New record of a haplocyonine amphicyonid in early Miocene of Nei Mongol fi lls a long-suspected geographic hiatus
2016-03-29WANgxiaomingWANGHongjiangjIANgZuOQigao
WANg xiao-mingWANG Hong-jiangjIANgZuO Qi-gao
(1Department of Vertebrate Paleontology,Natural History Museum of Los Angeles CountyLos Angeles, California 90007, USA * Corresponding author: xwang@nhm.org)
(2Key Laboratory of Vertebrate Evolution and Human Origins of Chinese Academy of Sciences,Institute of Vertebrate Paleontology and Paleoanthropology,Chinese Academy of SciencesBeijing 100044, China)
(3Administration Station of Cultural Relics of Xilinguole LeagueXilinhaote 026000, Nei Mongol, China)
(4University of Chinese Academy of SciencesBeijing 100049)
New record of a haplocyonine amphicyonid in early Miocene of Nei Mongol fi lls a long-suspected geographic hiatus
WANg xiao-ming1,2*WANG Hong-jiang3jIANgZuO Qi-gao2,4
(1Department of Vertebrate Paleontology,Natural History Museum of Los Angeles CountyLos Angeles, California 90007, USA * Corresponding author: xwang@nhm.org)
(2Key Laboratory of Vertebrate Evolution and Human Origins of Chinese Academy of Sciences,Institute of Vertebrate Paleontology and Paleoanthropology,Chinese Academy of SciencesBeijing 100044, China)
(3Administration Station of Cultural Relics of Xilinguole LeagueXilinhaote 026000, Nei Mongol, China)
(4University of Chinese Academy of SciencesBeijing 100049)
We place on the record a newly discovered amphicyonid (beardogs) upper molar from the Early Miocene Lower Red Mudstone Member of Aoerban Formation in central Nei Mongol. This molar is highly diagnostic of European haplocyonine or North American temnocyonine, two subfamilies of beardogs that have long been known in those continents but notably absent in Asia. The new molar is strikingly similar toHaplocyonoides mordaxandTemnocyon percussorwith its dumbbell-shaped M1 outline, reduced parastyle, isolated protocone by a surrounding cingulum, and extreme reduction of pre- and postprotocristae. Given the limited material at hand, we tentatively refer the new Chinese fossil to the EuropeanHaplocyonoidescf.H. mordaxbecause of their similar size and age relationship. If this identi fi cation is correct, our new record thus fills a large gap in the geographic distribution of the haplocyonines and represents an excursion of this rare subfamily from Europe.
Aoerban, Nei Mongol; Early Miocene; Amphicyonidae; zoogeography
1 Introduction
Straddling between Europe to the west and North America to the east, Asia has long been suspected to be either a source of certain Cenozoic mammals that, thus far, are only found in Europe and/or North America, or at the very least, must have been a “jumping board” to and from the continents of destination as they disperse. When taxa are found in both Europe and North America, their absence in Asia thus fuels speculation of a Holarctic continuity,and the expectation that filling in the Asian gap may be a matter of time. Temnocyonine and haplocyonine amphicyonids are such an example of a long-suspected geographic hiatus waiting to be fi lled.
Temnocyoninae and Haplocyoninae are peculiar lineages of amphicyonids, or beardogs, with hypercarnivorous dentitions and in some species, digitigrade posture (Hunt, 2011). Members of these groups typically have a very trenchant lower molar battery that align their main cusps in a single row to facilitate shearing function and associated narrow, highcrowned premolar series (e.g., Bonis, 1973; Hunt, 2011; Peigné and Heizmann, 2003). Asian records of amphicyonids have generally lagged behind those of Europe and North America, partly because of a shorter history of fi eld explorations associated with a late start of scienti fi c enterprise in almost all Asian countries. Occasionally, fi eld records in Asia do catch up, and we have the satisfaction of reporting a new fossil record of the haplocyonines from the Early Miocene of Nei Mongol (Inner Mongolia) that con fi rms, once again, Asia has much to offer in our understanding of Holarctic zoogeography.
Abbreviations ACM, Amherst College Museum of Natural History, Amherst, Massachusetts; AMNH, Division of Paleontology, American Museum of Natural History, New York; F:AM, Frick Collection, American Museum of Natural History, New York; IVPP, Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing; IM, IVPP fossil localities from Nei Mongol; LACM, Natural History Museum of Los Angeles County, Los Angeles; NM, Northwest Museum, Portland State University, Portland, Oregon; SMF, Senckenberg Forschungsinstitut und Naturmuseum, Frankfurt; TRO, Timberlane Research Organization, Lake Wales, Florida; UCMP, Museum of Paleontology, University of California, Berkeley; UNSM, Vertebrate Paleontology, University of Nebraska State Museum, Lincoln; USNM, Department of Paleobiology, Smithsonian Institution, Washington, D.C.; YPM-PU, Princeton Collection, Peabody Museum, Yale University, New Haven.
2 Systematic paleontology
Class Mammalia Linnaeus, 1758
Order Carnivora Bowdich, 1821
Family Amphicyonidae Haeckel, 1866
Subfamily Haplocyoninae Bonis, 1966
GenusHaplocyonoidesHürzeler, 1940
Haplocyonoidescf.H. mordaxHürzeler, 1940
Referred specimen IVPP V 22639, an isolated left M1. Collected by X. Wang on August 8, 2015.
