甜菜碱生物学功能及其在改善猪肉质中的应用
2016-03-21孙进华宋文涛孙岳丞单安山东北农业大学动物营养研究所哈尔滨150030
张 婧,孙进华,宋文涛,孙岳丞,单安山(东北农业大学动物营养研究所,哈尔滨 150030)
甜菜碱生物学功能及其在改善猪肉质中的应用
张婧,孙进华,宋文涛,孙岳丞,单安山*
(东北农业大学动物营养研究所,哈尔滨150030)
摘要:甜菜碱对机体营养物质代谢具有重要作用,可有效改善不同生长阶段猪生产性能、胴体品质和肉品质,通过母子效应传递给子代,对子代进行营养调控。文章从化学结构、生物学功能、营养物质代谢及母体效应等方面,系统阐述不同添加量甜菜碱在改善不同生长阶段猪的机体性能、胴体品质及肉品质方面作用。
关键词:甜菜碱;生物学功能;机理;猪;肉质;母体效应
张婧,孙进华,宋文涛,等.甜菜碱生物学功能及其在改善猪肉质中的应用[J].东北农业大学学报, 2016, 47(1): 93-101.
Zhang Jing, Sun Jinhua, Song Wentao, et al. Biological functions of betaine and its application for pork quality in swine production[J]. Journal of Northeast Agricultural University, 2016, 47(1): 93-101. (in Chinese with English abstract)
甜菜碱(Betaine)是具有三甲基结构的多功能营养型饲料添加剂,既是猪胴体调节剂,又是高半胱氨酸抑制剂。因甜菜碱在动物机体内合成量较少,需从饲料中摄取,又称为准维生素。甜菜碱主要在动物肝、肾细胞线粒体内代谢,其吸收与氨基酸运输系统、甜菜碱-γ-氨基丁酸运输密切相关。甜菜碱不仅参与机体甲基代谢、营养物质(氨基酸、蛋白质、脂肪)代谢,还具有维持饲料中维生素效价、缓解机体应激、促进机体生长发育、改善肉品质等生物学作用,广泛应用于养猪生产。研究发现,妊娠期和哺乳期补充甜菜碱不仅对母体健康和繁殖性能有促进作用,通过母子效应还对子代产生积极影响[1]。本文从化学结构、生物学功能、营养物质代谢及母体效应等方面,系统阐述不同添加量甜菜碱在改善不同生长阶段猪的机体性能、胴体品质及肉品质方面作用。
1甜菜碱来源与结构
1.1甜菜碱来源
甜菜碱在动物、植物、海生无脊椎动物和微生物体内分布广泛。植物体内甜菜碱主要分布在根、茎、叶及种子等部位。常见饲料原料以麸皮中甜菜碱含量最高。动物体内甜菜碱主要是代谢的中间产物,在营养代谢中具有重要作用。目前,广泛用于动物饲料的甜菜碱主要包括甜菜碱、甜菜碱盐酸盐和甜菜碱预混剂三种形式。几种常见饲料原料中甜菜碱含量见表1[2-3]。

表1几种常见饲料原料中甜菜碱含量Table 1 Betaine content in several common feeds (mg·kg-1)
1.2甜菜碱理化性质
甜菜碱(Betaine)又名三甲基甘氨酸,化学名称N-N-N-三甲基甘氨酸内盐,分子式为C5H11NO2,化学结构式见图1,属于具备3个非稳态甲基的季胺型生物碱。甜菜碱在常温下多为白色棱状晶体,结构与氨基酸相似,具有两性解离性质。甜菜碱理化性质稳定,具有较高溶解性、热稳定性、保湿性和抗氧化性,能防止饲料生产、贮存过程中脂溶性维生素氧化,有效维持维生素稳定性和生物学效价。甜菜碱及甜菜碱盐酸盐对动物无致畸形、致癌、致突变作用,是公认的无毒、无污染饲料添加剂。甜菜碱盐酸盐结构式见图2,水溶性较差,可调节动物胃肠道pH,改善营养物质消化率。甜菜碱预混剂一般为粉剂,主要是在甜菜碱水溶液中加入饲料用赋形剂制作完成。

图1甜菜碱结构式Fig. 1 Structure of betaine

图2甜菜碱盐酸盐结构式Fig. 2 Structure of betaine hydrochloride
2甜菜碱生物学功能
2.1甜菜碱转甲基作用
