无色透明的指甲油涂于叶片远轴面的中部,采集气孔印迹样品(5mm×15mm)。将叶片印迹玻片置于莱卡光学显微镜(DM2500,Leica Corp,Germany)下观察并利用显微镜装备的照相机(DFC 300-FX, Leica Corp, Germany)拍照。选择5个随机的显微视野,每个视下拍3张照片,得到15张气孔的显微照片(面积0.3 mm× 0.45 mm),从15张照片中随机选取5张来计算气孔的密度。另外,随机选取4张照片,利用ArcGIS10.0软件分别测量气孔的长、宽、周长、面积及形状指数[25]。形状指数是指通过计算单一气孔形状与相同面积的圆之间的偏离程度来测量其形状的复杂程度,即当气孔为圆形时,其形状指数即为1;若气孔的形状越扁长,则气孔形状指数的值就越大。形状指数S的计算公式为:

式中S为气孔的形状指数;P为气孔开口的周长,mm;A为气孔开口的面积,mm2。
1.3扫描电镜观察
从叶片中部随机采集3个2 mm×2 mm样品。将样品固定于2.5%(v/v)的戊二醛(0.1 mol/L磷酸缓冲液,pH值7.0),置于4℃下冷藏保存3 d,用磷酸缓冲液冲洗6次,用1%(v/v)锇酸固定3 h,用相同磷酸缓冲液冲洗干净。将组织进行不同酒精梯度的脱水和临界点干燥,再在观察台上固定,利用高压涂膜装置进行喷金处理。对单个气孔在Quanta 200扫描电子显微镜(FEI Corp, USA)下观察和拍照[25]。
1.4气孔的空间格局分析
本研究中,认为每个气孔都是叶片上分布的单点,气孔开口的最中间位置为单点。利用ArcGIS10.0软件将显微照片在相同坐标系进行数字化处理,得到每个气孔的坐标值,运用Ripley,s K-Function空间统计分析方法对气孔分布状况进行分析[25]。利用所有单点距离的二阶矩阵探究其在不同尺度上的二维分布格局。L(d)值表达为:

式中L(d)为最小邻域距离,mm;d为气孔空间分布尺度,mm。
为了确定95%的可信任区间,采用蒙特卡洛算法模拟随机分布点1 000次。当分布格局为随机分布时,所有的d值到L(d)距离均相等。若L(d)值大于95%可信任区间,气孔为簇状分布格局。如果气孔在给定尺度d下随机分布,则L(d)值在95%可信任区间内。反之,L(d)值小于95%可信任区间为规则分布[25]。
1.5气体交换参数测定
本研究中的叶片净光合速率(Pn)、气孔导度(Gs)、细胞间CO2浓度(Ci)和蒸腾速率(Tr)等气体交换参数使用光合测定系统(Li-6400;LI-COR Inc.Lincoln, Nebraska, USA)确定[26]。叶室内光照强度1000 μmol/m2·s,CO2浓度400 μmol/m2·s,光合有效辐射(photosynthetically available radiation,PAR),叶片温度为25℃水蒸汽压亏缺(water vapor pressure deficit,VPD)为2.0 kPa[26]。另外,光合反应速率测定结束后,关闭Li-6400便携式光合测定系统的光源,让叶片在黑暗环境下适应30 min后,测定叶片的暗呼吸速率Rd[38]。此外,叶片尺度的水分利用效率(water use efficiency,WUE)利用公式WUE=Pn/Tr来计算[18]。
1.6统计分析
本研究利用单因素方差分析和Duncan′s multiple range test比较处理间的显著性差异(P<0.05),利用SPSS 13.0(Chicago, IL)统计分析和Sigmaplot作图。
2 结果与分析
2.1高温对叶片气孔密度的影响
不同高温强度对3个南高丛蓝莓的气孔密度产生很大影响,且品种间存在较大的差异。各温度处理对奥尼尔叶片上气孔的密度没产生显著影响(P>0.05)见表1。高温处理使海岸的气孔密度先增加后降低,同对照相比,中度高温导致海岸的气孔密度增加29.8%(表1),而继续升高温度(重度高温)却使其气孔密度显著降低,比中度高温降低39.3%(P<0.05)。虽然高温也导致蓝脊的气孔密度呈现先增后降的趋势,但各处理间差异不显著(P>0.05)。
2.2高温对叶片气孔开度和形状的影响
