云南糯玉米种质资源的研究与利用
2016-02-06雷涌涛隆文杰周国雁伍少云
雷涌涛,隆文杰,周国雁,蔡 青,伍少云
(云南省农业科学院 生物技术与种质资源研究所/云南省农业生物技术重点实验室/农业部西南作物基因资源与种质创制重点实验室,云南 昆明 650223)
云南糯玉米种质资源的研究与利用
雷涌涛,隆文杰,周国雁,蔡 青*,伍少云*
(云南省农业科学院 生物技术与种质资源研究所/云南省农业生物技术重点实验室/农业部西南作物基因资源与种质创制重点实验室,云南 昆明 650223)
云南省是糯玉米的起源中心和遗传多样性中心,糯玉米种质资源丰富,品种类型多样,分布广泛。全省16个地州(市)78个县(市)都分布有糯玉米地方品种,西双版纳傣族自治州勐海县至今仍种植有被公认为具有一系列原始性状的地方糯玉米品种四路糯。从糯玉米地方品种的生态型或多样性、支链淀粉含量变化、过氧化物同工酶带型特征、分子标记揭示的遗传多样性等方面,回顾了相关研究机构和研究者对云南省糯玉米种质资源的研究,分析了勐海四路糯玉米与缅甸糯玉米、锡金原始玉米的关系,以及糯玉米在发展云南高原特色农业中的作用和面临的主要问题。
糯玉米; 种质资源; 研究与利用; 多样性; 云南
玉米是云南省的主要粮食作物之一,常年播种面积为133.33万hm2左右,其总产量的65%~70%用于饲粮,作为口粮、食品工业原料以及工业综合利用的产量只占30%~35%[1]。糯玉米属于玉蜀黍属(ZeaL.)玉蜀黍亚属(SectionZea)玉米种(Zeamays)栽培玉米亚种(ssp.mays)[2]。其用途广泛,营养丰富,经济价值较普通玉米高,是发展高产、优质、高效农业的重要作物之一。除其鲜嫩果穗是大众青睐的美味食物外,由其生产的淀粉或其变性淀粉是制糖、酿造、食品和黏合剂生产以及纺织、造纸和制药等轻工业领域广泛应用的原料[3-4]。但是,糯玉米等优质鲜食玉米在云南省的种植面积并不大,2007—2011年,年均种植面积仅6.67万hm2[1](含甜玉米),仅占当地玉米常年播种面积的5%左右。然而,由于云南省的许多少数民族都有喜欢食用糯性食物的饮食习俗或文化,保留了许多糯性食物的地方品种或类型,如糯稻、糯玉米、糯高粱和糯薏苡等。所以,云南省有丰富的糯玉米种质资源。为利用特色农业生物资源,发展高原特色农业和民族经济,简要回顾了有关研究机构、农业科技研究人员在云南糯玉米种质资源的品种类型或多样性、淀粉含量变化、过氧化物同工酶以及分子标记及其揭示的遗传多样性方面的研究进展,分析了云南勐海四路糯在中国糯玉米起源中的地位及其同缅甸糯玉米、锡金原始玉米的关系,提出了糯玉米种植业在发展云南高原特色农业中的作用及面临的主要问题。
1 云南糯玉米种质资源的多样性
1.1 云南糯玉米的生态型和品种多样性
1.2 云南糯玉米淀粉含量的变化
在上述提及的320份糯玉米地方种质资源材料中,测定淀粉含量的共64份。其中,支链淀粉含量占总淀粉含量73.11%~77.09%的5份,为64份材料的7.81%,其总淀粉含量为67.62%~71.24%,直链淀粉占其中的22.91%~26.89%;支链淀粉含量占81.20%~88.92%的6份,为64份材料的9.38%,其总淀粉含量为66.29%~70.99%,直链淀粉占其中的11.08%~18.80%;支链淀粉含量占90.43%~94.30%的4份,为64份材料的6.25%,其总淀粉含量为65.61%~68.95%,直链淀粉占其中5.70%~9.57%;剩余49份的支链淀粉含量为95.12%~99.64%,总淀粉含量为64.8%~70.98%,直链淀粉占总淀粉含量的0.36%~4.88%。