甲基是机体蛋白质和脂肪代谢,DNA和RNA甲基化必需的功能基团。动物体内不能直接合成甲基,需从食物中摄取。饲料中常见甲基源包括胆碱、甜菜碱、蛋氨酸等。其中,胆碱可转化为甜菜碱,但此过程不可逆。甜菜碱含有3个活性甲基,是高效甲基供体,提供甲基能力高于胆碱和蛋氨酸。饲料中添加甜菜碱不仅可减少如蛋氨酸和胆碱等其他甲基供体需要量,还能降低饲料成本。甜菜碱转甲基作用一般发生在动物肝脏中,受甜菜碱-高半胱氨酸-甲基转移酶(Betaine- Homocyste⁃ine Methyl-Transferase,BHMT)和胱硫醚-β-合成酶(Cystathionine-β-Synthase,CβS)双重调控[4]。当甜菜碱添加量较低时,BHMT活性增加,CβS活性不变,为机体物质代谢提供充分甲基;当甜菜碱添加量较高时,BHMT和CβS活性均升高,保持机体甲基代谢平衡[5]。甜菜碱转甲基作用可影响机体氨基酸、蛋白质和脂肪代谢,间接影响猪胴体品质和肉品质。Deminice等研究发现,饲粮中添加甜菜碱对预防脂肪肝有积极作用[6]。
2.1.1甜菜碱与氨基酸、蛋白质代谢
甜菜碱与氨基酸、蛋白质代谢关系见图3[4,7-10]。
甜菜碱甲基供体和氨基酸,通过蛋氨酸循环对机体蛋白质和脂肪代谢起重要作用[11]。蛋氨酸循环(即高半胱氨酸蛋氨酸循环)是体内重要生化过程。高半胱氨酸(Homocysteine,Hcy)通过BHMT和亚甲基四氢叶酸还原酶(Methylenetetrahydrofolate reductase,MTHFR)转化为蛋氨酸(Methionine,Met)[12]。两条转化途径此消彼长,一条途径受到抑制,另一途径则加强。首先,在肝脏和肾脏细胞内,甜菜碱在BHMT作用下提供甲基进行转化。其次,在体细胞中,四氢叶酸(Tetrahydrofolate,THF)在MTHFR作用下将甲基传递给VB12。VB12在蛋氨酸合成酶(Methionine synthase,MS)作用下再将甲基传递给Hcy生成Met。
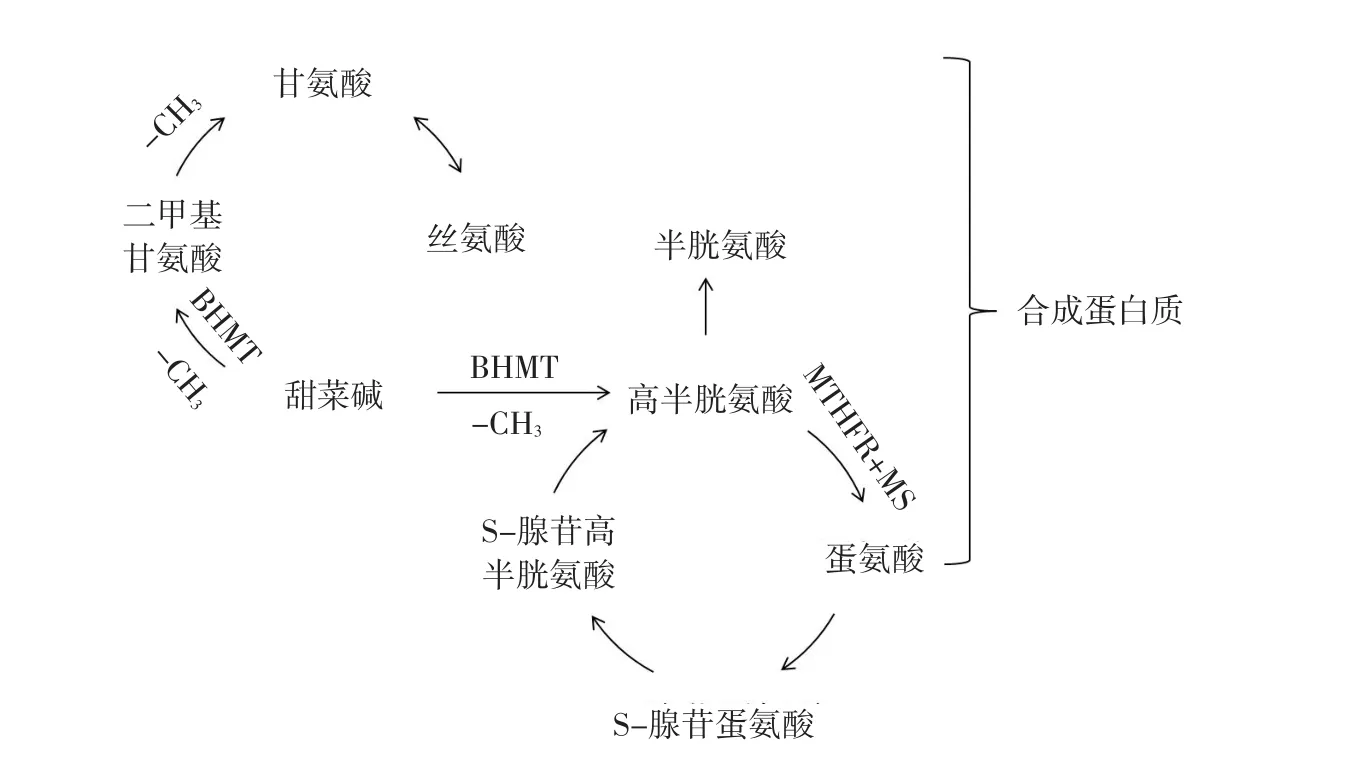
图3甜菜碱与氨基酸、蛋白质代谢Fig. 3 Betaine and amino acids, protein metabolism
甜菜碱与丝氨酸(Serine,Ser)、甘氨酸(Gly⁃cine,Gly)代谢有关。Ser与Gly既是机体合成甜菜碱前体物,又是甜菜碱代谢产物。甜菜碱通过转甲基和脱甲基2次作用后生成Ser和Gly,血液中游离Met、Ser、Gly含量升高[13]。
2.1.2甜菜碱与脂肪代谢
甜菜碱为体内肉碱和肌酸合成提供活性甲基,还可部分替代胆碱、蛋氨酸等参与体内脂肪代谢。当甜菜碱部分替代胆碱合成磷脂酰胆碱时,通过促进肝脏乳糜微粒合成,抑制肝脏脂肪蓄积,影响血液脂蛋白浓度[14]。当甜菜碱部分替代蛋氨酸合成肉碱时,促进脂肪酸β-氧化,加快脂肪分解,减少动物体脂含量,重新分配体脂[15]。另外,甜菜碱能提高胆碱活性,为合成低密度脂蛋白提供胆碱,促进低密度脂蛋白分泌。低密度脂蛋白可阻止脂肪在肝脏沉积,加快肝脏中脂肪动
员[16]。
甜菜碱还可通过增加脂肪组织中激素敏感脂酶(Hormone-Sensitive triglyceride lipase,HSL)活性和降低脂肪酸合成酶(Fatty acid syntheses,FAS)、苹果酸脱氢酶(Malate dehydrogenase,MDH)、脂蛋白脂酶(Lipoprotein lipase,LPL)活性,抑制脂肪组织合成[17-18]。此外,甜菜碱作为高半胱氨酸抑制剂,可降低机体高半胱氨酸含量,而高浓度高半胱氨酸可导致机体脂肪代谢紊乱。Cai等发现,饲粮中添加甜菜碱可显著降低生长肥育猪脂肪组织中FAS活性和FAS mRNA表达[19];在妊娠母猪饲粮中添加甜菜碱可下调新生仔猪肝脏中甘油三酯(Tri⁃glyceride,TG)含量及FAS mRNA表达量[19]。此外,甜菜碱代谢产物甘氨酸是胆酸盐轭合物主要组成成分,可间接促进脂肪吸收,提高脂肪消化吸收率[20]。Yang等添加甜菜碱可提高育肥猪生长性能,降低血液中总胆固醇浓度和肌肉中不饱和脂肪酸含量[21]。
2.2甜菜碱抗应激、调节渗透压作用