奥尼尔叶片上气孔的长度、宽度、面积和周长均随高温强度的增加呈现升高的趋势,但其最大值仍低于对照,且各处理间差异不显著;另外,在中度高温条件下,奥尼尔叶片的气孔形状指数最小。不同高温处理对海岸的气孔长度、面积及周长产生显著影响(P<0.05)见表1,呈现先降后升的趋势,在中度高温处理时最低;同对照相比,中度高温使海岸的气孔长度、面积及周长分别降低29.5%、43.2%和27%;虽然各温度处理对气孔宽度影响不显著(P>0.05),但仍是中度高温时气孔宽度最小。在中度高温时海岸叶片上气孔形状指数最低,而重度高温时升高,同中度高温相比,重度高温使气孔的形状指数显著增加约3%(P<0.05)(图1)。不同的高温处理使得蓝脊的气孔长度、面积和周长逐渐升高,且处理间差异显著(P<0.05),与对照相比,重度高温使气孔的长度、面积和周长分别增加18.7%、31.5%和14.8%;然而,蓝脊叶片上的气孔宽度则在中度高温时达到最大,分别比对照和轻度高温处理增加14.5%和19.3%;随着高温强度的继续加大(40℃),蓝脊气孔的宽度稍微降低,但与中度高温相比差异不显著(P>0.05)。另外,高温对蓝脊气孔的形状指数影响不显著,在中度高温条件下,气孔的形状指数最小(表1)。
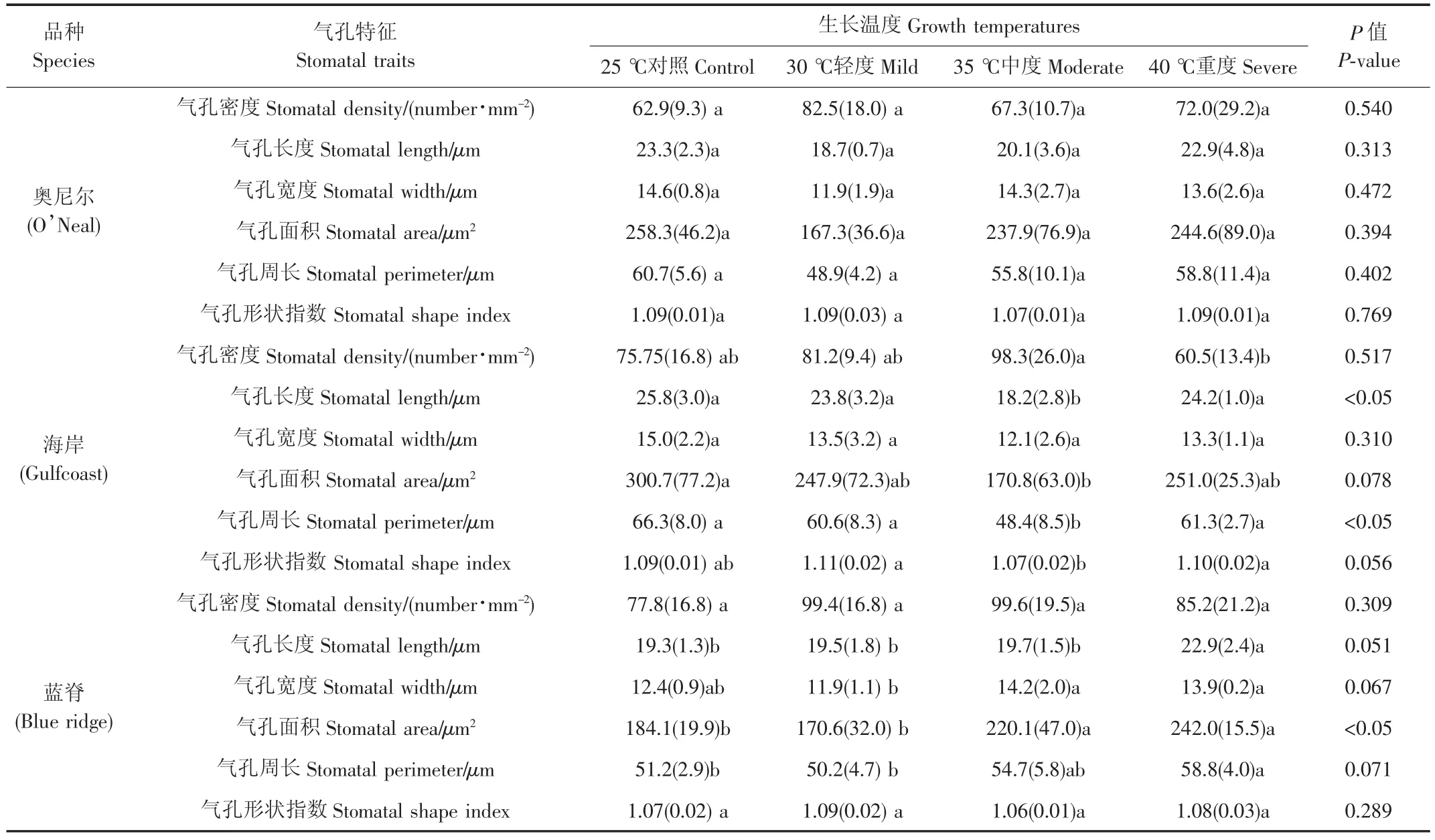
表1 高温对南高丛蓝莓叶片气孔特征的影响Table 1 Effects of high temperatures on leaf stomatal traits of south highbush blueberry
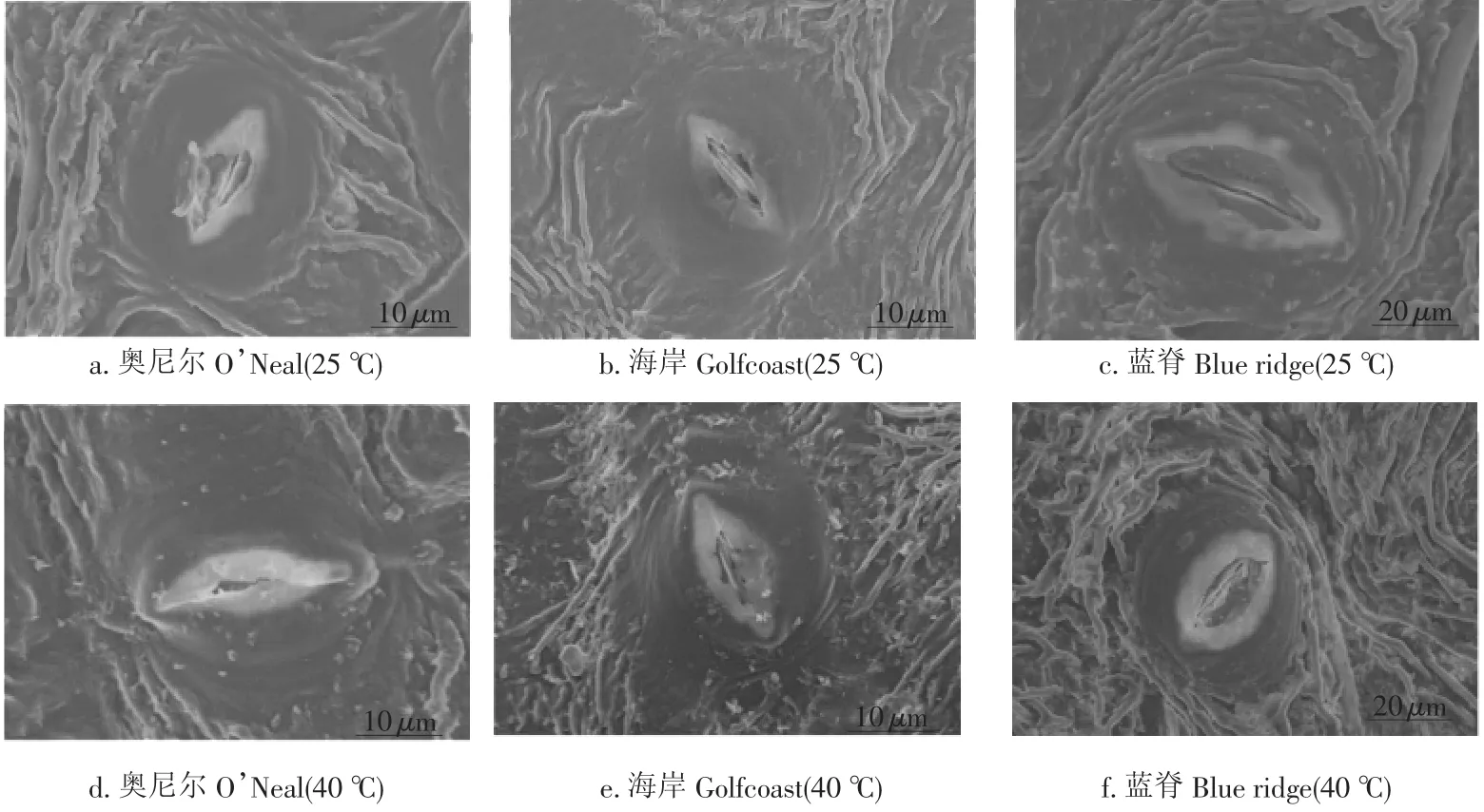
图1 南高丛蓝莓叶片单个气孔结构特征的扫描电子显微镜照片Fig.1 Scanning electron photographs of structural traits of individual stoma on south highbush blueberry leaves
2.3高温对气孔空间分布格局的影响
不同的高温强度下,3个品种的气孔均在小尺度范围内为规则分布(<162 μm),而在大尺度范围内随机分布(表2)。具体来说,与对照相比,重度高温使奥尼尔的空间尺度减少11.9%(表2),L(d)值减小14%(表3)。不同强度的高温处理对海岸叶片的气孔空间分布格局没有显著影响(P>0.05;表2和3)。此外,随着高温强度的增加蓝脊叶片上气孔的平均空间尺度和L(d)值均逐渐升高,在轻度高温时最小,而重度高温时最大,平均的空间尺度和L(d)值分别比轻度高温时增加17.8%和21.6%(表2、3)。
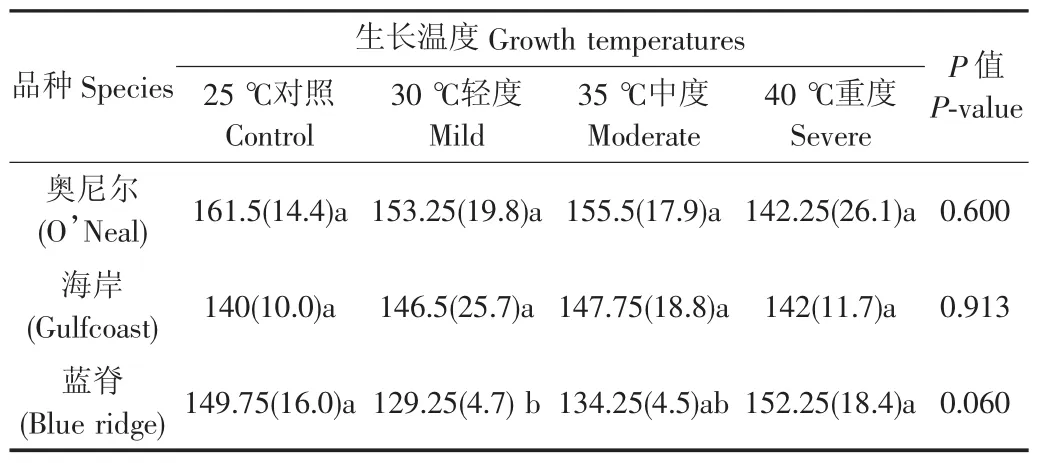
表2 高温对南高丛蓝莓叶片气孔空间分布尺度的影响Table 2 Effects of high temperatures on spatial distribution scales of stomata on the leaves of south highbush blueberries

表3 高温对南高丛蓝莓叶片单个气孔间最小邻域距离的影响Table 3 Effects of high temperatures on nearest neighbor distance between stomata on leaves of south highbush blueberries
2.4高温对叶片气体交换参数的影响
高温导致3个南高丛蓝莓叶片的净光合反应速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)均先升高后降低,但其最大值随品种的不同而变化(图2)。3个品种在中度增温时Pn最大,依次为奥尼尔>蓝脊>海岸。随后,奥尼尔的Pn明显降低,在重度高温时,奥尼尔、海岸和蓝脊叶片的Pn仍然比对照高2.02、1.44和1.61倍。另外,轻度高温导致海岸和蓝脊叶片Gs的最大值比对照提高2.43倍和1.80倍,中度高温使奥尼尔叶片Gs最大值比对照提高2.41倍。同时,轻度高温也使海岸和蓝脊叶片Tr的最大值比对照分别增加约1倍和1.5倍,中度高温使奥尼尔叶片Tr的最大值比对照增加约6倍。此外,高温导致奥尼尔的水分利用效率(WUE)呈现先升高后降低的趋势,轻度高温时WUE达到最大值,重度高温使WUE比对照显著降低2.75倍,海岸和蓝脊则是在中度高温时WUE达到最大值。然而,细胞间CO2浓度(Ci)却表现出相反的变化趋势,即不同高温强度导致3个蓝莓的Ci先降低后升高,中度高温下海岸、奥尼尔和蓝脊的Ci出现最小值,且比对照分别降低17%、27%和32%。同时,3个蓝莓叶片的暗呼吸速率始终随着高温强度的升高而增加,在重度高温时达到最大值;同对照相比,海岸、奥尼尔和蓝脊叶片的Rd分别提高约2倍、2.5倍和3倍(图2)。