Locality, stratigraphy, and age IVPP IM1530 locality (XWIM04), N43°20'34'', E113°54'30'', elevation 1050 m, Sonid Zuoqi, Xilinhot League, central Nei Mongol (Fig. 1).Locality IM1530 is stratigraphically straddling the boundary of the Lower Red Mudstone and Middle Green Mudstone members of Aoerban Formation (Fig. 2). Locally the two members transition into each other, possibly due to diagenetic effects of the green coloration. IVPPV 22639 thus belongs to the upper-most Lower Aoerban Fauna (Wang et al., 2009). Based on biochronology of small mammals, Qiu et al. (2013a) estimated that the upper part of the Lower Aoerban Fauna was approximately 19 Ma in the late Xiejian or possibly early Shanwangian Chinese land mammal ages (see Qiu et al., 2013b).
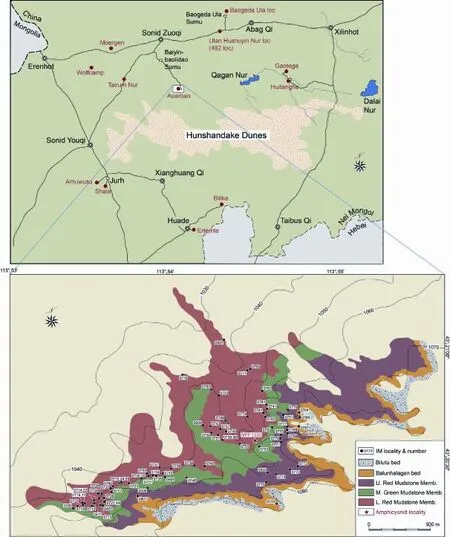
Fig. 1 Location of Aoerban area in central Nei Mongol (upper) and IVPP IM1530 locality (red star) and updated geologic map of Aoerban strata (lower) modi fi ed from Wang et al. (2009). Note that distribution of the Bilutu bed (light grey) differs substantially from that of Wang et al. (2009: fi g. 1).

Fig. 2 Photograph of the IVPP IM1530 locality (marked by geologic hammer) at a transitional zone between the Lower Red Mudstone and Middle Green Mudstone members of Aoerban Formation Stratigraphic scheme follows that of Wang et al. (2009: fi g. 3). Photo looking south
Description With the exception of the missing roots, IVPP V 22639 is almost perfectly preserved, missing only the anterior rim of the enamels at the waist. Fresh breakage at the base of the roots indicates that the roots were broken after re-emergence from sediment encasement during the erosion. V 22639 shows signs of only minor wear at the apex of the metacone and thus represents that of a young adult.
V 22639 is almost twice as transversely wide as anteroposteriorly long (see measurements in Table 1). Overall it is dumbbell-shaped with a distinctly constricted waist at the junction of the protocone on the lingual side and paracone-metacone on the labial side. In occlusal view, paracone and metacone are about the same size, although the paracone is taller; this is especially true when viewed from the lingual side, but less so from the labial side. A parastyle is poorly developed, indicated by a slightly thickened and higher cingulum surrounding the parastyle. A distinct notch separates the paracone and metacone. Both paracone and metacone have an anterior and posterior ridge, which are aligned in a single line, and are completely encircled by a thin cingulum.