云南糯玉米地方品种的支链淀粉和直链淀粉的含量与其籽粒颜色(用数字1代表白色、2代表黄色、3代表红色、4代表黑色)呈极显著相关,r分别为-0.680和0.680;与品种来源地的海拔、相对湿度、纬度呈显著相关,r分别为0.218和-0.218、-0.237 和0.237、0.231和-0.231[14]。因此,云南糯玉米地方种质资源不仅支链淀粉含量的变化范围大,支链淀粉含量高、糯性强的品种较多,而且原产地海拔、纬度甚至湿度对其支链淀粉含量的影响显著,海拔、纬度越高的品种,其支链淀粉的含量也越大。
1.3 云南糯玉米过氧化物同工酶的谱带特征
同工酶是不同遗传位点的产物,或是单个多肽次级结构发生改变的结果,常被遗传学家用于研究植物群体内的遗传多型性、种内和种间的遗传差异、发育过程中分化基因的表达、基因剂量对酶结构及其功能的影响、基因内和基因间的互补、杂种优势以及基因与有机体进化等[15]。因此,同工酶可以用于研究不同种、属的植物间的亲缘关系及起源问题。在玉米中,与胚乳酯酶同工酶相比,叶片过氧化物同工酶的第4、5谱带色浓、粗,在显色过程中最早出现,且显色最清楚,迁移率相当稳定,在不同的品种间也有明显的差异[15],可以用于研究不同类型玉米的关系。在勐海四路糯[6,16]及来自云南其他地方的15个糯玉米地方品种中,四路糯及其他11个品种占参试材料的75%,都只具有中国玉米特有的过氧化物同工酶的第5特征酶带,且未发现群体内不同植株之间的差异[16];2个品种具有混合谱带,另2个品种则只有美洲玉米有的特征带第4带[16]。另外,从云南西双版纳征集到的19份糯玉米品种也都只具有第5带[17]。云南新平红糯玉米、屏边白糯玉米、蒙自早白糯和贵州麻江双头糯、安龙长冲白糯、兴义大白糯等9份品种也只具有第5带[18]。85.3%的云南糯玉米和94.7%的西双版纳糯玉米,以及89.5%的广西糯玉米都具有第5带[16,19]。因此,西双版纳糯玉米的酶谱纯度更高[19]。
1.4 云南糯玉米的分子标记研究
1.4.1 分子标记揭示的云南糯玉米地方种质资源的遗传多样性 田孟良等[18,20]用79条SSR分子标记引物,研究了贵州普通玉米、贵州糯玉米及云南糯玉米之间的遗传差异或关系。结果表明,糯玉米和普通玉米在DNA分子水平上存在较大的差异,而且这种差异发生在整个基因组当中,而不只是出现在wax基因及其相关的位点上。因而不支持糯玉米是由普通玉米经单基因突变而形成[21]的观点。SSR标记聚类分析把云南屏边白糯玉米、蒙自早白糯玉米、5份贵州糯玉米及1份贵州普通玉米划分在同一群,而将云南新平红糯玉米单独分为一群,说明新平红糯玉米与所有参试材料的遗传距离最大,亲缘关系最远。刘永建[22]用64条SSR分子标记引物,研究了15份云南糯玉米(包括勐海四路糯)、30份贵州糯玉米的遗传多样。结果表明,云南省和贵州省的糯玉米群体(品种)平均遗传多样性指数分别为1.144 7和1.247 2,遗传多样性水平相当,不同品种的平均遗传多样性指数在0.085 1(勐海四路糯)~0.705 3(新平红糯玉米),群体间的遗传多样性比群体内个体(单株)间的遗传多样性更丰富,大多数云南糯玉米(66.7%)和大多数贵州糯玉米(70%)被分在一群内,表明它们之间的亲缘关系更近。