图2 高温对南高丛蓝莓叶片气体交换的影响Fig.2 Effects of high temperatures on leaf gas exchange of south highbush blueberries
3 讨论
3.1高温对南高丛蓝莓叶片气孔特征的影响
目前有关高温对叶片气孔频度及大小等方面特征的影响还没有一致的结论[43-47]。本研究结果显示,中度高温导致海岸和蓝脊的气孔密度最大,但气孔开口没有达到最大(表1),表明中度高温条件下,虽然高温使海岸和蓝脊叶片上的气孔密度增加,但并未对气孔的开度产生影响,即叶片上气体交换的气孔有效面积未达到最大值。尽管高温对奥尼尔叶片的气孔密度没有显著影响,但中度高温使叶片气孔的开度增加,此时叶片上气体交换的有效面积显著增加。另外,中度高温时奥尼尔的气孔导度和蒸腾速率均具有较高的数值也直接支持了上述的结论。然而,虽然重度高温时奥尼尔叶片的气孔长度增加,气孔的宽度减小,即气孔变得更扁,表明重度高温时植物减小气孔开度来降低由叶片内水分过度散失而带来的伤害。另外,扫描电子显微镜的观察结果也证实重度高温条件下奥尼尔叶片上的气孔与对照相比开口变小、气孔关闭(图2)。重度高温环境下叶片蒸腾速率的降低表明叶片水分散失的减少,这可能是植物为了抵抗高温胁迫而对气孔密度和开度进行调整的具体表现。然而,植物通过调整气孔大小来应对外界高温环境的能力在不同品种间存在着很大的差异。中度高温处理对奥尼尔叶片的气孔密度、气孔长度、气孔宽度以及气孔周长和面积均没有产生显著的影响,但气孔的形状指数却最小(气孔最圆),表明奥尼尔具有通过优化气孔结构来抵抗外界高温环境的能力。研究结果表明,南高丛蓝莓可以通过调整叶片的气孔结构特征(气孔密度和气孔开度)来最优化其进行气体交换的效率,从而降低高温胁迫伤害,但该抗高温的能力在不同品种间存在变异(奥尼尔>海岸>蓝脊)。
3.2高温对南高丛蓝莓叶片气孔空间分布格局的影响
以往的研究结果发现植物叶片上气孔的空间分布格局由细胞的分裂和分化过程所决定,而细胞的分裂和分化过程会受到遗传信号的调控[16,48-52]和环境因素的影响[51,53]。与对照相比,重度高温使奥尼尔的L(d)值减少14%,表明高温导致奥尼尔叶片气孔的空间分布格局更加规则,从而有利于提高奥尼尔叶片的气体交换效率(表4)。此外,随着温度的增高,平均L(d)值比轻度高温时增加21.6%(表3),表明高温可能导致蓝脊叶片上的气孔分布更加不规则,从而使植物气体交换的速率降低。
3.3高温对南高丛蓝莓叶片气体交换参数的影响
光合作用是植物对环境温度变化响应最为敏感的生理过程之一[54-60]。本研究的结果显示,在中度高温条件下(35℃)奥尼尔叶片上气孔的开度较大且气孔形状指数最小,即气孔开口的形状接近于圆形(表1),表明气孔开度的增大可能更加有利于大气CO2通过气孔进入叶片的内部,并扩散到相应的光合作用位点,从而提高蓝莓叶片的净光合速率。随着温度的增加,奥尼尔叶片上的气孔空间分布格局更加规则,表明更多的CO2分子易于由规则的气孔扩散到叶片组织内部,更加利于提高其净光合反应速率。此外,对奥尼尔叶片气体交换参数的测量结果表明,Pn在中度高温条件下的显著提高也为上述结论提供了直接的证据支持(图2a)。然而,与中度高温相比,尽管重度高温条件下奥尼尔叶片的气孔长度以及气孔面积和周长均增加,但气孔的形状指数增大(气孔变得扁而长),表明气孔的开度减小,且气孔接近关闭的状态;因此,植物为防止高温对光合反应位点造成伤害通过减小气孔的开度来控制CO2进入叶片的一种保护策略,最终导致Gs和Pn的降低(图1)。综上所述,南高丛蓝莓在应对高温胁迫时采取了多种策略,从叶片结构和功能的不同水平都做出了调整,以有利于其进行正常的生理代谢和生长;然而,南高丛蓝莓在叶片水平上做出调整应对高温环境的能力是十分有限的,轻度和中度高温条件下能保证植物正常的生长和代谢过程,但重度高温对3个南高丛蓝莓品种均造成不同程度的伤害。因此,未来气候变暖背景下南高丛蓝莓的种植仍存在高温气候事件伤害的潜在风险。