The protocone has a rounded base and located slightly anterior to the paraconemetacone notch. The low-crowned protocone is about half as tall as the metacone and about 1/3 of paracone. An extremely faint preprotocrista is present on its anterolabial face and ispointed to the narrowest point (anterior waist) of the tooth. A postprotocrista is slightly more distinct than the preprotocrista. It is anteroposteriorly oriented and ends posteriorly at the posterior cingulum. Both paraconule (protoconule) and metaconule are absent and this gives the protocone an isolated appearance. The protocone is nearly completely encircled by a cingulum of various thickness. Lingually this cingulum forms an almost perfect half circle and has similar crown-heights throughout the protocone. The cingulum, however, thickens toward the posterolingual aspect of the protocone, more than twice as thick as its anterior segment, and along the medial and
posterior sides of the protocone the cingulum has some minor wrinkles on its crest.
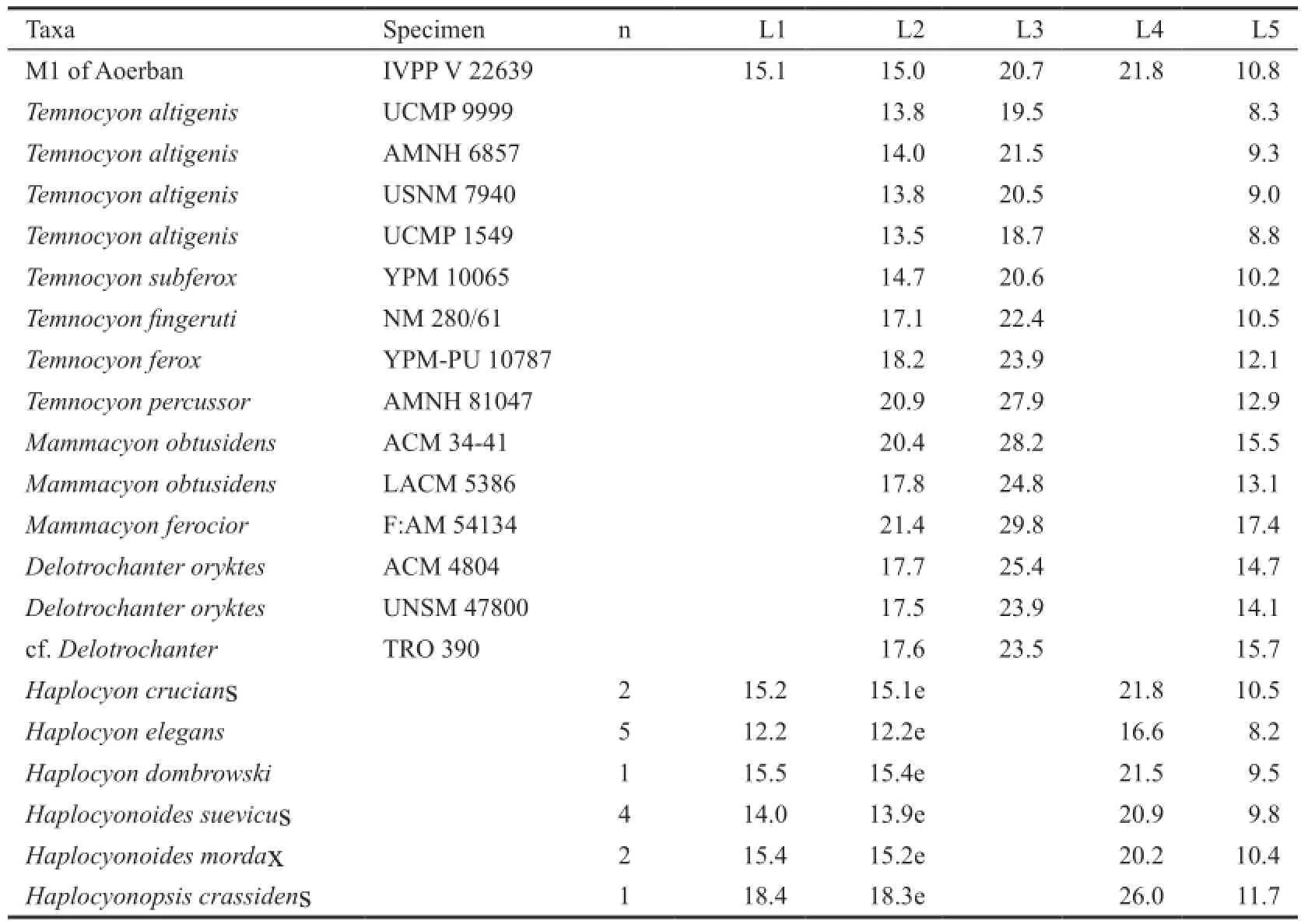
Table 1 Measurements of upper fi rst molars of Temnocyoninae and Haplocyoninae (mm)
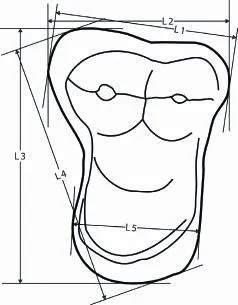
Fig. 3 De fi nitions of measurements for IVPP V 22639 L1. labial length; L2. anteroposterior length; L3. mediolateral width; L4. maximum transverse width; L5. length at protocone area
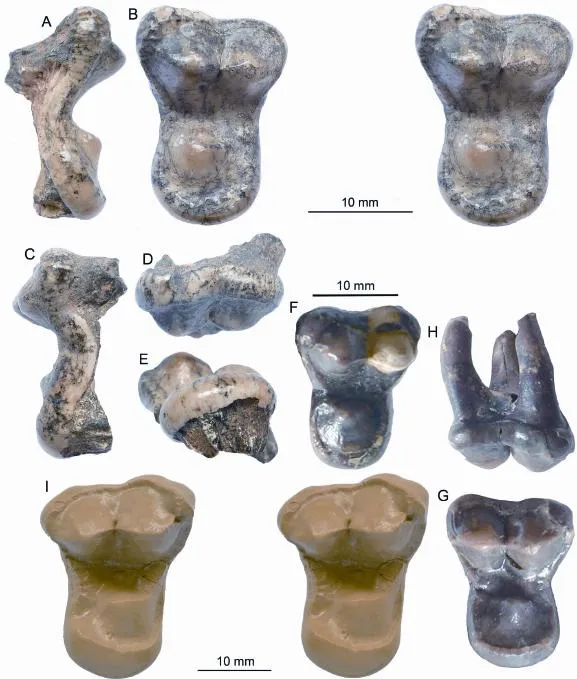
Fig. 4Haplocyonoidescf.H. mordaxHürzeler, 1940 (A-E), IVPP V 22639, left M1 from Aoerban, Nei Mongol,Haplocyonoides mordaxHürzeler, 1940 (F-H), andTemnocyon percussorCook, 1909 (I), AMNH 81047, right M1 from Cook Quarry, Sioux County, Nebraska A. anterior view; B. stereo occlusal view; C. posterior view; D. labial view; E. lingual view; F. occlusal view of SMF-M1653e, left M1, paralectotype (sensu Peigné and Heizmann, 2003) from Hessler, Mainz Basin, Germany; G. occlusal and H. labial views of SMF-M6000, right M1, referred specimen from Budenheim, Rhineland-Palatinate, Germany (photograph courtesy of Stéphane Peigné of Muséum national d’Histoire naturelle, Paris); I. stereo occlusal view (photograph of a cast). Top scale bar is for A-E, middle one is for F-H, and lower one is for I
In anterior and posterior views, V 22639 has a distinctly concave appearance (Fig. 4A,C) due to a deep trigon basin that occludes with a high-crowned m1 hypoconid. The highcrowned paracone-metacone crest shears with the labial face of the m1 hypoconid.
3 Comparisons
Despite the meager material from Aoerban, there is no doubt that V 22639 belongs to Amphicyonidae, although a dumbbell-shaped M1 is found in other similar-sized carnivorans, such asEnhydrocyonandParaenhydrocyonof hesperocyonine canids (Wang, 1994),Megalictisof basal musteloids (Matthew, 1907), andEomellivoraof mellivorine mustelids (e.g., Valenciano et al., 2015). However, in all of the latter three cases, a prominently developed M1 parastyle dominates the labial half of the tooth with a much larger paracone and very reduced metacone, in sharp contrast to a more evenly developed paracone and metacone (with a more quadrate outline for the labial half of the tooth) in most amphicyonids.