应激与下丘脑-垂体-肾上腺皮质轴(Hypotha⁃lamic pituitary adrenocortical,HPA)联系紧密。发生应激时,下丘脑神经元分泌促肾上腺皮质激素释放激素(Corticotrophin releasing Hormone,CRH),引起垂体-肾上腺皮质系统反应,促进肾上腺皮质激素(Adreno cortical hormones,ACTH)分泌,ACTH刺激糖皮质类固醇分泌,参与应激反应。甜菜碱抗应激作用与体内高半胱氨酸含量密切相关。甜菜碱通过降低体内高半胱氨酸含量,促进脑内γ-氨基丁酸(γ-aminobutyric acid,GABA)合成。GABA是哺乳动物中枢神经系统中重要的抑制性神经传导物质,参与神经系统活动,抑制与应激有关激素分泌,维持各中枢对机体各系统正常调节机能,具有提高生产性能、改善胴体品质、增强机体免疫力及缓解应激等多种功能及调节作用。GABA缺乏时,易产生焦虑、不安等负面情绪;GABA合成增加时,中枢抑制作用加强,机体抗应激能力增强。此外,甜菜碱两性离子特征在肾脏、肝脏、脑和肠道中发挥抗盐应激和高温应激作用,其中肠道是甜菜碱发挥渗透作用主要场所[22-23]。贺绍君等认为,饲粮中添加甜菜碱可改善热应激AA肉鸡的生产性能,提高十二指肠消化酶活性,维持盲肠微生物平衡,减轻热应激对肉鸡的危害[24]。
饲料中甲基供体浓度和肠道等器官的渗透压直接影响动物对甜菜碱需要量。若甜菜碱不能满足内源代谢,饲料中补充甜菜碱可提高特定营养物质消化率,改善动物健康和生产性能。同时,甜菜碱作为一种渗透压缓冲剂,应激时不仅可维持动物体渗透压平衡,防止细胞中离子浓度激变,还能有效增加Na+-K+泵效率,减少能量消耗,帮助维持正常渗透压平衡。当机体遇到缺水、长期腹泻等渗透压应激时,适量补充甜菜碱,可防止细胞中水分流出和盐分入侵,维持渗透压平衡,增强动物抗腹泻能力[25]。甜菜碱可增强猪肺动脉内皮细胞抗高渗环境的耐受力,防止细胞凋亡[26]。甜菜碱调节渗透压作用不仅可缓解离子载体类抗球虫药造成的肠道渗透压紊乱,还可促进肠道上皮细胞增殖,降低断奶仔猪粪便含水量和腹泻发生率,提高仔猪营养物质(如氨基酸)消化率[27]。
2.3甜菜碱诱食与促营养物质吸收作用
甜菜碱具有动物易感知的甜味和鲜味(尤其是仔猪),可诱导动物采食并影响猪胃肠道。一定范围内,仔猪回肠营养物质(如干物质、中性洗涤纤维、粗蛋白、大多数氨基酸等)消化率和仔猪大肠中细菌降解的粗蛋白质和氨基酸降解率,会随饲粮中甜菜碱添加量升高而升高[28-29]。另一方面,甜菜碱作为有机渗透保护剂,具有调节小肠上皮细胞渗透压,维持十二指肠上皮细胞水分平衡,维持肠壁细胞完整性,促进营养物质吸收等功能[13]。还可刺激肠道微生物活性,影响微生物发酵,提高产气量和氨基酸浓度,增加氨基酸消化率。但此作用仅限于甜菜碱纯品,其盐酸盐和预混剂对肠道微生物发酵无影响[26]。
3甜菜碱在改善猪生产性能和肉品质中的应用
甜菜碱作为饲料添加剂,在猪养殖业上应用广泛。添加甜菜碱可有效改善母猪繁殖性能;增加新生仔猪活重、断奶重、平均日增重和饲料转化率以及育肥猪的日增重和采食量;降低生长育肥猪的料重比及脂肪在猪胴体和内脏的沉积效率,提高屠宰率,改善胴体品质和肉品质。在生命的初期阶段,甜菜碱是重要甲基源,对胚胎和胎儿生长发育至关重要。母猪在妊娠期间补充甜菜碱,显著降低母猪肝脏中甘油三酯含量,降低子代肝脏中胆固醇含量,下调子代脂肪合成基因表达量,影响子代肝脏胆固醇代谢[1,4-5, 19, 28, 30-39]。甜菜碱对猪生产性能和肉品质影响见表2。
3.1甜菜碱与胴体品质
胴体品质包括胴体性状(屠宰率、膘厚、眼肌面积等)、胴体分离指标(瘦肉率、脂肪率等)及产肉性能。胴体品质不仅受基础日粮营养水平影响,还与品种、性别、生长阶段和饲养管理等有关。研究发现,甜菜碱具有改善生长育肥猪胴体品质作用,降低胴体脂肪分数,提高胴体瘦肉率、背最长肌面积以及里脊肉、肩颈肉、脊柱等重量[4, 35-37, 40]。因此,甜菜碱又称为“胴体调节剂”。甜菜碱作用效果与日粮营养水平、生长阶段、添加量及使用时间等密切相关,其作用机理如下。