2.3 2007-2011年西城区协助北京市新生儿疾病筛查中心(以下称新筛中心)追访情况 5年中北京市新筛中心仅10%的可疑病例需要区级层面协助追访,其中PKU可疑异常4例,CH可疑异常31例,追访后复诊率分别为83.87%和50.00%。见表2。
吴渝生等[12]利用61对SSR引物,分析了代表云南不同生态区域的16份糯玉米地方种质的遗传多样性。结果表明,82.0 %的引物能检测出片段大小为70~320 bp的2~7个等位基因,云南省的糯玉米地方品种不仅具有较高的遗传多样性,而且能被划分成海拔2 400~3 300 m、1 700~2 200 m和1 000~1 300 m的3个类群及滇西北、滇东北、滇西—滇中、滇南和滇东南—滇西南5个亚群。很明显,这个聚类结果基本上符合云南不同海拔地势的变化走向。由此可见,云南糯玉米种质资源的遗传多样性是与海拔、气候等生态条件紧密相连的。该结论从分子生物学层面支持张建华等[10-11,14]以形态和生态区域对云南糯玉米地方品种的分类研究结果。张金渝等[23-24]用SSR标记研究了37份云南糯玉米地方种质和3个不同来源且形态差异明显的勐海四路糯的遗传多样性。结果表明,云南糯玉米地方品种的遗传基础广泛,37份材料可被划分为5个群13个亚群,64.86%的大多数材料同国内5个自交系杂种优势群的亲缘关系较远,被划分在这5个优势群外的其他3个群5个亚群中。在3份勐海四路糯玉米中,总的基因多样性(HT)为0.183 0,居群内(同一来源的四路糯)的基因多样性(Hs)为0.142 2。因此,居群内的基因多样性远较居群间(不同来源的四路糯间)的基因多样性高。吴斌等[25]用100对多态性好、稳定性高的SSR引物,分析了采自西双版纳的36份小糯玉米地方品种和2份对照品种的遗传多样性。结果表明,虽然36份糯玉米地方品种能被划分成6个不同的类群,但这些类群都独立于对照品种,表明它们与对照品种的遗传差异大、亲缘关系较远。
1.4.2 云南糯玉米waxy基因序列多态性及其突变类型 玉米之所以表现出糯质,是因为转座子及反转录转座子插入到waxy基因侧翼区、内含子、外显子之中,或者是因为waxy基因内部发生碱基缺失,而使直链淀粉合成受阻[5]所致。目前,在美洲糯玉米中已发现了近50种糯质突变类型[5]。
田孟良等[5]对包括云南蒙自早白糯玉米、镇康偏头糯玉米、屏边花糯玉米、屏边白糯玉米和镇康烂地花糯玉米等在内的16份中国西南糯玉米的waxy基因,第9—14外显子之间的核苷酸序列进行了测定。结果表明,中国西南糯玉米、西南硬粒玉米、小颖玉米分别具有14、19、40个多态性位点,糯玉米waxy位点的核苷酸多态性只是硬粒玉米的30%和小颖玉米的14%。序列比对发现,16份糯玉米中12份的第10外显子的同一位置发生了15个核苷酸的缺失。这个15 bp的缺失突变被称之为wx-D10突变类型。该位点是糖基转移酶(glucosyl transferase domain 1,GTD1) 结构域的起始位置,此处缺失导致GTD1功能丧失而表现出糯质。赵耀[26]也测定了包括云南蒙自早白糯玉米、镇康偏头糯玉和四路糯玉米在内的6份西南糯玉米的waxy基因编码区序列,通过比对发现,四路糯玉米也和田孟良等[5]在早白糯玉米、偏头糯玉米、白糯玉米和烂地花糯玉米中的发现一样,是wx-D10缺失突变型。