The status of Nrf2-based therapeutics: current perspectives and future prospects
2016-02-09IrinaGazaryanBobbyThomas1DepartmentofCellBiologyandAnatomySchoolofMedicineNewYorkMedicalCollegeValhallaNYUSAandDepartmentofChemistryandPhysicalSciencesDysonCollegePaceUniversityPleasantvilleNYUSA2DepartmentsofPharmacolo
Irina G. Gazaryan, Bobby Thomas1 Department of Cell Biology and Anatomy, School of Medicine, New York Medical College, Valhalla, NY, USA; and Department of Chemistry and Physical Sciences, Dyson College, Pace University, Pleasantville, NY, USA2 Departments of Pharmacology, Toxicology and Neurology, Medical College of Georgia, Augusta University, Augusta, GA, USA
The status of Nrf2-based therapeutics: current perspectives and future prospects
Irina G. Gazaryan1,*, Bobby Thomas2,*
1 Department of Cell Biology and Anatomy, School of Medicine, New York Medical College, Valhalla, NY, USA; and Department of Chemistry and Physical Sciences, Dyson College, Pace University, Pleasantville, NY, USA
2 Departments of Pharmacology, Toxicology and Neurology, Medical College of Georgia, Augusta University, Augusta, GA, USA
How to cite this article:Gazaryan IG, Thomas B (2016) The status of Nrf2-based therapeutics: current perspectives and future prospects. Neural Regen Res 11(11):1708-1711.
Open access statement:This is an open access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 3.0 License, which allows others to remix, tweak, and build upon the work non-commercially, as long as the author is credited and the new creations are licensed under the identical terms.
Funding:This work is supported in part by grants from NIH NS062165, NS060885, Michael J Fox Foundation for Parkinson’s Research, Parfore Parkinson, National Parkinson Foundation (CSRA chapter) to BT and Winifred Masterson Burke Foundation, and Thomas Hartman Foundat ion for Parkinson’s disease to IGG.
This mini-review presents the authors’ vision on the current status and future trends in the development of neuroprotective agents workingviaactivation of nuclear factor erythroid 2-related factor 2 (Nrf2), and in particular,viadisruption of Nrf2-Keap1 interaction. There are two opposite “chemical” mechanisms underlying such activation: the first one is a non-specific covalent modification of Keap1 thiols, resulting in side effects of varied severity, and the second one is the shift of the Nrf2-Kelch-like ECH associated protein-1 (Keap1) binding equilibrium in the presence of a competitive and chemically benign displacement agent. At this point, no displacement activators exhibit sufficient biological activity in comparison with common Nrf2 activators workingviaKeap1 thiol modification. Hence, the hope in therapeutics is now linked to the FDA approved dimethylfumarate, whose derivative, monomethylfumarate, as we demonstrated recently, is much less toxic but equally biologically potent and an ideal candidate for clinical trials right now. A newly emerging player is a nuclear inhibitor of Nrf2, BTB domain and CNC homolog 1 (Bach1). The commercially developed Bach1 inhibitors are currently under investigation in our laboratory showing promising results. In our viewpoint, the perfect future drug will present the combination of a displacement activator and Bach1 inhibitor to insure safety and efficiency of Nrf2 activation.
Nrf2; Keap1; Bach1; electrophiles; oxidative stress; antioxidants
Nrf2 based Therapeutics
A promising strategy for the treatment of neurodegenerative diseases involves pharmacologically induced activation of a coordinated antioxidant genetic program to shift the intracellular redox balance towards a homeostatic state by means of expression of detoxifying proteins and antioxidant enzymes. A key transcription factor orchestrating antioxidant defense is nuclear factor erythroid 2–related factor 2 (Nrf2), a member of the cap’n’collar family of basic leucine zipper transcription factors. In addition to its role in protection from oxidative stress, Nrf2 triggers expression of genes responsible for drug detoxification, iron metabolism, excretion transporters, immunomodulation, calcium homeostasis, growth factors (such as brain derived neurotrophic factor and nerve growth factor-γ), intracellular signaling (including neuronal guanine nucleotide exchange factor), and neurotransmitter receptors (Moi et al., 1994). In addition a recent report on identification and validation of 12 antioxidant response element (ARE)-sequences targeted by Nrf2 in 9 autophagy genes (Pajares et al., 2016) points to additional role of Nrf2 in combating proteinopathies. The breadth of this endogenous response suggests that its activation might counterbalance many of the large number of etiological pathways implicated in the pathogenesis of majority if not all neurodegenerative diseases (Crunkhorn, 2012; Joshi and Johnson, 2012; Gan and Johnson, 2014).