4 结论
本研究探讨了高温对3种南高丛蓝莓叶片气孔特征及其气体交换参数的影响,得到如下结论:
1)轻度和中度高温环境下的南高丛蓝莓通过优化叶片气孔的开度和分布提高气体交换效率。
2)重度高温时南高丛蓝莓调整气孔的开度或气孔密度来控制叶片组织内的CO2浓度,从而降低光合反应位点受高温伤害程度。
3)南高丛蓝莓通过调整气孔特征抵抗高温胁迫,不同品种抗高温胁迫的能力为奥尼尔>海岸>蓝脊。
研究结果有助于从气孔特征角度深入理解蓝莓耐高温胁迫的潜在机理,为抗高温蓝莓的引种和栽培提供理论支持和数据支撑。
[参考文献]
[1] Woodward F I.Stomatal numbers are sensitive to increases in CO2from preindustrial levels[J].Nature, 1987, 327: 617-618.
[2] Hetheringto A M, Woodward F I.The role of stomata in sensing and driving environmental change [J].Nature, 2003, 424: 901-908.
[3] Franks P J, Beerling D J.Maximum leaf conductance driven by CO2effects on stomatal size and density over geologic time[J].Proceedings of the National Academy of Sciences, 2009, 106: 10343-10347.
[4] Haworth M, Heath J, Mcelwain J C.Differences in the response sensitivity of stomatal index to atmospheric CO2among four genera of Cupressaceae conifers[J].Annals of Botany, 2010, 105: 411-418.
[5] Taylor S H, Franks P J, Hulme S P, et al.Photosynthetic pathway and ecological adaptation explain stomatal trait diversity amongst grasses[J].New Phytologist, 2012, 193: 387-396.
[6] Apple M E, Olszyk D M, Ormrod D P, et al.Morphology and stomatal function of Douglas fir needles exposed to climate change: Elevated CO2and temperature[J].International Journal of Plant Science, 2000, 161: 127-132.
[7] Kouwenberg L L R, Kurschner W M, Mcelwain J C.Stomatal frequency change over altitudinal gradients: Prospects for paleoaltimetry[J].Reviews in Mineralogy & Geochemistry, 2007, 66: 215-241.
[8] Fraser L H, Greenall A, Carlyle C, et al.Adaptive phenotypic plasticity of Pseudoroegneria spicata: response of stomatal density, leaf area and biomass to changes in water supply and increased temperature[J].Annals of Botany, 2009, 103: 769-775.