除此外,在云南蒙自早白糯、镇康偏头糯和贵州安龙长冲白糯的第9外显子内还发现一段16 bp的重复插入序列,这是西南糯玉米waxy基因的一种新的突变型。比对发现,该序列位于glycogen synthase结构域,此处发生突变,可能导致糖原合成酶失活。另外发现,镇康偏头糯在第2内含子结尾处由正常碱基AG突变成了AT,这可能是西南糯玉米waxy基因的又一种新的突变型,该突变属剪切位点的碱基突变。刘汉梅[27]则测定包括云南蒙自早白糯玉米、镇康偏头糯玉米和四路糯玉米在内的6份西南糯玉米waxy基因编码区的全长序列,比对发现,这些糯玉米品种除是wx-D10缺失突变型外,偏头糯分别在第2内含子的剪接受体位点发生了由AG向AT的突变,在第2外显子多次发生了不同碱基的插入突变,早白糯和四路糯分别在第2外显子正常编码区发生了GC碱基的缺失和AGCC四碱基的插入移码突变,在第9外显子区域,早白糯和偏头糯发生了一段16 bp的重复插入突变。
王勇斌[28]以自交系黄早四和478为对照,分析了云南花糯、白糯等8份西南糯玉米地方品种与普通玉米之间的waxy基因的表达差异及糯玉米waxy基因的cDNA序列。结果表明,与普通玉米相比,糯玉米胚乳中无Wx蛋白(颗粒结合淀粉合成酶,Granule-bound starch synthases,GBSS)的合成,使waxy基因的表达降低,两者间的差异主要受控于转录水平;对糯玉米waxy基因的cDNA序列分析发现,花糯、保山白糯等7个糯玉米在转运肽区域插入了3个碱基(CAC)、柳城糯插入了9个碱基(CACCAGCAG),使相应的氨基酸序列由普通玉米的Gly突变为糯玉米的Ala。这种错义突变可能改变了转运肽的功能。另外,花糯、保山白糯等7个糯玉米的waxy基因在第2外显子存在有同义突变,使编码Gly的密码子由普通玉米的GGC突变为糯玉米的GGA;而在第8外显子,8份糯玉米的waxy基因也都有同义突变,使编码Pro氨基酸的密码子从普通玉米的CCG突变为糯玉米的CCC,在第10外显子则都发了生15 bp的缺失,导致了wx-D10突变型。姚坚强等[29]对来自中国、泰国和韩国的325份糯玉米种质资源材料的突变类型进行了鉴定。结果表明,在309份中国糯玉米中,有299份(占96.8%)表现为wx-D7和wx-D10突变型,另有10份(地方种质和自交系各5份)为其他突变类型,waxy基因的遗传多样性很低。其中,中国北方的204份及韩国的5份糯玉米都是wx-D7突变型,而南方的95份及泰国的11份则都是wx-D10突变型;在90份地方种质资源中,71份(占78.9%,其中云南16份)为wx-D10突变型,而在62份商业杂交种和157份自交系中分别有54份和136份表现为wx-D7突变型,占87.1%和86.6%。
2 勐海四路糯在糯玉米起源中的地位及与其他原始玉米的关系
2.1 勐海四路糯玉米在糯玉米起源中的特殊地位
自1908年,原产中国云南的糯玉米被美国传教士法哈姆(Famharm)通过上海领事传入美国,并被植物学家柯林斯(Collins)首次详细描述和报道其特征后,糯玉米起源于中国就已经被各国学者所公认。然而,关于其起源地点、起源方式和起源时间至今也有不同的观点。