在蛋白质沉积方面。甜菜碱转甲基作用促进机体内甲基代谢,增加RNA含量及其稳定性,提高肌肉中RNA/DNA比例,促进蛋白质生物合成;增加血清中游离Met、Gly和Ser等氨基酸含量[5],为蛋白质合成提供物质基础;诱导机体分泌生长激素(Growth Hormone,GH),改善胰岛素(Insulin,INS)和胰岛素生长因子-Ⅰ(Insulin-like Growth Fac⁃tors-Ⅰ,IGF-Ⅰ)受体信号,促进蛋白质合成与沉积,改善胴体品质[37,41];还能减少用于维持需要的能量[42],提高蛋白质合成效率。在脂肪沉积方面,甜菜碱通过转甲基作用、调节酶活性两条途径,参与体内脂肪代谢,抑制脂肪组织合成,加快脂肪动员,改善胴体品质。甜菜碱可提高甜菜碱合成酶活性,促进高半胱氨酸转化成蛋氨酸,为机体合成肉碱提供充足甲基,增加肉碱在机体肝脏或肌肉中合成量,改善胴体品质。甜菜碱还能通过调节细胞渗透压提高肉组织重量。
Lothong等研究表明,育肥猪饲粮中添加1 250 mg·kg-1甜菜碱可显著降低LSQ指数(背膘厚度与臀中肌宽度的比值)、极显著降低平均背膘厚及血清中甘油三脂浓度[43]。王平等以30~90 kg生长猪为研究对象,发现在饲粮中添加1 000 mg·kg-1甜菜碱可显著降低料肉比,明显提高平均日增重[44]。侯慧丽在饲粮中添加甜菜碱显著提高生长育肥猪日增重,降低料重比[45]。饲粮中添加甜菜碱可显著提高仔猪(断奶后1周)平均采食量和平均日增重,显著增加血清中GH、IGF-Ⅰ含量,当添加量为600 mg·kg-1时,显著降低血清中BUN和LDL含量[4]。

表2甜菜碱对猪生产性能和肉品质的影响Table 2 Effect of betaine on pig performance and pork quality
3.2甜菜碱与肉品质
肉品质评定主要包括肉色、pH、系水力及肌肉脂肪含量等指标。肉色深浅由肌肉中肌红蛋白、色素和血红蛋白量决定,其中肌红蛋白占主导地位。肌肉颜色随肌红蛋白含量增多而加深,原因为琥珀酰-CoA和Gly可影响肌红蛋白合成,Gly是甜菜碱代谢产物,饲粮中补充甜菜碱可增加Gly含量,甜菜碱通过一系列反应间接促进三羧酸循环,提供较多合成卟琳所需底物—琥珀酰- CoA。甜菜碱通过增加Gly和琥珀酰-CoA含量,间接影响肌红蛋白合成。
肉风味源于肌肉内脂肪酸氧化降解的产物,评定较复杂。肌肉中肌苷酸(Inosine Monophosphate,IMP)含量是影响肉风味主要因素之一。甜菜碱通过促进肌肉中脂肪酸β-氧化,生成腺嘌呤核苷三磷酸(Adenosine Triphosphate,ATP),经ATP、腺嘌呤核糖核苷酸(Adenosine Monophosphate,AMP)转化过程,生成肌苷酸(Inosine Monophosphate,IMP),改善猪肉风味。
肌肉pH是评价肉品质关键指标,主要反映动物屠宰后肌糖原酵解速度,受肌肉中乳酸和肌酸含量影响。饲料中添加甜菜碱可减少屠宰后肌肉中乳酸积累量,增加肌肉中肌酸含量,延缓肌肉pH下降[5]。肌肉pH还与肉色、系水力、嫩度等肉质性状高度相关。肌肉pH能改变肌肉蛋白质带电荷状况。肌肉系水力随肌肉蛋白质负电荷减少而降低。当pH下降到接近肌肉蛋白质等电点时,蛋白质的净电荷为零,此时肌肉的系水力最低。此外,肌肉pH越低,肌红蛋白稳定性越差,易被氧化,导致肌肉颜色加深。Matthews等研究发现,甜菜碱可极显著提高69~115 kg肥育猪屠宰后24 h的肌肉pH,但对屠宰后45 min pH无显著影响[46]。
肌肉大理石花纹可反映肌间脂肪含量,与肌肉嫩度、风味、多汁性密切相关,是优质肉的重要标准。当肌束间积聚脂肪时,结缔组织失去弹性,肌束分离,肌肉柔软多汁,易咀嚼,改善肉的风味。王平等在生长育肥猪的饲粮中添加1 000 mg·kg-1甜菜碱,发现背最长肌肌红蛋白、粗脂肪、肌酐酸含量及肌肉大理石纹评分明显升高[44]。肌肉系水力是指肌肉受到外界刺激后保持原有水分的能力,是影响食用品质的重要指标[47]。肌肉系水力常用滴水损失、蒸煮损失等指标衡量,数值取决于肌细胞结构完整程度和蛋白质空间结构,直接关系到肉在贮藏、蒸煮中重量损失。Madeira等发现,饲料中补充甜菜碱可改变生长育肥阶段猪肉的蒸煮损失[48]。甜菜碱通过参与机体脂肪代谢,影响肌间脂肪含量(即肌肉的大理石花纹),改善肌肉系水力。甜菜碱通过改变肌肉pH,使肌肉蛋白收缩、变性甚至降解,吸水力降低。甜菜碱通过促进矿物质沉积与吸收,升高细胞渗透压,增加肌肉系水力。郭建凤等发现[49],在生长育肥猪饲料中添加100和150 g甜菜碱,可使其肉色鲜红,肌内脂肪分布均匀、丰富,不同程度的提高肉色评分、大理石纹评分和肌内脂肪含量,失水率分别降低32.98%和68.01%。