Numerous proof-of-principle studies involving gain and loss of function of Nrf2 have suggested that its induction can ameliorate neurodegeneration, whereas its deficiency can exacerbate neurodegeneration (Joshi and Johnson, 2012; Gan and Johnson, 2014). Despite the well-known fact of Nrf2 activation at late stages of all neurodegenerative diseases, there is insufficient nuclear staining of Nrf2 in the brains of Alzheimer’s (AD) (Ramsey et al., 2007), as well as in aging (senescent animals) (Shenvi et al., 2012) and an increased nuclear staining of Nrf2 in surviving neurons of postmortem Parkinson’s disease (PD) patients (Ramsey et al., 2007). Pre-treatment of pharmacological activators of Nrf2 has been shown to be protective in animal models of numerous neurological conditions such as Huntington’s disease, amyotrophic lateral sclerosis, PD, AD, and cerebral ischemia (Gan and Johnson 2014; Tu et al., 2015). Post-treatment with Nrf2 activators such as sulforaphane and tert-butylhydroquinone has been shown to be beneficial mainlyfor acute injury models such as traumatic brain injury and hemorrhagic stroke, with a very narrow therapeutic window of 30 minutes to 2 hours (Saykally, 2009). We have shown that pre-treatment of Nrf2 activators demonstrate therapeutic effects in animal models of PD. Recently we demonstrated that L-3,4-dihydroxyphenylalanine (L-DOPA) bearing a catechol motif as a weak Nrf2 activator, and nordihydroguaiaretic acid, a potent natural di-catechol activator of Nrf2, is neuroprotective against 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) mouse model of PD (Smirnova et al., 2016). Another recent work of ours that used the FDA approved dimethylfumarate (DMF) and its bioactive metabolite monomethylfumarate (MMF) also showed neuroprotective effects against MPTP-induced PD in mice (Ahuja et al., 2016).
The problem with pharmacologic activation of Nrf2 for the treatment of chronic neurodegenerative disorders originates from the fact that most common Nrf2-based pharmacophores activate Nrf2 by increasing oxidative stress and are harmful chemicals (fore.g., sulforaphane, curcumin, fumarate, triterpenoids, etc.). Increasing oxidative load on already unhealthy cells may significantly counterbalance the benefits of activation of the antioxidant genetic program, making the unambiguous interpretation of the posttreatment results difficult. This is especially true for the MPTP mouse model of PD, where only pre-treatment, but not post-treatment with Nrf2 activators are protective. There is a desperate need in Nrf2 activating chemical tools that are non-electrophilic, non-toxic, and sufficiently stable in the body to unequivocally demonstrate benefits of Nrf2 activation for chronic neurodegenerative diseases. One of the alternative strategies to activate Nrf2 pathway is to bypass the Keap1-Nrf2 interaction by inhibiting BTB domain and CNC homolog 1 (Bach1), which is an Nrf2 inhibitor at the step of transcription complex formation (seeFigure 1). Bach1 inhibition will complement Nrf2 stabilization achievedviadisruption of its complex with Keap1. One may speculate that a combinatorial approach [a) to inhibit Bach1, and b) disrupt Keap1-Nrf2 interaction] is the best way to activate this neuroprotective pathway. The recent investigation on the cause of a significantly lower level of glutamate cysteine ligase (GCLC) pointed to Nrf2 binding from an active ARE to an alternative ARE element, which is not adequate to maintain basal expression of hepatic GCLC in old rats, provides a potential mechanism for the age-related loss of glutathione synthetic and other phase II enzymes. Moreover, the activity at this ARE locus is diminished during aging because of the presence of Bach1 and the absence of CREB-binding protein (CBP), a transcriptional repressor and co-activator, respectively (Shenvi et al., 2012).