勐海四路糯在确定糯玉米起源于云南西双版纳和广西的热带、亚热带地区的合理推测与研究中有着举足轻重、不可替代的作用[21,30]。它是曾孟潜等[6]于1970年从云南省西双版纳傣族自治州勐海县一带征集到的农家品种,具有果穗顶端雄花枝梗长且有两性花、单株果穗数多而穗行数少、果穗柄长而籽粒被颖片半包裹、植株矮分蘖多而气生根发达等一系列的原始性状。这些原始特性恰好符合科学家设想的玉米野生祖先的主要特点[6]。另外,它具有中国薏苡、糯质及硬粒型玉米过氧化物同工酶的典型特征——第5带,而不是引自美洲的马齿玉米及美洲糯玉米的第4带[6,16,19]。李艳天等[31]、曾孟潜[30]和孙建英[21]认为中国糯玉米的形成时间早于1760年以前,而四路糯则是自1890年以来就是当地傣族、哈尼族、拉祜族和其他少数民族一直在种植的糯玉米品种之一[6]。
除原始性状和同工酶特殊带型外,分子标记及waxy基因测序研究的结果也支持勐海四路糯在糯玉米起源方面的特殊作用。田孟良[7]对勐海四路糯玉米、有稃玉米、甜质玉米和马齿玉米的ITS序列,以及多年生玉米、二倍体多年生玉米、小颖玉米、墨西哥玉米和美洲栽培玉米的ITS序列的比对研究表明,四路糯具有假基因(Zeapseudogenes)性质的祖先序列。刘坚[32]对23份北方、4份四路糯及其他22份南方糯玉米waxy基因部分片段的测序分析表明,四路糯同南方糯玉米的waxy基因序列完全一致,认为四路糯可能是南方糯玉米生态类型的祖先。
在20世纪70年代,四路糯玉米在云南省勐海县、景洪县和勐腊县等都有分布,但勐海县勐海镇的曼贺村和团结寨的种植较多[6]。20世纪80年代初,即“六五”期间(1981—1985年),云南省农业科学院从勐海县收集到1份名为勐海四路糯的玉米地方品种(保存编号为231272-1025)。其主要性状特征为:秆高不足200 cm、糯质、穗长11 cm左右、穗行数4~8、千粒质量不足70 g。21世纪初,云南省农业科学院再次从勐海县收集到1份也名为勐海四路糯的玉米地方品种(232019-2138),但该材料在昆明种植时株高达250 cm以上、穗位高150 cm以上、糯质、穗长16 cm左右、穗行数10~12、千粒质量140 g左右。在此期间,云南省德宏傣族景颇族自治州农科所顾中量先生,从瑞丽边境缅甸一侧收集到1份名为四棱玉米(232231-2350)的特糯玉米地方品种,并将其交由云南省农业科学院种植观察和保存。在昆明,该材料表现为花丝红色、花药褐色、秆高130 cm左右、穗位高50 cm左右、穗长10 cm左右、穗行数4~6、行粒数20左右、千粒质量170 g左右。2007年, “云南及周边地区农业生物资源调查(2006FY110700)”项目勐海县调查队,在勐海县勐遮镇曼扫村允竜组也采集到1份名为四棱玉米的糯玉米(采集号2007532221,云南保存编号2802)。在昆明,该材料表现花丝红色、花药黄色、秆高140 cm左右、穗位高1 m左右、果穗多穗小、每株结实4个果穗、穗行数4~8、糯质、千粒质量仅80 g左右。从表型性状看,保存编号为232019-2138的四路糯不符合曾孟潜等[6]描述的勐海四路糯玉米的主要特征,而编号为231272-1025的四路糯、编号为232231-2350和采集号为2007532221的四棱玉米则基本符合。但是,这3份材料和1970年曾孟潜等[6]收集到的勐海四路糯玉米是否是同名异物或是同物异名,以及它们在糯玉米起源与演化中的作用和地位有待进一步研究和确定。