3.3甜菜碱与母体效应
母体效应(Maternal effect)在动物界普遍存在,是指母体在妊娠、哺乳期间,由于自身遗传基因、营养状况、健康情况及外界环境等因素,影响子代性状(能)、特征、行为现象。主要包括mRNA转录、激素分泌、母乳成分和母体抚育行为等,即通过母体遗传基因,影响子代表型;通过改变关键激素和生长因子含量与表达量,间接影响子代出生前及出生后早期的生长发育;通过母体营养作用,影响子代生长发育;通过母体血液供给和母乳营养组分,向子代转移病原或抗体;子代模仿母体行为方式而产生母体效应。在生命初期阶段,母体营养均衡是影响胚胎生长发育的关键因素,影响子代出生前后的生长性能、肌肉发育与代谢、激素分泌和肌内脂肪沉积等[50-52]。研究发现,在母体妊娠期和哺乳期补充甜菜碱,可通过母体效应对子代产生影响[1,19]。
母体妊娠期和哺乳期补充甜菜碱,还可通过降低同型半胱氨酸浓度、增强机体甲基代谢、促进机体激素分泌、改变酶活性等途径,提高母体繁殖性能,对子代出生前及出生后早期的生理状况、新陈代谢等产生影响。首先,甜菜碱作为高半胱氨酸抑制剂,可降低机体高半胱氨酸含量,降低妊娠失败率,提高产仔数。刘大艳等研究表明[53],高半胱氨酸含量升高,严重影响胚胎存活率、致畸率及胎儿正常的生长发育和初生体重等。其次,母体妊娠中期和后期是胎儿肌纤维形成、母体适度生长及乳腺发育的关键时期,也是胎儿生长发育最迅速阶段。在此阶段,胎儿生长需要母体提供大量甲基,甜菜碱是重要甲基来源。甜菜碱作为高效甲基供体,一方面增强机体甲基代谢,促进蛋白质生物合成,改善子代;另一方面,减少肉碱和蛋氨酸的供甲基消耗,节约饲料成本[54]。再次,甜菜碱通过影响母体内HSL、FAS、MDH、LPL等酶的活性,调节母体脂肪代谢,间接对子代脂肪代谢产生影响[19]。Cai等报道[1,19],母体在妊娠期间补充甜菜碱,可调节新生仔猪肝脏中甘油三酯含量以及FAS mRNA表达量,影响子代胆固醇代谢。甜菜碱还可改变机体GH、INS、IGF-Ⅰ等激素水平。机体正常生长发育需要GH、IGF-Ⅰ、Ins、T3、T4等激素相互作用与协调。这些激素不仅对体内蛋白质、糖及脂类代谢具有重要调控作用,还可促进细胞生长、增殖和分化。GH可促进肝脏内蛋白质、DNA和RNA合成,增加血液中游离脂肪酸浓度,降低氨基酸的分解。IGF-Ⅰ可促进胚胎肌肉组织发育、骨骼肌蛋白质合成及骨骼生长。动物缺乏GH时导致IGF-Ⅰ水平低下,表现出生长障碍。Ins是胎儿生长推动激素,含量与胎儿血糖浓度、初生重呈正相关。GH 和Ins均可通过自身作用或者刺激IGF-Ⅰ合成影响胎儿代谢,提高组织合成能力。此外,IGF-Ⅰ还可和Ins共同作用启动卵裂、促进胚泡形成。Nakev等发现[17],IGF-I和LH有协同作用,促进卵泡成熟,增加排卵数,影响胚胎存活数。母猪妊娠期补充甜菜碱,可显著增加新生仔猪肌肉中线粒体COX酶(COX1、COX2和ND5)的活性,显著升高子代血浆中甜菜碱含量和肌肉中甲基转移酶(BHMT and GNMT)表达[38]。
4小结
甜菜碱独特的化学结构及生物学功能,不仅影响机体营养物质代谢,还能有效改善不同生长阶段猪繁殖性能、生产性能、胴体品质和肉品质,通过母子效应传递给子代,调控子代营养。甜菜碱对猪胴体品质和肉品质的改善作用效果显著,但作用机理和调控机理需深入研究。此外,甜菜碱可调控母子两代营养,调控机制仍需系统研究。猪在不同生长阶段对甜菜碱的需要量不同,探讨不同阶段甜菜碱的适宜添加量,具有重要意义。
[参考文献]