The Aoerban tooth is easily distinguished from all known amphicyonids in Asia. Excluding small, primitive forms in the Late Eocene through Early Miocene, such asGuangxicyon sinoamericanusfrom the Late Eocene of Guangxi Province (Zhai et al., 2003), unnamed species from Late Eocene of Mongolia (Egi et al., 2009),Cynodictis elegansfrom middle Oligocene of Nei Mongol (Huang, 1982), and the questionableIctiocyoncf.I. socialisfrom Early Miocene of Gansu (Wang et al., 2005), almost all Asian Miocene forms are large amphicyonine amphicyonids. Well known examples include the Early MioceneAmphicyon confucianusandYsengriniasp. from Shanwang, Shandong Province (Qiu and Qiu, 2013; Qiu et al., 1986; Young, 1937), Middle MioceneYsengriniasp. of Japan (Kohno, 1997; Kohno et al., 1997), Middle MioceneA. tairumensisof Nei Mongol (Colbert, 1939) and Gansu (Deng et al., 2013), Middle MioceneA. ulungurensisof Xinjiang (Qi, 1989), and Late MioceneA. palaeoindicusandVishnucyoncf.V. chinjiensisfrom Yunnan Province (Qi, 2006), although caution must be exercised in some of these generic assignments because many Asian taxa of large size, often based on fragmentary materials, tend to be placed inAmphicyonas tentative assignments (Hunt, 2003). From Southeast and South Asia, several taxa are assignable to amphicyonines, such as Middle MioceneMaemohcyonpotisatiof Thailand (Peigné et al., 2006), Late MioceneAmphicyonof Myanmar (Egi et al., 2010; Sein and Thein, 2011),A. sindiensis,A. palaeoindicus,A. pithecophilus,Arctamphicyon lydekkeri, andVishnucyon chinjiensisfrom Early to Late Miocene Siwalik strata of India and Pakistan (Colbert, 1935; Lydekker, 1884; Pilgrim, 1932). M1 inV. chinjiensishas a markedly narrowed lingual half with a constricted appearance, but its protocone retains distinct pre- and postprotocristae (Pilgrim, 1932:plate II, fi g. 9). When present, the M1s in these Asian forms have triangular outlines with distinct pre- and postprotocristae typical of amphicyonines, and are easily distinguishable from the dumbbell-shaped Aoerban M1. Even without an upper molar, these taxa have the reduced, low-crowned lower premolars or basined lower molars in contrast to the high-crowned premolars and trenchant molars in haplocyonines and temnocyonines.
Aktaucyon brevifacialisfrom the Early Miocene of Kazakhstan shows a hint of waistconstriction in M1 (Kordikova et al., 2000). However, its pre- and postprotocristae are still distinct and it has a large M2 relative to M1, features that suggest that its M1 constriction may be a result of convergence. The Middle MioceneGobicyon macrognathusfrom Nei Mongol (Colbert, 1939; Zhai, 1964), and possibly also from Xinjiang (Wang et al., 1998) and Gansu (Deng et al., 2013), may be an aberrant form of haplocyonines because of its high-crowned, piercing premolars, although its fragmentary m1 talonid appears to be not quite as trenchant as its European relatives (Hunt, 1998:208). Unpublished upper M1s from Hezheng area, Gansu Province, also exhibit a dumbbell shape (Qiu Zhanxiang pers. comm.). A Serbian (formerly Yugoslavian) partial mandible previously referred toG. macrognathusby Pavlovic and Thenius (1959) has since become the basis of a new speciesHaplocyonoides serbiae(Ginsburg, 1999a), although its diagnostic characters are still in dispute (Peigné and Heizmann, 2003).
The distinctly constricted waist with a dumbbell outline in V 22639 is the most salient feature of this specimen, seen only in European haplocyonines and North American temnocyonines. Haplocyoninae is a lineage of hypercarnivorous amphicyonids from Late Oligocene to middle Miocene of Europe, commonly including genera such asHaplocyon,Haplocyonoides, andHaplocyonopsis(Ginsburg, 1999a; Peigné and Heizmann, 2003). Dentally, it is characterized by high-crowned premolars and highly trenchant lower molars that may lose metaconids and entoconids (Bonis, 1966, 1973; Helbing, 1928; Peigné and Heizmann, 2003; Rothausen, 1988). Most haplocyonine species with a known M1 (includingHaplocyon crucians,Haplocyon elegans,Haplocyon dombrowskii,Haplocyonopsis crassidens,Haplocyonoides mordax,Haplocyonoides suevicus) tend to be somewhat more primitive than V 22639 in their stronger parastyle, larger paracone relative to metacone with more acute angle of labial border of M1 with that of P4, M1 protocone with at least a remnant pre- and postprotocristae, and not very constricted waist area.