2.2 勐海四路糯玉米与缅甸糯玉米及锡金原始玉米的关系
勐海四路糯是1970年发现的农家糯玉米品种[6],现在又在勐海县及瑞丽对面的缅甸境内收集到农艺性状、特别是果穗大小及穗行数与其酷似的四棱糯玉米。如果三者确系同物异名,而缅甸北部栽种四棱玉米也如勐海县少数民族种植四路糯玉米的历史一样悠久,那么糯玉米的起源地至少应从西双版纳延伸至缅甸东北部,而不是田孟良[7]推测的仅限于中国云南、贵州及广西3省的结合部(23°~25°N、105°E附近)。
在喜马拉雅山东北发现的锡金原始玉米,其主要特征为:植株高大,一般在200 cm以上;多果穗,每株结4~8穗;穗位高,多在茎上部;上位穗顶生雄花,穗瘦小、呈圆筒形、顶端渐尖、长6~12 cm;穗行数8~10,籽粒不规则排列、乳白色至黄色,尖顶、爆粒型;植株分蘖力强等[33]。这些特征与考古学家们根据墨西哥出土的玉米穗和部分植株遗存,描绘出的史前原始玉米的主要特征:植株结多个果穗且大小一致;穗小、瘦长圆形,籽粒排列不规则、圆形尖顶、爆粒种;果穗顶生雄花;植株有分蘖力等[33]极其相似。据研究,与马齿型、硬粒型和粉质型玉米相比,糯玉米的核型与公认较原始的爆裂型和有稃型玉米相似,都具有基本对称的核型[34],具有这种较对称核型的植物在系统演化上一般都是处于较古老、原始地位的物种,进化程度较高的植物则会出现非对称的核型,所以糯玉米比马齿型、硬粒型玉米更加原始[17]。无论锡金原始玉米或是考古学家描绘的史前原始玉米都有一个共同的特征,即多穗、穗小和爆裂型。云南省有不少爆裂玉米地方品种,吴渝生等[12]研究了其中14份种质的遗传多样性,并把它们分成了海拔2 400~2 600 m、1 700~2 200 m 和1 000~1300 m 3个群和滇中—滇西、滇西、滇南和滇东南4个亚群。目前,云南省作物种质中期库保存有爆裂玉米种质资源近40份,其中耿马八苞玉米(亦称手指玉米或炸花玉米,保存编号2579/3041)秆高260 cm左右、穗位高约160 cm、穗行数12~14、单株多则可结8穗、千粒质量150 g左右,与锡金原始玉米和考古学家描绘的史前原始玉米的主要特点有相似之处。刘永建[22]研究了勐海四路糯等15份云南糯玉米、30份贵州糯玉米与云南鲁甸爆裂玉米之间的遗传关系,结果表明,糯玉米与爆裂玉米间的遗传相似性较高,遗传距离较小;而糯玉米与普通玉米间的遗传相似性较小,遗传差异较大;聚类分析也把66.7%的云南糯玉米、70%的贵州糯玉米及爆裂玉米分在一群,表明糯玉米与爆裂玉米之间有更亲密的遗传关系。
到目前为止,对玉米细胞学核型研究的结果[17,34]和少量分子生物学研究的结果[22],支持糯玉米与爆裂玉米有更加亲近的关系。大多数分子生物学研究,可能由于试验选材问题,得出了支持或者不支持糯玉米由硬粒玉米或马齿玉米经单基因突变[21]演化而来[18,35]的观点。因此,糯玉米的起源或演化路径有待深入研究。
3 糯玉米种植在发展云南高原特色农业中的作用和面临的主要问题