[ 1 ] Cai D, Jia Y, Lu J, et al. Maternal dietary betaine supplementa⁃tion modifies hepatic expression of cholesterol metabolic genes via epigenetic mechanisms in newborn piglets[J]. British Journal of Nutrition, 2014, 112(9): 1459-1468.
[ 2 ] Chendrimada T P, Neto M G, Pesti G M, et al. Determination of the betaine content of feed ingredients using high-performance liquid chromatography[J]. Journal of the Science of Food and Agri⁃culture, 2002, 82(13): 1556-1563.
[ 3 ] Zeisel S H, Mar M H, Howe J C, et al. Concentrations of cho⁃line-containing compounds and betaine in common foods[J]. The Journal of Nutrition, 2003, 133(5): 1302-1307.
[ 4 ]董冠.甜菜碱对生长育肥猪生产性能及血清指标影响的研究[D].泰安:山东农业大学, 2012.
[ 5 ]冯杰.甜菜碱对肥育猪生长性能,胴体组成和肉质影响及其作用机理的探讨[D].杭州:浙江农业大学, 1996.
[ 6 ] Deminice R, da Silva R P, Lamarre S G, et al. Betaine supplemen⁃tation prevents fatty liver induced by a high-fat diet: Effects on one-carbon metabolism[J]. Amino Acids, 2015, 47(4): 839-846.
[ 7 ]李炳龙,齐永秀,刘常丽,等.甜菜碱药理作用机制及其药效学研究进展[J].中国新药杂志, 2008, 17(18): 1571-1574.
[ 8 ]张兴会.天然甜菜碱在猪饲料中的应用[J].饲料广角, 2013(15): 49-50.
[ 9 ] Mahmoudnia N, Madam Y. Effect of betaine on performance and carcass composition of broiler chicken in warm weather-a review [J]. International Journal of Agriculture Science, 2012, 2(8): 675-683.
[10] Craig S A S. Betaine in human nutrition[J]. The American Jour⁃na1 of Clinical Nutrition, 2004, 80(3): 539-549.
[11] Shankar R, Murthy S, Pavadi P, et al. Effect of betaine as a feed attractant on growth, survival, and feed utilization in fingerlings of the Indian major carp, Labeo rohita[J]. The Israeli Journal of Aquaculture-Bamidgeh, 2008, 60(2): 95-99.
[12] Olthof M R, Verhoef P. Effects of betaine intake on plasma homo⁃cysteine concentrations and consequences for health[J]. Current Drug Metabolism, 2005, 6(1): 15-22.
[13]冯杰,许梓荣.甜菜碱对肥育猪肌肉和肝脏及血清氨基酸组成的影响[J].浙江大学学报:农业与生命科学版, 2001, 27(1): 107-110.