Advanced haplocyonines may have a dumbbell-shaped upper M1s, such as inHaplocyonoides mordax(Hürzeler, 1940). However, published figures of the only M1 ofH. mordax(SMF-M1653e; Hürzeler, 1940:figs. 3-5) from the type locality in Hessler, Mainz Basin show several differences from that of V 22639: a relatively larger protocone, narrow cingulum surrounding the protocone, and a stronger preprotocrista with a distinct paraconule at the end (see Fig. 4F). Peigné and Heizmann (2003), however, referred a second M1 (SMF-M6000) from Budenheim, Rhineland-Palatinate, Germany, which has never been fi gured thus far. S. Peigné has graciously provided a photograph of this specimen to us that permits a much needed sense of variation (Fig. 4G, H). This referred specimen is strikingly similar to V 22639 in its much reduced protocone as a result of a widened surrounding cingulum, greatly reduced pre- and postprotocristae, and a nearly complete loss of a paraconule. As a result, lingual half of SMF-M6000 assumes a distinct appearance of an isolated protocone, not only because of its smaller protocone footprint relative to the broad surrounding cingulum but also its isolation from the trigon basin due to the reduction of preand postprotocristae. Details on the labial half of this tooth are also remarkably consistent withthat of V 22639. The labial side of SMF-M6000 is almost symmetrical with nearly identical size of paracone and metacone to the point that it is dif fi cult to ascertain which side this tooth belongs without associated P4 or M2. Nevertheless judging by the slightly higher-crowned paracone and parastyle (Fig. 4H), we interpret this tooth to be a right M1, which, if correct, would be slightly more derived than V 22639 because of the general morphocline among European haplocyonines toward decreased parastyle and symmetrical paracone and metacone. Minor differences between the European and Chinese forms include a more anteriorly located protocone, a slightly more bulging parastyle, and a less distinct (not very ridge-like) lingual cingulum in V 22639.
Haplocyonoides mordaxwas initially established from Hessler near Wiesbaden in Mainz Basin, Germany in the Early Miocene (MN2) based on isolated upper and lower teeth (Hürzeler, 1940) that may or may not belong to a single individual (Rothausen, 1988). Bonis (1973:pl. 4, fi g. 5) referred an additional right maxillary fragment with P4-M1 from Laugnac (MN2, Lot-et-Garonne) as ?H. mordax, although he noted that its M1 lacks a median constriction. More recently Ginsburg (1999b:119) also listedH.cf.H. mordaxfrom“faluns of Anjou (MN3 or 5)” in addition to recognizing a new species,H. serbiae(formerlyGobicyon macrognathusby Pavlovic and Thenius, 1959), plus the previously describedH. ponticus(Kuss, 1960). While describing a new German species,H. suevicus, Peigné and Heizmann (2003:appendix) provided a complete list of hypodigm forH. mordax. In addition to materials from the type locality and Laugnac mentioned above, they included one specimen from Weisenau (MN1) and three specimens from Budenheim (MN2), both from Mainz Basin in Rhineland-Palatinate, Germany. Furthermore, they (Peigné and Heizmann, 2003:68) listed several sites from the MN3 of France and M2 of Spain, but preferred to treatH. mordax huerzeleri(Rothausen, 1988) asnomen dubiumas well as casting doubt over generic assignments ofH. serbiae andH. ponticus.
V 22639 is also quite comparable to some North American temnocyonines, which share considerable similarities with European haplocyonines, interpreted to be convergences by Hunt (2011). In several respects temnocyonines have become even more hypercarnivorous than haplocyonines with short and robust crushing P4 in advanced forms, M1 protocone isolated by a fl at platform of surrounding cingulum, substantial reduction of M2 and loss of M3, and digitigrade posture (Hunt, 2011). The M1 morphology is particularly diagnostic for the temnocyonines and clear trends can be observed to help delineate lineages. The smallest and most basal species,Temnocyon altigenis, has an enlarged parastyle, a protocone at the beginning stage of isolation but still with a preprotocrista connecting to a small paraconule, and a lingual cingulum slightly widened. Successively larger and more derived species, such asT. subferoxandT. ferox, have a more isolated protocone, lost preprotocrista (inT. subferoxbut not inT. ferox) and paraconule, and more expanded lingual cingulum.T. fingeruti, apparently a side branch from the main chronospecies series (T. altigenis-subferox-ferox) (Hunt, 2011:fig. 70), does not quite follow this same trend, such as its retaining a strongpre- and postprotocristae and a lack of middle constriction. Large, bone-crushing genera,MammacyonandDelotrochanter, further elaborate this theme by widening the cingulum surrounding protocone, among other specializations.
In light of above trends within the mainTemnocyonchronospecies series (Hunt, 2011: fi g. 70), V 22639 falls somewhere close toT. subferoxandT. feroxboth in metric plot (Fig. 5) and some qualitative characters. The Chinese form has extremely reduced pre- and postprotocristae, not quite the complete loss inT. subferox(Hunt, 2011: fi g. 10A) but certainly exceeded that condition inT. ferox(Hunt, 2011: fi g. 12B). However, neither of the two North American species has reduced their parastyles quite to the extent as in V 22639. The degree of protocone area enlargement in V 22639 is also somewhat more pronounced than the above two American species. We also note that some characters are not strictly in a linear sequence, such as the precocious reduction of pre- and postprotocristae inT. subferox, although current knowledge about intraspeci fi c variations is too limited to permit evaluation of morphological outliers.
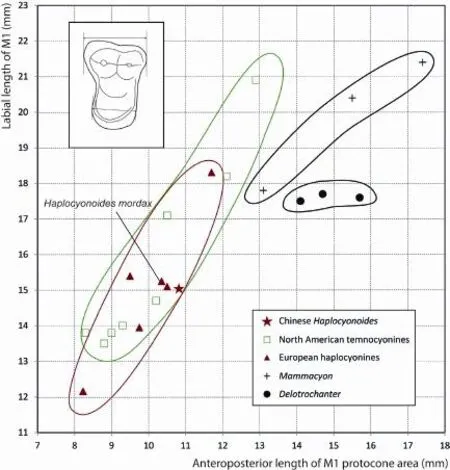
Fig. 5 Two dimensional plots of M1 anteroposterior length vs. M1 protocone length (see Fig. 3 for de fi nitions) of Haplocyoninae and Temnocyoninae, as originally devised by Hunt (2011: fi g. 46) to illustrate relative enlargement of the protocone area (Data from Table 1)
A poorly known large species,Temnocyon percussorCook, 1909, is also intriguingly close to V 22639. An isolated M1 (AMNH 81047) from the type locality (Cook Quarry,Anderson Ranch Formation, Sioux County, Nebraska, late Arikareean) was referred to this species (Fig. 4I). As described by Hunt (2011), this tooth is remarkably similar to V 22639. Its large size aside, AMNH 81047 has almost identical morphology of V 22639 in its protocone area enlargement: widening of lingual cingulum, reduction of pre- and postprotocristae, and reduction of parastyle. The only subtle difference is a slightly more convex labial border, compared to a very faint concave border in V 22639.