糯玉米用途广泛,其青果穗可蒸、煮鲜食或籽可菜用,采青果穗后的秆、叶是饲养牲畜的优质饲料,而干籽粒可加工造纸、纺织、粘着剂、食品及医药等轻工业发展所需的糯性淀粉,因此糯玉米是发展云南高原特色农业、增强农村经济实力的多用作物之一。云南山地多、水田少,耕地坡度大,水利灌溉设施条件差,农业生产抵御干旱风险的能力弱,适宜种植玉米等抗旱节水的旱地作物。另外,云南保山市、临沧市、普洱市、西双版纳州、红河州和文山州等热带、亚热带地区的光、热、水资源充足,具有发展热区农业、冬季农业的气候优势,这些地区周年可种植玉米。再有,云南糯玉米种资源丰富、类型多样、支链淀粉含量高、糯性强、口感好的地方品种众多。因此,云南具有发展糯玉米种植业得天独厚的气候优势和生物资源优势。
据报道,2008年全国糯玉米的种植面积为48.93万hm2,鲜果穗总产量达567.6万t[36]。其中,种植面积最大的是吉林省,为14.67万hm2,占全国总种植面积的30.0%(表1)。2013年,全国糯玉米的种植面积已达到66.67万hm2以上[3],而云南省2011年包含甜玉米在内仅有6.67万hm2左右[4],且甜玉米在2008年时已有4万hm2[36]。
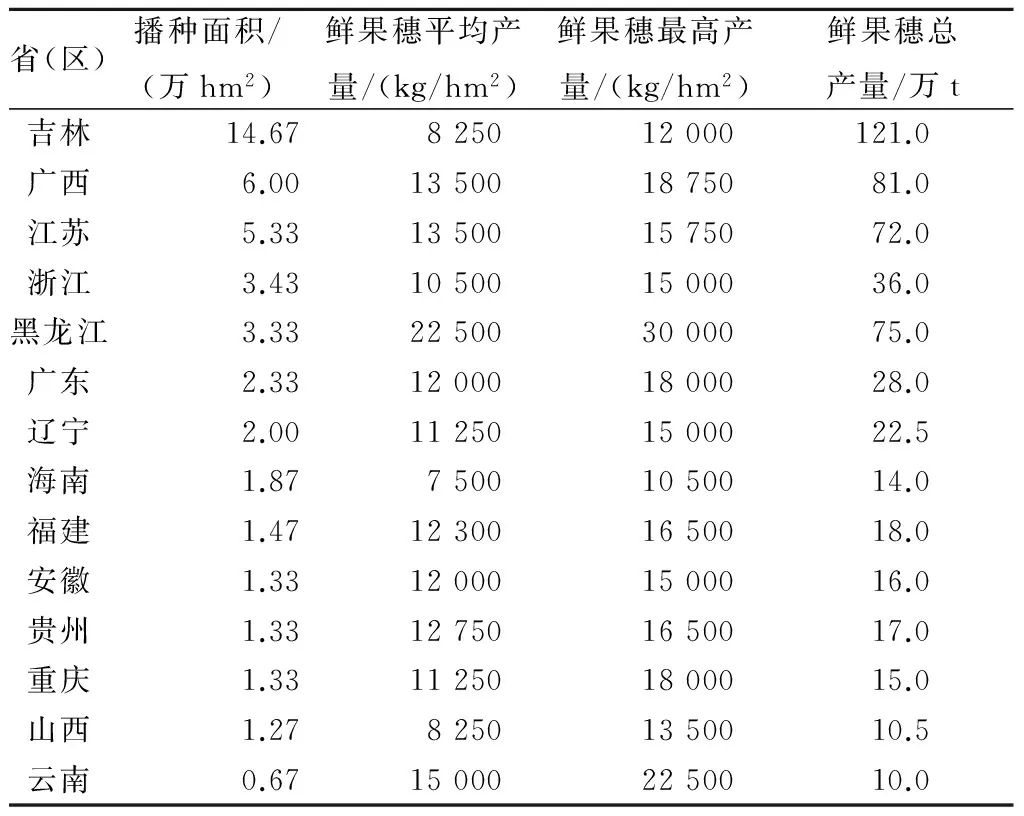
表1 2008年部分省(区)种植的糯玉米面积及产量
注:摘自俞琦英“浙江省甜、糯玉米品种管理现状与对策研究”。
从表1看出,无论种植规模还是产量,云南省的糯玉米生产远低于广东、广西、江浙及山西等省(区)。究其原因可能主要有以下3点:
① 种质资源丰富,但种质利用与创新研究重视力度不够,可用于生产推广的新品种少。同为中国西部的山西省,虽然糯玉米种质资源的数量、丰富性和多样性不及云南,但由于对糯玉米种质资源开发利用的重视,该省从种质资源收集评价[37]、特色资源的遗传学研究[38]、自交系选育与多样性分析[39]等基础研究开始,到新品种选育[40]及配套栽培技术研究[41]、产品贮藏与保鲜[42-43]等形成了糯玉米产前、产中及产后全产业链发展格局,带动了省内几十家企业共同从事糯玉米或鲜食玉米种子生产、种植、果穗贮藏保鲜以及淀粉的生产与销售[44]。
②糯玉米产品单一,层次低,经营粗放。糯玉米虽然用途广泛,但云南生产种植的糯玉米,多数都是以供应本省大中集市鲜嫩果穗为主,少量销往省外的产品也是鲜果穗,缺乏加工糯玉米罐装产品、淀粉产品等深加工的龙头企业引领。相比之下,广西种植的糯玉米只有1/4的面积是用于销售鲜果穗的,而用于深加工的面积达到了3/4[36]。
③种植分散、规模小、农户缺乏规范种植的技术与培训、比较效益低。在云南的种植业中,历来以粮食生产为基础,以烟草生产为主要经济收入来源,其他经济作物如甘蔗、茶、桑及果树等都是配角。因此,在云南,糯玉米既不是主要的粮食作物也不是重要的经济作物,甚至是可有可无的作物,缺少政策引导和种植规划。所以,糯玉米的种植规模小且散,种植户也缺乏规范种植无公害、绿色或有机生态糯玉米的技术与指导,糯玉米较普通玉米更有利用价值的比较经济效益无从体现。但是,随着新一轮产业结构的调整,危害大众健康安全的烤烟生产必将受到控制。对于政府和广大的农户而言,在这种情况下如何保障家庭经济收入不减少、农村经济发展不受影响,将是双方或多方都需要认真研究的社会问题。
因此,建议由政府牵头,组织有关科研、种植农户或者专业合作社、农产品加工企业等,借鉴省外发展糯玉米的先进经验,采取公司+农户+技术保障的发展模式,充分利用云南的光、热资源及山地条件,大力发展糯玉米等特色农产品,带动广大种植户迈向小康生活水平。
[1] 刘帆,谭静,番兴明.云南省优质专用玉米研究现状及发展前景[J].云南农业科技,2011(4):7-11.