[9]张洁,李天来.日光温室亚高温对番茄光合作用及叶绿体超微结构的影响[J].园艺学报,2005,32(4):614-619.Zhang Jie, Li Tianlai.Effects of daytmie sub-high temperature on photosynthesis and chloroplast ultrastructure of tomato leaves in greenhouse[J].Acta Horticulturae Sinica, 2005, 32(4): 614-619.(in Chinese with English abstract)
[10] Xu Z Z, Zhou G S, Shimizu H.Effects of soil drought with nocturnal warming on leaf stomatal traits and mesophyll cell ultrastructure of a perennial grass[J].Crop Science, 2009, 49: 1843-1851.
[11] Jin B, Wang L, Wang J, et al.The effect of experimental warming on leaf functional traits, leaf structure and leaf biochemistry in Arabidopsis thaliana[J].BMC Plant Biology, 2011, 11: 35.
[12] Hu J, YangY Q, Huang W, et al.Effects of temperature on leaf hydraulic architecture of tobacco plants [J].Planta, 2014, 240: 489-496.
[13]张立荣,牛海山,汪诗平,等.增温与放牧对矮嵩草草甸4种植物气孔密度和气孔长度的影响[J].生态学报,2010,30 (24):6961-6969.Zhang Lirong, Niu Haishan, Wang Siping, et al.Effects of temperature increase and grazing on stomatal density and length of four alpine Kobresia meadow species, Qinghai-Tibetan Plateau[J].Acta Ecologica Sinica, 2010, 30(24): 6961-6969.(in Chinese with English abstract)
[14] Chen W L, Yang W J, Lo H F, et al.Physiology, anatomy, and cell membrane thermostability selection of leafy radish (Raphanus sativus var.oleiformis Pers.)with different tolerance under heat stress[J].Scientia Horticulturae, 2014, 179: 367-375.
[15] Croxdale J L.Stomatal patterning in angiosperms[J].American Journal of Botany, 2000, 87: 1069-1080.
[16] Shpak E D, Mcabee J M, Pillitteri L J, et al.Stomatal patterning and differentiation by synergistic interactions of receptor kinases [J].Science, 2005, 309: 290-293.
[17] Berry J, Bjorkman O.Photosynthetic response and adaptation to temperature in higher plants [J].Annual Review of Plant Physiology, 1980, 31: 491-543.