There are many Nrf2 activators known from herbal medicines that workviathe alkylation/covalent modification mechanism,e.g., curcumin (turmeric), sulforaphane (broccoli), flavonoids (quercetin, fisetin), nordihydroguaiaretic acid (chapparal). There are multiple herbal supplements on the market based on curcumin (turmeric) in combination with other herbs such as Protandim, Nrf2 Activator (Xymogen), Nrf2 Optimizer (Nuley), Nrf2 Antiox (Progressive Nutracare), and Ultimate Protector, however, there is only one FDA-approved Nrf2 activator – Tecfidera (dimethylfumarate) for the treatment of multiple sclerosis. All currently reported stabilizers of Nrf2 protein workviacovalent modification (alkylation) of Keap1 active cysteines. For example, bardoxolone (oral formulation of a triterpenoid CDDO-Im) is the most potent Nrf2 activator described to date, working in the nanomolar range, likely having a specific binding site in the Keap1 intervening region (IVR) near Cys-298 and Cys-226, as we described (Kaidery et al., 2013). However, it becomes highly toxic in the sub-micromolar range presumably due to covalent and indiscriminate alkylation of multiple proteins required for normal cellular homeostasis (Figure 2).
Both Bardoxolone and Tecfidera (dimethylfumarate) are electrophiles and potent alkylating agents, meaning that they can non-specifically and covalently modify nucleophilic groups in proteins such as cysteine residues. Clinical trials of bardoxolone were discontinued because of patient death http://reatapharma.com/companystatement-termination-of-the-beacon-trial/). Dimethylfumarate is also an alkylating agent, similar to the classic Nrf2 activator sulforaphane, and has been approved by the FDA in 2013 for the treatment of multiple sclerosis (Fox et al., 2012), despite its common side effect of a 30% decline in the lymphocyte count (Sweetser et al., 2013). It has been recently repurposed as a therapeutic against a mouse model of α-synuclein-induced PD (Lastres-Becker et al., 2016). Our recent study demonstrates similar therapeutic benefits for DMF and its bioactive metabolite MMF in anin vivorodent model of PD (Ahuja et al., 2016). Despite these similarities inin vivoefficacy, ourin vitrostudies revealed that MMF is a less potent Nrf2 activator and has much lower toxicity due to the orders of magnitude lesser non-specific alkylating capacity than DMF itself. Our discovery of therapeutic effects of MMF in an experimental PD model without inducing significant alkylating properties compared to DMF suggests that MMF could be a promising candidate for PD therapeutics. Monoethylfumarate (MEF) can also be considered as a therapeutic agent, since its alkylating capacity is very low and is similar to that of MMF. Despite the fact that fumarates are also alkylating agents, their monosubstituted variants such as MMF and MEF will be less toxic than the FDA approved DMF, and may be ideal candidates for future clinical trials in PD. It is obvious that the magic bullet could be a non-alkylating Nrf2 activator, but in the absence of one, MMF and MEF could be the best candidates that can be used as therapeutic agents for PD.
The recently developed Bach1 inhibitor by vTv Therapeutics (High Point NC, USA) represents a novel class of hemeoxygenase 1-inducing compounds. These Bach1 inhibitors are not reactive electrophiles, are not suppressed by N-acetyl cysteine, and do not perturb either ROS or cellular glutathione and are activators of the antioxidant response through the modulation of Bach1 binding to the ARE binding site of target genes (Attucks et al., 2014). Our preliminary results validate Bach1 inhibition as a very effective target against MPTP-induced dopaminergic neurodegeneration by virtue of its ability to activate neuroprotective Nrf2/ARE genetic program (Ahuja et al., 2016). However, more mechanistic and preclinical validation studies in genetic models of PD are required to determine Bach1 inhibition as a safe strategyto activate the Nrf2 pathway.
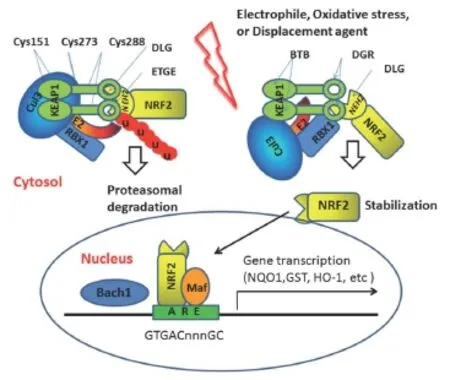
Figure 1 Current model of Keap1/Nrf2/ARE pathway activation.
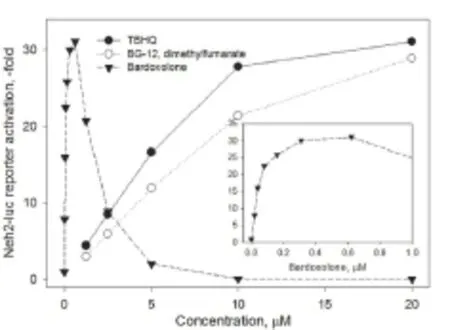
Figure 2 Activation of Neh2-luc reporter– a perfect screening tool for nuclear factor erythroid 2-related factor 2 (Nrf2) activators working via stabilization of Nrf2 protein (Smirnova et al., 2011) –shows a very narrow safe window for bardoxolone at 3 hour incubation in contrast to a wide range of safe biologically active concentrations of dimethylfumarate (DMF).