[14] Eklund M, Bauer E, Wamatu J, et al. Potential nutritional and physiological functions of betaine in livestock[J]. Nutrition Re⁃search Reviews, 2005, 18(1): 31-48.
[15]尹靖东.动物肌肉生物学与肉品科学[M].北京:中国农业大学出版社, 2011.
[16] Kharbanda K K, Todero S L, Ward B W, et al. Betaine administra⁃tion corrects ethanol-induced defective VLDL secretion[J]. Mo⁃lecular and Cellular Biochemistry, 2009, 327(1-2): 75-78.
[17] Nakev J, Popova T, Vasileva V. Influence of dietary betaine sup⁃plementation on the growth performance and carcass characteris⁃tics in male and female growing-finishing pigs[J]. Bulgarian Jour⁃nal of Agricultural Science, 2009, 15: 263-268.
[18]黄其春.甜菜碱对肥育猪脂肪代谢及其关键酶基因表达的影响与机理研究[D].杭州:浙江大学, 2006.
[19] Cai D, Wang J, Jia Y, et al. Gestational dietary betaine supple⁃mentation suppresses hepatic expression of lipogenic genes in neonatal piglets through epigenetic and glucocorticoid recep⁃tor-dependent mechanisms[J]. Biochimica et Biophysica Acta (BBA)-Molecular and Cell Biology of Lipids, 2016, 1861(1): 41-50.
[20]王磊.甜菜碱的渗透压调节作用与动物肠道健康[J].饲料博览, 2013(9): 49-52.
[21] Yang H S, Lee J I, Joo S T, et al. Effects of dietary glycine betaine on growth and pork quality of finishing pigs[J]. Asian-Austral⁃asian Journal of Animal Sciences, 2009, 22(5): 706-711.
[22] Dave Hall. Betaine supplementation for better pig productivity[J]. Pig Progress, 2014, 30(8): 31-32.
[23]赖玉娇,罗海玲,王朕朕,等.甜菜碱的生理学功能及其在畜牧生产中应用的研究进展[J].中国畜牧兽医, 2014, 41(1): 101-107.
[24]贺绍君,赵书景,李静,等.甜菜碱对热应激肉鸡生长性能、十二指肠消化酶活性及盲肠微生物区系的影响[J].动物营养学报, 2014, 26(12): 3731-3739.
[25] Eklund M, Mosenthin R, Tafaj M, et al. Effects of betaine and con⁃densed molasses solubles on nitrogen balance and nutrient digest⁃ibility in piglets fed diets deficient in methionine and low in com⁃patible osmolytes[J]. Archives of Animal Nutrition, 2006, 60(4): 289-300.
[26] Alfieri R R, Cavazzoni A, Petronini P G, et al. Compatible osmo⁃lytes modulate the response of porcine endothelial cells to hyper⁃tonicity and protect them from apoptosis[J]. The Journal of Physi⁃ology, 2002, 540(2): 499-508.
[27] Xu Z R, You Y D. Effect of betaine on digestive function of weaned piglets[J]. Chinese Journal of Veterinary Science, 2000, 20(2): 201-204.
[28] Ratriyanto A, Mosenthin R, Jezierny D, et al. Effect of graded lev⁃els of dietary betaine on ileal and total tract nutrient digestibili⁃ties and intestinal bacterial metabolites in piglets[J]. Journal of Animal Physiology and Animal Nutrition, 2010, 94(6): 788-796.
[29] Mosenthin R, Ratriyanto A, Jezierny D, et al. Intestinal effects of dietary betaine in piglets[J]. Bulletin of University of Agricultural Sciences and Veterinary Medicine Cluj-Napoca. Veterinary Medi⁃cine, 2009, 66(1): 398-403.
[30] Mendoza S M. Sow performance in response to dietary betaine fed in lactation and weaning-to-35 d post-insemination during mod⁃erate heat stress[C]//ADSA-ASAS 2015 Midwest Meeting. Asas, 2015.
[31] Ramis G, Evangelista J N B, Quereda J J, et a1. Use of betaine in gills and sows during Lactation: Effects on milk quality, repro⁃duce five parameters and piglet performance[J]. Journal of Swine Health and Production, 2011, 19(4): 226-232.
[32] van Wettere W H, Herde P, Hughes P E. Supplementing sow ges⁃tation diets with betaine during summer increases litter size of sows with greater numbers of parities[J]. Animal Reproduction Science, 2012, 132(1): 44-49.
[33] Wilcock P. The effect of increasing dietary betaine concentration on starter pig performance[C]//ADSA-ASAS Midwest Meeting. Asas, 2014.
[34]马琳旭,边连全,刘显军,等.甜菜碱对育肥猪生长性能和PRKAG3基因相对表达量的影响[J].黑龙江畜牧兽医, 2012, (7): 23.
[35]王中华,黄修奇.甜菜碱对育肥猪生长性能,胴体品质和肉质的影响[J].中国饲料, 2011(17): 18-20.
[36]边连全,张冬梅,安磊旭,等.肉碱与甜菜碱对育肥猪胴体、肉品质及肝脏营养成分的影响[J].中国饲料, 2009(4): 28-30.