[18] Djanaguiraman M, Prasad P V V, Boyle D L, et al.Hightemperature stress and soybean leaves: Leaf anatomy and photosynthesis[J].Crop Science, 2011, 51: 2125-2131.
[19] Llorens L, Penuelas J, Estiarte M, et al.Contrasting growth changes in two dominant species of a Mediterranean shrubland submitted to experimental drought and warming [J].Annals of Botany, 2004, 94: 843-853.
[20]郭延平,周慧芬,曾光辉,等.高温胁迫对柑橘光合速率和光系统Ⅱ活性的影响[J].应用生态学报,2003,14(6):867-870.Guo Yanping, Zhou Huifen, Zeng Guanghui, et al.Effects of high temperature stress on net photosynthetic rate and photosystemⅡactivity in Citrus [J].Journal of Applied Ecology, 2003, 14(6): 867-870.(in Chinese with English abstract)
[21]张富存,张波,王琴,等.高温胁迫对设施番茄光合作用特性的影响[J].中国农学通报,2011, 27(28): 211-216.Zhang Fuchun, Zhang Bo, Wang Qin, et al.Effects of high temperature stress on photosynthesis characters of tomato [J].Chinese Agricultural Science Bulletin, 2011, 27(28): 211-216.(in Chinese with English abstract)
[22]杜尧东,李键陵,王华,等.高温胁迫对水稻剑叶光合和叶绿素荧光特征的影响[J].生态学杂志,2012, 31(10):2541-2548.Du Yaodong, Li Jianling, Wang Hua, et al.Effects of high temperature stress on the flag leaf photosynthesis and chlorophyll fluorescence parameters of rice[J].Chinese Journal of Ecology, 2012, 31(10): 2541-2548.(in Chinese with English abstract)
[23] Feng B, Liu P, Li G, et al.Effect of heat stress on the photosynthetic characteristics in flag leaves at the grain-filling stage of different heat-resistant winter wheat varieties[J].Journal of Agronomy & Crop Science, 2014, 200: 143-155.
[24] Han C, Liu Q, Yang Y.Short-term effects of experimental warming and enhanced ultraviolet-B radiation on photosynthesis and antioxidant defense of Picea asperata seedlings[J].Plant Growth & Regulation, 2009, 58: 153-162.
[25] Zheng Y P, Xu M, Hou R X, et al.Effects of experimental warming on stomatal traits in leave s of maize(Zea may L.)[J].Ecology and Evolution, 2013, 3: 3095-3111.
[26] Niu S L, Wan S Q.Warming changes plant competitive hierarchy in a temperate steppe in northern China[J].Journal of Plant Ecology, 2008, 1: 103-110.
[27] Battaglia M, Beadle C, Loughhead S.Photosynthetic temperature responses of Eucalyptus globulus and Eucalyptus nitens[J].Tree Physiology, 1996, 16: 81-89.
[28] Bae J Y, Lim S S, Kim S J, et al.Bog blueberry anthocyanins alleviate photoaging in ultraviolet-B irradiation-induced human dermalfibroblasts[J].Molecular Nutrition&Food Research, 2009, 53: 726-738.
[29] Liu J, Zhang W, Jing H, et al.Bog bilberry(Vaccinium uliginosum L.)extract reduces cultured Hep-G2, Caco-2 and 3T3 -L1 cell viability, affects cell cycle progression and has variable effects on membrane permeability[J].Journal of Food Science, 2010, 75: 103-107.
[30] Li C Y, Feng J, Huang W Y, et al.Composition of polyphenols and antioxidant activity of rabbiteye blueberry(Vaccinium ashei)in Nanjing[J].Journal of Agriculture Food Chemistry, 2013, 61: 523-531.
[31] Wang L J, Su S, Wu J, et al.Variation of anthocyanins and flavonols in Vaccinium uliginosum berry in Lesser Khingan Mountains and its antioxidant activity[J].Food Chemistry, 2014, 160: 357-364.
[32] White T A, Campbell B D, Kemp P D.Impacts of extreme climatic events on competition during grassland invasions [J].Global Change Biology, 2001, 7: 1-13.
[33] Wand D, Heckathorn S A, Mainali K.Effects of N on plant response to heat-wave: A field study with prairie vegetation[J].Journal of Integrative Plant Biology, 2008, 50: 1416-1425.
[34] Meehl G A, Tebaldi C.More intense, more frequent, and longer lasting heat waves in the 21st century[J].Science, 2004, 305: 994-997.
[35] Schar C, Vidale P L, Luthi D, et al.The role of increasingtemperature variability in European summer heatwaves [J].Nature, 2004, 427: 332-336.
[36] Fischer E, Schar C.Consistent geographical patterns of changes in high-impact European heat waves[J].Nature Geosciences, 2010, 3: 398-403.
[37] Kudo G, Suzuki S.Warming effects on growth, production, and vegetation structure of alpine shrubs: a five-year experiment in northern Japan[J].Oecologia, 2003, 135: 280-287.
[38] Zhao C, Liu Q.Growth and physiological responses of Picea asperata seedlings to elevated temperature and to nitrogen fertilization[J].Acta Physiologia Plantarum, 2009, 31: 163-173.
[39] Prieto P, Penuelas J, Liusia J, et al.Effects of experimental warming and drought on biomass accumulation in a Mediterranean shrubland [J].Plant Ecology, 2009, 205: 179-191.
[40] Lin D, Xia J, Wan S.Climate warming and biomass accumulation of terrestrial plants: a meta-analysis[J].New Phytologist, 2010, 188: 187-198.