Nrf2 activation without covalent modification of Keap1 cysteines can be achievedviaNrf2 protein displacement from its complex with Keap1 either by a small molecule, or Nrf2 peptide (Steel et al., 2012). Academic laboratories, small and big pharmaceutical companies are hunting for small molecule Nrf2 activators workingviathis mechanism. Five small molecules have been reported so far, but only two validated (Figure 3). Consistent with this mechanism, more recently a peptide activator of the Nrf2 pathway that functions by disrupting Nrf2-Keap1 interaction was shown to exhibit robust therapeutic effects in a global cerebral ischemia model in rats when the peptide was administered systemically in a post-injury paradigm (Tu et al., 2015). While these findings are highly significant in ameliorating disease pathology in an acute preclinical model, similar peptide-based approaches should be also utilized as a therapeutic strategy for chronic neurodegenerative diseases.
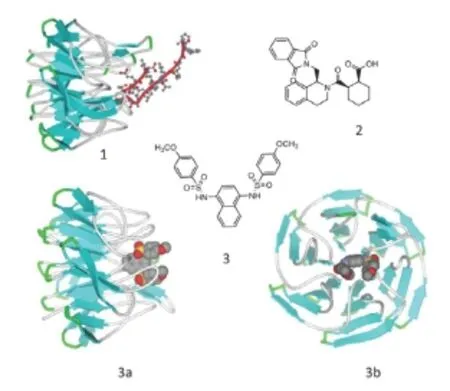
Figure 3 Displacement activators of nuclear factor erythroid 2-related factor 2 (Nrf2).
Compound 1 has been identified in thein vitrohomogeneous assay, re-synthesized, 4 enantiomers separated from each other (compound 1 has 3 chiral centers), and only one enantiomer showed the biological activity: a binding constant of 1 μM to recombinant Keap1, and activation in thein vitrocellular assay only above 10 μM (Hu et al., 2013). Compound 2 has been identified in a joint effort by Biogen Idec, Merrimack Pharmaceuticals, Celgene Corporation (USA), Evotec AG (Germany), and NoValix (France) (Marcotte et al., 2013). It binds Keap1 in thein vitroassay in the low micromolar range, however, in thein vitrocellular assay it works only above 30 μM. There is a shift in potency by at least one order of magnitude when one compares thein vitrobinding constant to Keap1 with the actual performance of the Nrf2 activator in a cellular assay. This offset in potency when one switches from a test-tube to a cell line illustrates the fact that competing with a protein for binding another protein in the real life is far more challenging than competing with a peptide for a recombinant protein in the tube. On the top of their actual low potency, the molecules identified have little chance of crossing blood-brain barrier because of the complexity of the structure bearing multiple cycles and negatively charged groups, predicted easiness of oxidative decomposition in the liver to form active intermediates, and capable of inducing Nrf2 by Keap 1 covalent modification.
One of the future developments could be a combination ofNrf2 displacement activity with mild alkylation potency: in this way the concentration of an alkylating agent will be enriched around Nrf2-Keap1 complex to enhance the specificity of alkylation. The very recent discovery of Nrf2 activators in the low micromolar range among NAD-dependent deacetylase sirtuin-2 (SIRT2) triazole inhibitors (Quinti et al., 2016) may reflect their specific action on Keap1, since in addition to a pro-alkylating motif, they have a stereostructure similar to compound 3 inFigure 3. However, in the absence of a crystal structure, this is just a speculation.
Conclusion
Activation of the Nrf2 signaling pathway is an established mechanism for reducing neurodegeneration due to oxidative stress, and represents a promising therapeutic approach for several neurodegenerative diseases. Unfortunately, Nrf2-based therapeutics has relied on electrophilic pharmacophores. There is indeed a desperate need of safe, potent, blood-brain barrier permeable displacement-type Nrf2 activators as therapeutic agents for chronic neurodegenerative diseases. Given the very low potency of existing displacement-type Nrf2 activators and their complex chemical structures that are not ideal for them to cross the blood-brain barrier, it is going to be an arduous task to develop such compounds as therapeutic agents. At this point MMF, which upon hydrolysis gives a natural metabolite – fumaric acid, has a low alkylating potency and thus, low toxicity, could be the best option to treat chronic neurodegenerative disease such as PD. More studies should be carried out to test the therapeutic effects of peptidebased inhibitors of Nrf2-Keap1 interaction as therapeutic agents in preclinical models of chronic neurodegenerative diseases. Furthermore, efforts should also focus on validating Bach1 inhibition as an alternate strategy for safe and efficient activation of the Nrf2 pathway as a novel therapeutic target for PD and related neurodegenerative diseases.