Although we cannot completely rule out the new Aoerban material being related to the North American temnocyonine, the balance of evidence, as shown in a single M1, seems to tip toward something close toHaplocyonoides mordax. In favor of such an assignment is the relatively closer age relationship of the Chinese and European forms (see Zoogeographic remarks below). If such a taxonomic assessment is correct, future discoveries of more complete materials of this Nei Mongol species should bear additional haplocyonine characters.
4 Zoogeographic remarks
In his study of Aquitanian mammals, Bonis (1973) commented the striking morphological similarities between European haplocyonines and North American temnocyonines, as was also noted earlier by Viret (1929). Bonis regarded these similarities as parallel developments by groups that are confined to their respective continents, a conclusion also maintained by Hunt (1996, 1998, 2011), who further explicitly derived his Temnocyoninae from within an early stock of Haplocyoninae (Hunt, 1998:fig. 11.3). Additionally, European haplocyonines and North American temnocyonines seem to have their independent evolutionary trajectory and chronological sequences, at least in the case of more derived temnocyonines (Fig. 5). If that is the case, it implies a single immigration event during the Early Oligocene that brought a primitive haplocyonine to North America to give rise to the temnocyonines, as has been postulated by Hunt (1998). However,Haplocyonoides mordaxis much too late and too derived to be ancestral to temnocyonine.
If Peigné and Heizmann’s (2003) concept ofHaplocyonoides mordaxis to be followed, the age range forH. mordaxextends to European Neogene mammal zones 1-3. If we take this at its face value, i.e., a species range of as long as ~5 million years (23-18 Ma), the later part of the European records would be equivalent in age to the new Nei Mongol record. However, based on actual M1s from MN2 of Germany that we can compare, the Chinese form is likely an eastward expansion of this lineage. If so, the Aoerban form would be the fi rst occurrence of this lineage in Asia, possibly followed by laterGobicyonin the Middle Miocene.
Acknowledgments We have profited greatly from discussions with Robert M. Hunt, Jr., who has been unstinting in sharing his vast knowledge of European and North American amphicyonids. We also greatly appreciate exchanges with Stéphane Peigné, who generously provided unpublished photos that are critical in the formulation of ideas in this paper.Discussion with Louis de Bonis is also helpful in fossil identi fi cation. We thank Naoki Kohno for providing relevant reprints and a cast, and Naoko Egi for furnishing photos of Myanmar amphicyonid materials. We appreciate careful reviews by Qiu Zhanxiang and Zhang Zhaoqun; their comments and suggestions greatly improved this paper. We gratefully acknowledge the hard works by our 2015 fi eld crew: Deng Tao, Yuri Kimura, Li Yikun, Sun Danhui, as well as our drivers Feng Wenqing and Gao Wei. Funding for fi eld works during the past 20 years was provided by: Chinese National Natural Science Foundation (No. 41430102), Strategic Priority Research Program of the Chinese Academy of Science (XDB03020104), Chinese Academy of Science Outstanding Overseas Scholar Fund (KL205208), National Science Foundation (US) (EAR-0446699, 0444073, 0958704, 1227212 to XW), and National Geographic Society (NGS 5527-95 to XW).
Bonis L d, 1966. Sur l’évolution du genreHaplocyonSchlosser (Carnivora). Bull Soc géol Fr, 8: 114-117
Bonis L d, 1973. Contribution à l’Étude des Mammifères de l’Aquitanien de l’Agenais, rongeurs-carnivoresperissodactyles. Mém Mus Natl Hist Nat, 28: 1-192
Bowdich T E, 1821. An analysis of the natural classi fi cations of Mammalia, for the use of students and travellers. Paris: J. Smith. 1-115
Colbert E H, 1935. Siwalik mammals in the American Museum of Natural History. Trans Am Philos Soc, Philadelphia, New Ser, 26: 1-401
Colbert E H, 1939. Carnivora of the Tung Gur Formation of Mongolia. Bull Am Mus Nat Hist, 76: 47-81
Cook H J, 1909. Some new Carnivora from the Lower Miocene beds of western Nebraska. Bull Nebraska Geol Surv, 3(9): 261-272