[41] Wang J, Duan B, Zhang Y.Effects of experimental warming on growth, biomass allocation, and needle chemistry of Abies faxoniana in even-aged monospecific stands[J].Plant Ecology,2012,213:47-55.
[42] Albert K R, Ro-Poulsen H, Mikkelsen T N, et al.Interactive effects of elevated CO2, warming, and drought on photosynthesis of Deschampsia-exuosa in a temperate heath ecosystem [J].Journal of Experimental Botany,2011, 62: 4253-4266.
[43] Young J J, Mehta S, Israelsson M, et al.CO2signaling in guard cells:Calciumsensitivityresponsemodulation,a Ca2+-independent phase, and CO2insensitivity of the gca2 mutant[J].Proceedings of National Academy of Sciences, 2006, 103: 506-7511.
[44] Shimazaki K, Doi M, Assmann S M, et al.Light regulation of stomatal movement[J].Annual Review of Plant Biology, 2007, 58: 219-247.
[45] Shang Z, Laohavisit A, Davies J M.Extracellular ATP activates an Arabidopsis plasma membrane Ca2+-permeable conductance [J].Plant Signaling & Behavior, 2009, 4: 989-991.
[46] Anderson V J, Brisk D D.Stomatal distribution, density and conductance of three perennial grasses native to the southern true trairie of Texas[J].American Midland Naturalist, 1990, 123: 152-159.
[47] Lammertsma E I, Boer H J, Dekker S C, et al.Global CO2rise leads to reduced maximum stomatal conductance in Florida vegetation[J].Proceedings of National Academy of Sciences, 2011, 108: 4035-4040.
[48] Franks P J, Drake P L, Beerling D J.Plasticity in maximum stomatal conductance constrained by negative correlation between stomatal size and density: an analysis using Eucalyptus globulus[J].Plant, Cell & Environment, 2009, 32: 1737-1748.
[49] Nadeau J A, Sack F D.Control of stomatal distribution on the Arabidopsis leaf surface[J].Science, 2002, 296: 1697-1700.
[50] Juarez M, Twigg R, Timmermans M.Specification of adaxial cell fate during maize leaf development[J].Development, 2004, 131: 4533-4544.
[51] Wang H, Ngwenyama N, Liu Y, et al.Stomatal development and patterning are regulated by environmentally responsive mitogenactived protein kinases in Arabidopsis[J].The Plant Cell, 2007, 19: 63-73.
[52] Hunt L, Baley K J, Gray J E.The signalling peptide EPFL9 is a positive regulator of stomatal development[J].New Phytologist, 2010, 186: 609-614.
[53] Casson S A, Gray J E.Influence of environmental factors on stomatal development[J].New Phytologist, 2008, 178: 9-23.
[54]王忠.植物生理学[M].北京:中国农业出版社,2006.Wang Zhong.Plant Physiology[M].Beijing: China Agriculture Press, 2006.(in Chinese with English abstract)
[55]张顺堂,张桂莲,陈立云,等.高温胁迫对水稻剑叶净光合速率和叶绿素荧光参数的影响[J].中国水稻科学,2011,25(3):335-338.Zhang Shuntang, Zhang Guilian, Chen Liyun, ett al.Effects of high temperature stress on the net photosynthetic rate and chlorophyll fluorescence parameters of rice flag leaves [J].Chinese Journal of Rice Science, 2011, 25(3): 335-338.(in Chinese with English abstract)
[56]罗海波,马苓,段伟,等.高温胁迫对‘赤霞珠’葡萄光合作用的影响[J].中国农业科学,2010,43(13):2744-2750.Luo Haibo, Ma Ling, Duan Wei, et al.Influence of Heat Stress on Photosynthesis in Vitis vinifera L.cv.Cabernet Sauvignon[J].Scientia Agricultura Sinica, 2010, 43(13): 2744 -2750.(in Chinese with English abstract)
[57]许大全.光合作用学[M].北京:科学出版社,2013.Xu Daquan.Photosynthesis[M].Beijing: Science Press, 2013.(in Chinese with English abstract)
[58]李天来,李淼.短期昼间亚高温胁迫对番茄光合作用的影响[J].农业工程学报,2009,25(9):220-225.Li Tianlai, Li Miao.Effect of short-term daytime sub-high temperature stress on photosynthesis of tomato leaves [J].Transactions of the Chinese Society of Agricultural Engineering (Transactions of the CSAE), 2009, 25(9):220-225.(in Chinese with English abstract)
[59]张洁,李天来,徐晶.昼间亚高温处理时期对日光温室番茄光合作用与产量的影响[J].农业工程学报,2008,24(3): 193-197.Zhang Jie, Li Tianlai, Xu Jing.Effects of sub-high temperature in daytime from different stages on tomato photosynthesis and yield in greenhouse[J].Transactions of the Chinese Society of Agricultural Engineering(Transactions of the CSAE), 2008, 24 (3): 193-197.(in Chinese with English abstract)
[60]梁文娟,王美玲,艾希珍,等.黄瓜幼苗光合作用对亚适温弱光胁迫的适应性[J].农业工程学报,2008,24(8):240-244.Liang Wenjuan, Wang Meiling, Ai Xizhen, et al.Photosynthesis adaptation of cucumber seedlings to suboptimal temperature and low light intensity stress[J].Transactions of the Chinese Society of Agricultural Engineering(Transactions of the CSAE), 2008, 24 (8): 240-244.(in Chinese with English abstract)
Effects of high temperature on leaf stomatal traits and gas exchange parameters of blueberry
Zhu Yu1,2, Huang Lei3, Dang Chenghua4, Wang Hexin2, Jiang Guobin3, Li Genzhu2, Zhang Zichuan2, Lou Xin2, Zheng Yunpu4,5※
(1.School of Life science and Technology, Dalian University, Dalian 116622, China; 2.Institute of Modern Agricultural Research, Dalian University, Dalian 116622, China; 3.College of Environment and Resources, Dalian Nationality University, Dalian 116600, China; 4.School of Water Conservancy and Hydropower, Hebei University of Engineering, Handan 056038, China; 5.Research Center of Water Ecological Civilization and Social Management, Handan 056038, China)