Author contributions:IGG and BT wrote and revised the manuscript. IGG created the figures.
Conflicts of interest:None declared.
Ahuja M, Ammal Kaidery N, Yang L, Calingasan N, Smirnova N, Gaisin A, Gaisina IN, Gazaryan I, Hushpulian DM, Kaddour-Djebbar I, Bollag WB, Morgan JC, Ratan RR, Starkov AA, Beal MF, Thomas B (2016) Distinct Nrf2 signaling mechanisms of fumaric acid esters and their role in neuroprotection against 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-induced experimental Parkinson’s-like disease. J Neurosci 36:6332-6351.
Ahuja M., Kaidery NA, Morgan J, Igarashi K, Attucks OC, Thomas B. (2016) Preclinical validation of Bach1 inhibition for the development of Parkinson’s disease therapy. Society for Neuroscience 2016 Session 787 - Parkinson’s Neuroprotection II, 787.21/I9.
Attucks OC, Jasmer KJ, Hannink M, Kassis J, Zhong Z, Gupta S, Victory SF, Guzel M, Polisetti DR, Andrews R, Mjalli AM, Kostura MJ (2014) Induction of heme oxygenase I (HMOX1) by HPP-4382: a novel modulator of Bach1 activity. PLoS One 9:e101044.
Crunkhorn S (2012) Deal watch: Abbott boosts investment in NRF2 activators for reducing oxidative stress. Nat Rev Drug Discov 11:96.
Fox RJ, Miller DH, Phillips JT, Hutchinson M, Havrdova E, Kita M, Yang M, Raghupathi K, Novas M, Sweetser MT, Viglietta V, Dawson KT (2012) Placebo-controlled phase 3 study of oral BG-12 or glatiramer in multiple sclerosis. N Engl J Med 367:1087-1097.
Gan L, Johnson JA (2014) Oxidative damage and the Nrf2-ARE pathway in neurodegenerative diseases. Biochim Biophys Acta 1842:1208-1218.
Hu L, Magesh S, Chen L, Wang L, Lewis TA, Chen Y, Khodier C, Inoyama D, Beamer LJ, Emge TJ, Shen J, Kerrigan JE, Kong AN, Dandapani S, Palmer M, Schreiber SL, Munoz B (2013) Discovery of a small-molecule inhibitor and cellular probe of Keap1-Nrf2 protein-protein interaction. Bioorg Med Chem Lett 23:3039-3043.
Joshi G, Johnson JA (2012) The Nrf2-ARE pathway: a valuable therapeutic target for the treatment of neurodegenerative diseases. Recent Pat CNS Drug Discov 7:218-229.
Kaidery NA, Banerjee R, Yang L, Smirnova NA, Hushpulian DM, Liby KT, Williams CR, Yamamoto M, Kensler TW, Ratan RR, Sporn MB, Beal MF, Gazaryan IG, Thomas B (2013) Targeting Nrf2-mediated gene transcription by extremely potent synthetic triterpenoids attenuate dopaminergic neurotoxicity in the MPTP mouse model of Parkinson’s disease. Antioxid Redox Signal 18:139-157.
Lastres-Becker I, García-Yagüe AJ, Scannevin RH, Casarejos MJ, Kügler S, Rábano A, Cuadrado A (2016) Repurposing the NRF2 activator dimethyl fumarate as therapy against synucleinopathy in Parkinson’s disease. Antioxid Redox Signal 25:61-77.
Marcotte D, Zeng W, Hus JC, McKenzie A, Hession C, Jin P, Bergeron C, Lugovskoy A, Enyedy I, Cuervo H, Wang D, Atmanene C, Roecklin D, Vecchi M, Vivat V, Kraemer J, Winkler D, Hong V, Chao J, Lukashev M, Silvian L (2013) Small molecules inhibit the interaction of Nrf2 and the Keap1 Kelch domain through a non-covalent mechanism. Bioorg Med Chem 21:4011-4019.