Deng T, Qiu Z x, Wang B Y et al., 2013. Late Cenozoic biostratigraphy of the Linxia Basin, northwestern China. In: Wang X, Flynn L J, Fortelius M eds. Fossil Mammals of Asia: Neogene Biostratigraphy and Chronology. New York: Columbia University Press. 243-273
Egi N, Tsubamoto T, Tsogtbaatar K, 2009. New amphicyonid (Mammalia: Carnivora) from the upper Eocene Ergilin Dzo Formation, Mongolia. Paleont Res, 13(3): 245-249
Egi N, Sein C, Maung-Thein Z-M et al., 2010. A new amphicyonid (Mammalia: Carnivora) from the lower Irrawaddy sediments (Myanmar) with comments onAmphicyonspecies from the Miocene of Asia. J Vert Paleont, 30(Suppl 3): 84A
Ginsburg L, 1999a. Order Carnivora. In: Rössner G E, Heissig K eds. The Miocene Land Mammals of Europe. München: Verlag Dr. Friedrich Pfeil. 109-148
Ginsburg L, 1999b. Order Creodonta. In: Rössner G E, Heissig K eds. The Miocene Land Mammals of Europe. München: Verlag Dr. Friedrich Pfeil. 105-108
Haeckel E, 1866. Generelle Morphologie der Organismen. Allgemeine Grundzüge der organischen Formen-Wissenschaft, mechanisch begründet durch die von Charles Darwin reformirte Deszendenz-Theorie. Band I: Allgemeine Anatomie der Organismen. Berlin: Georg Reimer. 1-462
Helbing H, 1928. Carnivoren des oberen Stampien. Abh Schweiz Palaeont Ges, 47: 1-82
Huang X S, 1982. Preliminary observation on the Oligocene stratigraphic section and on the fauna of the Wulantata’er area, Alxa Left Banner, Inner Mongolian Autonomous Region. Vert PalAsiat, 20(4): 337-345
Hunt R M Jr., 1996. Amphicyonidae. In: Prothero D R, Emry R J eds. The Terrestrial Eocene-Oligocene Transition in North America. Cambridge: Cambridge University Press. 476-485
Hunt R M Jr., 1998. Amphicyonidae. In: Janis C M, Scott K M, Jacobs L L eds. Evolution of Tertiary Mammals of North America, Volume 1: Terrestrial Carnivores, Ungulates, and Ungulatelike Mammals. Cambridge: Cambridge University Press. 196-227
Hunt R M Jr., 2003. Intercontinental migration of large mammalian carnivores: earliest occurrence of the Old World beardogAmphicyon(Carnivora, Amphicyonidae) in North America. In: Flynn L J ed. Vertebrate Fossils and Their Context: Contributions in Honor of Richard H. Tedford. Bull Am Mus Nat Hist, 279: 77-115
Hunt R M Jr., 2011. Evolution of large carnivores during the Mid-Cenozoic of North America, the temnocyonine radiation (Mammalia, Amphicyonidae). Bull Am Mus Nat Hist, 358: 1-153
Hürzeler J, 1940.Haplocyonoidesnov. gen., ein aberranter Canide aus dem Aquitanien des Hesslers (Mainzer Becken). Eclogae geol Helv, 33(2): 224-229
Kohno N, 1997. The first record of an amphicyonid (Mammalia: Carnivora) from Japan, and its implication for amphicyonid paleobiogeography. Paleont Res, 1(4): 311-315
Kohno N, Yamaoka T, Sugihara M, 1997. A Miocene amphicyonid (Mammalia: Carnivora) from Shobara City, Hiroshima Prefecture, southwestern Japan. J Hiba Soc Nat Hist, 182: 1-9
Kordikova E G, Heizmann E P J, Mavrin A V, 2000. Early Miocene Carnivora of Aktau Mountains, south eastern Kazakhstan. Paläont Z, 74(1/2): 195-204
Kuss S E, 1960.Haplocyonoides ponticusn. sp., ein neuer aberranter Canide aus dem Unterpliocän von Melchingen. Ber Naturf Ges Freiburg, 50(2): 247-250
Linnaeus C, 1758. Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Vol. 1: Regnum animale. Editio decima, 1758 (12th ed of Linaeus 1758). Stockholm: Societatis Zoologicae Germanicae. 1-824
Lydekker R, 1884. Indian Tertiary and post-Tertiary vertebrata. Vol. II, Part 6, Siwalik and Narbada Carnivora. Mem Geol Surv India Palaeont Indica, 2: 178-355
Matthew W D, 1907. A Lower Miocene fauna from South Dakota. Bull Am Mus Nat Hist, 23: 169-219
Pavlovic M, Thenius E, 1959.Gobicyon macrognathus(Canidae, Mammalia) aus dem Miozän Jugoslawiens. Anz Akad Wiss Wien, 11: 214-222
Peigné S, Heizmann E P J, 2003. The Amphicyonidae (Mammalia: Carnivora) from Ulm-Westtangente (MN2, Early Miocene), Baden-Württemberg, Germany - systematics and ecomorphology. Stuttg Beitr Naturkd Ser B, 343: 1-133
Peigné S, Chaimanee Y, Yamee C et al., 2006. A new amphicyonid (Mammalia, Carnivora, Amphicyonidae) from the late middle Miocene of northern Thailand and a review of the amphicyonine record in Asia. J Asian Earth Sci, 26: 519-532
Pilgrim G E, 1932. The fossil Carnivora of India. Mem Geol Surv India Palaeont Indica New Ser, 18: 1-232
Qi g Q, 2006. Order Carnivora. In: Qi G Q, Dong W eds.Lufengpithecus hudienensisSite. Beijing: Science Press. 148-177