Abstract:Blueberry is one of the most important fruit trees all over the world and also the major economic tree species cultivated in China.Blueberry was suggested to be planted only in Northern China, because the warmer climate in Southern China could not meet its chilling duration.Several blueberry cultivars have recently been introduced to subtropical China and these cultivars can blossom and bear fruits.However, the temperature in subtropical China often approach 40℃or even higher in summer, and thus high temperature has become the most significant abiotic stress limiting the growth and production of blueberry in this area.Meanwhile, previous studies also reported that the heat endurance ability varied with different blueberry cultivars.In this study, we examined the effects of high temperatures on the leaf stomatal traits and gas exchange of blueberry cultivars with four growth chambers controlling different high temperatures including control(25℃), mild high temperature(30℃), moderate high temperature(35℃), and severe high temperature(40℃).Two-year-old seedlings of three highbush blueberry cultivars including‘O’Neal’,‘Gulfcoast’, and‘Blue Ridge’were selected from field plots and transplanted into pots(10 cm diameter×25 cm long)filled with fritted clay(one plant per pot)and grown in a greenhouse with an average temperature of 25/20℃(day/night)and about 1 000 μmol m-2·s-1photosynthetic active radiation(PAR)in natural sun light, and 60%~75% relative humidity for 30 d(March-April 2014)to establish canopy.During the establishment period, plants were irrigated to water-holding capacity daily and fertilized once per week with half-strength Hoagland's solution.We selected 20 healthy and uniform growth plants for each cultivar and then randomly planted the plants into each of four walk-in growth chambers(5 plants for each cultivar), where the temperature was determined as 25, 30, 35, or 40℃, respectively.Other environmental factors maintained throughout all the four chambers include humidity(60%~75%), light intensity(1 000 μmol m-2·s-1PAR), photoperiod(light on at 8 am, and off at 8 pm), soil type(fritted clay, same brand and package for all), water amount(200 ml, watered once per week), and nutrition type(plain tap water).Plants were fertilized once weekly with half-strength Hoagland's solution throughout the growth period.In order to minimize confounding effects of environmental variation between different chambers, we randomly changed the temperature of each growth chamber every week, and then we relocated the high temperature treated plants to the growth chambers with corresponding temperature.The large volume of the pot with frequent watering and fertilization ensured enough space for root growth and ample nutrient supply to avoid“bonsai effect”.Our results showed that comparing with the control, high temperatures increased the stomatal density of Gulfcoast and Blue Ridge, but had little effect on the stomatal density of O′Neal(P>0.05).Moderate high temperature increased the stomatal aperture width, stomatal aperture length, and stomatal area of O′Neal and Blue Ridge, whereas the stomatal aperture length of Gulfcoast was significantly reduced by 23.5% under moderate high temperature(P<0.05)compared with the mild high temperature(30℃).High temperatures resulted in a more regular stomatal distribution pattern on the leaves of O′Neal, while had little effect on the stomatal distribution pattern of Gulfcoast and Blue Ridge.Mild and moderate high temperatures significantly increased the net photosynthetic rates(Pn), stomatal conductance(Gs)and transpiration rates(Tr)of the three blueberry cultivars, whereas the severe high temperature caused them sharply decrease, although their maximum values varied across the three cultivars.In Conclusion, high temperatures increased the efficiency of leaf gas exchange by adjusting the structural characteristics and optimizing the spatial distribution pattern of stomata on the leaves of the south highbush blueberry.However, the ability of optimization for stomatal structure and function in the leaves of the south highbush blueberry was cultivar dependent, and thus resulted in the different responses to high temperatures in leaf gas exchange across cultivars, especially for heat stress resistance under extremely high temperature.Our results may not only be helpful for further understanding the potential mechanisms of high temperatures on leaf gas exchange of south highbush blueberry from the changes in leaf stomatal traits, but also provide theory for the selection and introduction of heat tolerance cultivars.
Keywords:temperature; stresses; optimization; blueberry; heat stress; stomatal structure and function; stomatal distribution pattern; leaf gas exchange
通信作者:※郑云普(1981-),男,河北邯郸人,讲师,博士,硕士生导师,主要从事农业水土环境及全球变化生态学方面的研究。邯郸河北工程大学水电学院,056038。Email:zhengyunpu_000@sina.com
作者简介:朱玉(1989-),女,河南周口人,主要从事蓝莓生理生态学及菌根学研究。大连大连大学生命科学与技术学院,116622。
基金项目:国家自然科学基金青年项目(31400418);辽宁省科技计划项目(2013204001);河北省自然科学基金项目(14964206D-3);中国博士后基金面上项目(2014M561044)
收稿日期:2015-08-01
修订日期:2015-12-02
中图分类号:S641
文献标志码:A
文章编号:1002-6819(2016)-01-0218-08
doi:10.11975/j.issn.1002-6819.2016.01.031