Moi P, Chan K, Asunis I, Cao A, Kan YW (1994) Isolation of NF-E2-related factor 2 (Nrf2), a NF-E2-like basic leucine zipper transcriptional activator that binds to the tandem NF-E2/AP1 repeat of the beta-globin locus control region. Proc Natl Acad Sci U S A 91:9926-9930.
Pajares M, Jiménez-Moreno N, García-Yagüe ÁJ, Escoll M, de Ceballos ML, Van Leuven F, Rábano A, Yamamoto M, Rojo AI, Cuadrado A (2016) Transcription factor NFE2L2/NRF2 is a regulator of macroautophagy genes. Autophagy 12:1902-1916.
Quinti L, Casale M, Moniot S, Pais TF, Van Kanegan MJ, Kaltenbach LS, Pallos J, Lim RG, Naidu SD, Meisel L, Rauf NA, Leyfer D, Maxwell MM, Saiah E, Landers JE, Luthi-Carter R, Abagyan R, Dinkova-Kostova AT, Steegborn C, Marsh JL, et al. (2016) SIRT2- and NRF2-targeting thiazole-containing compound with therapeutic activity in Huntington’s disease models. Cell Chem Biol 23:849-861.
Ramsey CP, Glass CA, Montgomery MB, Lindl KA, Ritson GP, Chia LA, Hamilton RL, Chu CT, Jordan-Sciutto KL (2007) Expression of Nrf2 in neurodegenerative diseases. J Neuropathol Exp Neurol 66:75-85.
Saykally JN, Rachmany L, Hatic H, Shaer A, Rubovitch V, Pick CG, Citron BA (2012) The nuclear factor erythroid 2-like 2 activator, tert-butylhydroquinone, improves cognitive performance in mice after mild traumatic brain injury. Neuroscience 223:305-314.
Shenvi SV, Smith E, Hagen TM. (2012) Identification of age-specific Nrf2 binding to a novel antioxidant response element locus in the Gclc promoter: a compensatory means for the loss of glutathione synthetic capacity in the aging rat liver? Aging Cell 11:297-304.
Smirnova NA, Haskew-Layton RE, Basso M, Hushpulian DM, Payappilly JB, Speer RE, Ahn YH, Rakhman I, Cole PA, Pinto JT, Ratan RR, Gazaryan IG (2011) Development of Neh2-luciferase reporter and its application for high throughput screening and real-time monitoring of Nrf2 activator. Chem Biol 18:752-765.
Smirnova NA, Kaidery NA, Hushpulian DM, Rakhman II, Poloznikov AA, Tishkov VI, Karuppagounder SS, Gaisina IN, Pekcec A, van Leyen K, Kazakov SV, Yang L, Thomas B, Ratan TT, Gazaryan IG (2016) Bioactive flavonoids and catechols as Hif1 and Nrf2 protein stabilizers - implications for Parkinson’s disease. Aging Dis doi:10.14336/ AD.2016.0505.
Steel R, Cowan J, Payerne E, O’Connell MA, Searcey M (2012) Anti-inflammatory Effect of a Cell-Penetrating Peptide Targeting the Nrf2/ Keap1 Interaction. ACS Med Chem Lett 3:407-410.
Sweetser MT, Dawson KT, Bozic C (2013) Manufacturer’s response to case reports of PML. N Engl J Med 368:1659-1661.
Tu J, Zhang X, Zhu Y, Dai Y, Li N, Yang F, Zhang, Q, Brann DW, Wang R (2015) Cell-Permeable peptide targeting the Nrf2-Keap1 interaction: A potential novel therapy for global ceberal ischemia. J Neurosci 35:14727-14739.
*Correspondence to: Irina G. Gazaryan, Ph.D. or Bobby Thomas, Ph.D., igazaryan@pace.edu or bthomas1@augusta.edu.
10.4103/1673-5374.194706
Accepted: 2016-10-30
杂志排行

中国神经再生研究(英文版)的其它文章
- Six psychotropics for pre-symptomatic & early Alzheimer’s (MCI), Parkinson’s, and Huntington’s disease modification
- The Epigenome as a therapeutic target for Parkinson’s disease
- Neuroinflammation, neurodegeneration and regeneration in multiple sclerosis: intercorrelated manifestations of the immune response
- Applicability of tooth derived stem cells in neural regeneration
- Cortical spreading depression-induced preconditioning in the brain
- Targeting neuronal nitric oxide synthase as a valuable strategy for the therapy of neurological disorders