Qi T, 1989. Miocene carnivores from Altai region, Xinjiang. Vert PalAsiat, 27(2): 133-139
Qiu Z D, Qiu Z x, 2013. Early Miocene Xiejiahe and Sihong fossil localities and their faunas, eastern China. In: Wang X, Flynn L J, Fortelius M eds. Fossil Mammals of Asia: Neogene Biostratigraphy and Chronology. New York: Columbia University Press. 142-154
Qiu Z D, Wang X, Li Q, 2013a. Neogene faunal succession and biochronology of central Nei Mongol (Inner Mongolia). In: Wang X, Flynn L J, Fortelius M eds. Fossil Mammals of Asia: Neogene Biostratigraphy and Chronology. New York: Columbia University Press. 155-186
Qiu Z x, Qiu Z D, Deng T et al., 2013b. Neogene land mammal stages/ages of China - toward the goal to establish an Asian land mammal stage/age scheme. In: Wang X, Flynn L J, Fortelius M eds. Fossil Mammals of Asia: Neogene Biostratigraphy and Chronology. New York: Columbia University Press. 29-90
Qiu Z x, Yan D F, Jia H et al., 1986. The large-sized ursid fossils from Shanwang, Shandong. Vert PalAsiat, 24(3): 182-194
Rothausen K, 1988. Carnivoren im Kalktertiär (Oberoligozän-Untermiozän) des Mainzer Beckens (1. Amphicyonidae). Geol Jahrb, 110: 241-260
Sein C, Thein T, 2011. A new amphicyonid (Mammalia, Carnivora) from the Ayeyarwady Formation of central Myanmar. Univ Res J, 4(5): 45-57
Valenciano A, Abella J, Sanisidro O et al., 2015. Complete description of the skull and mandible of the giant mustelidEomellivora piveteauiOzansoy, 1965 (Mammalia, Carnivora, Mustelidae), from Batallones (MN10), Late Miocene (Madrid, Spain). J Vert Paleont, e934570
Viret J, 1929. Les faunes de mammifères l’Oligocène supérieur de la Limagne Bourbonnaise. Ann Univ Lyon N S, 47: 1-328
Wang X, 1994. Phylogenetic systematics of the Hesperocyoninae (Carnivora: Canidae). Bull Am Mus Nat Hist, 221: 1-207
Wang X, Ye J, Meng J et al., 1998. Carnivora from middle Miocene of northern Junggar Basin, Xinjiang Autonomous Region, China. Vert PalAsiat, 36(3): 218-243
Wang X, Qiu Z x, Wang B Y, 2005. Hyaenodonts and carnivorans from the Early Oligocene to Early Miocene of Xianshuihe Formation, Lanzhou Basin, Gansu Province, China. Palaeont Electr, 8(1.6A): 1-14
Wang X, Qiu Z D, Li Q et al., 2009. A new Early to Late Miocene fossiliferous region in central Nei Mongol: lithostratigraphy and biostratigraphy in Aoerban strata. Vert PalAsiat, 47(2): 111-134
Young C C, 1937. On a Miocene mammalian fauna from Shantung. Bull Geol Soc China, 17: 209-245
Zhai R j, 1964.Leptarctusand other Carnivora from the Tung Gur Formation, Inner Mongolia. Vert PalAsiat, 8(1): 18-32
Zhai R j, Ciochon R L, Tong Y S et al., 2003. An aberrant amphicyonid mammal from the latest Eocene of the Bose Basin, Guangxi, China. Acta Palaeont Pol, 48(2): 293-300
内蒙古敖尔班地区早中新世犬熊类的新记录
王晓鸣1,2*王洪江3江左其杲2,4
(1 美国洛杉矶自然历史博物馆 洛杉矶 CA 90007 * 通讯作者)
(2 中国科学院脊椎动物演化与人类起源重点实验室,中国科学院古脊椎动物与古人类研究所 北京 100044)
(3 内蒙古锡林郭勒盟文物站 锡林浩特 026000)
(4 中国科学院大学 北京 100049)
犬熊科(Amphicyonidae)的4个亚科(Amphicyoninae, Daphoeninae, Haplocyoninae和Temnocyoninae)中,Haplocyoninae (简齿犬熊亚科)和Temnocyoninae (剪切犬熊亚科)是两类比较特殊的、高度食肉化的姊妹群。Haplocyoninae仅出现在欧洲的晚渐新世到早中新世,而Temnocyoninae则局限在北美的早渐新世到早中新世。目前亚洲虽还没有记录,但新近纪的亚洲无疑是欧洲与北美之间迁徙途中的必经之路。因此本文记录的新材料既是意外发现又是意料之中。2015年我们在内蒙古中部早中新世敖尔班组下红层中首次发现一枚犬熊类的左上第一臼齿。这颗臼齿虽然零星,但非常特征,属Haplocyoninae或Temnocyoninae无疑,值得记述。该牙齿由于中间的收缩及原尖周边齿带的变宽而具有明显的哑铃型轮廓。另外其原尖上的前、后脊也几乎消失,形成一种原尖缩小而且孤立的形态。上述特征与欧洲早中新世的Haplocyonoides mordax(咬合似简齿犬熊)似乎最接近,但由于材料稀少敖尔班犬熊类与北美的Temnocyon percussor更为接近的可能也不能完全排除。如果上述判断正确的话,新发现的Haplocyonoides则是目前发现的少数几例早中新世从欧洲迁移到亚洲的种类之一。这一支系在亚洲有可能延续到中中新世的Gobicyon(戈壁犬熊)。
内蒙古敖尔班,早中新世,犬熊亚科,地理分布
Q915.874
A
1000-3118(2016)01-0021-15
2015-10-26
Wang X M, Wang H J, Jiangzuo Q G, 2016. New record of a haplocyonine amphicyonid in Early Miocene of Nei Mongol fills a long-suspected geographic hiatus. Vertebrata PalAsiatica, 54(1): 21-35
国家自然科学基金重点项目(批准号:41430102)资助。
猜你喜欢
杂志排行

古脊椎动物学报(中英文)的其它文章
- Morphology and taxonomy of Gazella (Bovidae, Artiodactyla) from the Late Miocene Bahe Formation, Lantian, Shaanxi Province, China
- Restudy of the Late Oligocene dormice from northern Junggar Basin
- Female preference promotes asynchronous sex evolution in Elephantiformes
- A new hadrosauroid dinosaur from the Late Cretaceous of Tianzhen, Shanxi Province, China
- A new type of dinosaur eggs from Early Cretaceous of Gansu Province, China