[37]张幸彦,冯涛,许晓玲,等.甜菜碱添加方式对初产母猪繁殖性能、血清生长激素和胰岛素样生长因子-Ⅰ含量的影响[J].动物营养学报, 2015, 27: 2878-2883.
[38] Jia Y, Song H, Gao G, et al. Maternal betaine supplementation during gestation enhances expression of mtDNA-encoded genes through D-loop DNA hypomethylation in skeletal muscle of new⁃born piglets[J]. Journal of Agricultural and Food Chemistry, 2015, 63(46): 10152-10160.
[39] Cai D, Jia Y, Lu J, et al. Maternal dietary betaine supplementa⁃tion modifies hepatic expression of cholesterol metabolic genes via epigenetic mechanisms in newborn piglets[J]. British Journal of Nutrition, 2014, 112(9): 1459-1468.
[40] Rojas-cano M L, Lara L, Lachica M, et al. Influence of betaine and conjugated linoleic acid on development of carcass cuts of Iberian pigs growing from 20 to 50 kg body weight[J]. Meat Sci⁃ence, 2011, 88(3): 525-530.
[41] Cholewa J M, Wyszczelska-Rokiel M, Glowacki R, et al. Effects of betaine on body composition, performance, and homocysteine thiolactone[J]. Journal of the International Society of Sports Nutri⁃tion. 2013, 10(1): 694-709
[42] Schrama J W, Heetkamp M J, Simmins P H, et al. Dietary betaine supplementation affects energy metabolism of pig[J]. Journal of Animal Science, 2003, 81(5): 1202-1209.
[43] Lothong M, Assavacheep P, Tachampa K. Effects of dietary beta⁃ine supplementation on growth performance, carcass quality andserum IGF-Ⅰand triglyceride of finishing pigs. in: kris angka⁃naporn. international graduate research conference[C]. Thailand: Chiang Mai University, 2013: 176-181.
[44]王平.甜菜碱对商品猪生产性能与肉质的影响[J].中国畜牧兽医文摘, 2012(12): 205.
[45]侯慧丽.甜菜碱替代蛋氨酸对生长猪生产性能的影响[J].中国畜牧兽医文摘, 2014(12): 200.
[46] Matthews L O, Southern L L, Higbie A D, et al. Effects of betaine on growth, carcass characteristics pork quality, and plasma metab⁃olites of finishing pigs[J]. Journal of Animal Science, 2001, 79(3): 722-728.
[47]刘立意,刘影,刘冬梅,等.鲜猪肉系水力与应力松弛特性相关性的试验研究[J].东北农业大学学报, 2015, 46(8): 88-93.
[48] Madeira M S, Alfaia C M, Costa P, et a1. Effect of betaine and ar⁃ ginine in lysine-deficient diets on growth, carcass traits, and pork quality[J]. Journal of Animal Science, 2015, 93(10): 4721-4733.
[49]郭建凤,武英,呼红梅.甜菜碱对育肥猪生产性能及胴体的肉质影响[J].家畜生态学报, 2007, 28(2): 21- 24.
[50]梁静.母猪日粮蛋白水平对仔猪脂肪沉积的影响及leptin介导的脂肪代谢调控研究[D].南京:南京农业大学, 2010.
[51]彭济昌.母猪妊娠期能量限饲对子代猪肌纤维发育及肉质的影响[D].哈尔滨:东北农业大学, 2010.
[52]曲艺.低蛋白营养对后代仔猪生产性能和早期肌纤维发育的母体效应[D].哈尔滨:东北农业大学, 2009.
[53]刘大艳,肖立.孕妇羊水同型半胱氨酸、叶酸及VitB12与胎儿畸形的关系[J].吉林医学, 2011, 32(10): 1893-1894.
[54]李玉坤,刘建华,李华文,等.甜菜碱的生物学功能及其在动物生产中的应用[J].中国饲料, 2013(3): 6-11.
Biological functions of betaine and its application for pork quality in swine production
ZHANG Jing, SUN Jinhua, SONG Wentao, SUN Yuecheng, SHAN Anshan (Institute of Animal Nutrition, Northeast Agricultural University, Harbin 150030, China)
Abstract:Betaine was not only affected nutrient metabolism, but also improved swine performance and pork quality of swine in different growth period, and even effect on nutrient regulation of offspring by maternal. The progress in pig performance and pork quality with various levels of betaine in different growth period, which contained the aspects of biological functions, chemical structure, nutrient metabolism and maternal effect of betaine were summarized systematically in this paper.
Key words:betaine; biological function; mechanism; swine; pork quality; maternal effect
*通讯作者:单安山,教授,博士生导师,研究方向为饲料资源开发与利用。E-mail: asshan@neau.edu.cn
作者简介:张婧(1988-),女,博士研究生,研究方向为动物营养与调控。E-mail: 825984292@qq.com
基金项目:现代农业产业技术体系建设专项(CARS-36)
收稿日期:2015-10-27
中图分类号:S816.7;S828
文献标志码:A
文章编号:1005-9369(2016)01-0093-09