[2] 田松杰,石云素,宋燕春,等.利用AFLP技术研究玉米及其野生近缘种的遗传关系[J].作物学报,2004,30(4):354-359.
[3] 张伟.浅谈糯玉米的加工利用[J].现代化农业,2014,(1):22-23.
[4] 李祥艳,唐海涛,张彪,等.我国鲜食甜糯玉米产业现状及前景分析[J].农业与技术,2014,34(4):219-221.
[5] 田孟良,黄玉碧,谭功燮,等.西南糯玉米地方品种waxy基因序列多态性分析[J].作物学报,2008,34(5):729-736.
[6] 曾孟潜,杨太兴,王璞.勐海四路糯玉米品种的亲缘分析[J].遗传学报,1981,8(1):91-96.
[7] 田孟良.中国糯玉米的分子系统学研究[D].雅安:四川农业大学,2005.
[8] 伍少云,孙荣,奉有壁.云南省地方玉米种质资源的类型、品种及其子粒颜色多样性分析[J].玉米科学,2004,12(S1):14-16.
[9] 伍少云,孙荣,奉有壁.云南省玉米地方种质资源类型及其品种的地理和生态分布[J].西南农业学报,2004,17(S1):1-6.
[10] 张建华,陈勇,孙荣,等.云南糯玉米资源的生态类型及其分布[J].云南农业科技,1995(1):41-43.
[11] 张建华,米艳华,张金渝,等.云南糯玉米资源的多样性及其利用[J].西南农业学报,2004,17(6):712-716.
[12] 吴渝生,郑用琏,孙荣,等.基于SSR标记的云南糯玉米、爆裂玉米地方种质遗传多样性研究[J].作物学报,2004,30(1):36-42.
[13] 黄玉碧,荣廷昭,刘礼超.西南地区糯玉米种质资源的遗传多样性——Ⅰ.农艺性状[J].作物杂志,1998(增刊):81-83.
[14] 张建华,杨晓洪,张金渝,等.云南糯玉米地方品种的籽粒淀粉的研究[J].西南农业学报,2006,19(4):543-547.
[15] 李继耕,杨太兴,曾孟潜.栽培玉米起源的同工酶研究[J].遗传学报,1980,7(3):223-230.
[16] 杨太兴,曾孟潜,王璞.我国南方糯玉米(ZeamaysSinensis)的过氧化物酶同工酶分析[J].植物学报,1981,23(2):110-115.
[17] 黄玉碧,荣廷昭.我国糯玉米种质资源的遗传多样性和起源进化[J].作物杂志,1998(增刊):77-80.
[18] 田孟良.云南、贵州糯玉米与普通玉米种质资源的遗传差异比较研究[D].雅安:四川农业大学,2000.
[19] 杨太兴.我国四川糯玉米(Zeamayssinensis)的过氧化物酶同工酶分析[J].科学通报,1988(4):294-296.
[20] 田孟良,黄玉碧,刘永建,等.SSR标记揭示的云南省、贵州省糯玉米与普通玉米种质资源的遗传差异[J].四川农业大学学报,2003,21(3):213-216.
[21] 孙建英.我国的糯玉米[J].种子世界,1988(11):13-14.
[22] 刘永建.用SSR标记研究西南糯玉米种质资源的遗传多样性[D].雅安:四川农业大学,2002.
[23] 张金渝,张建华,杨晓洪,等.用SSR标记划分云南糯玉米地方品种资源遗传类群的研究[J].玉米科学,2007,15(1):53-58.
[24] 张金渝,张建华,杨晓洪,等.西双版纳地区糯玉米品种四路糯、小黄糯的遗传多样性分析[J].中国农业科学,2007,40(2):234-243.
[25] 吴斌,李建,柯洁,等.西双版纳糯玉米地方品种遗传多样性分析[J].玉米科学,2013,21(2):17-23.
[26] 赵耀.西南糯玉米地方品种waxy基因突变类型的初探[D].雅安:四川农业大学,2008.
[27] 刘汉梅.玉米waxy基因和基因组的密码子用法研究[D].雅安:四川农业大学,2008.
[28] 王勇斌.西南糯玉米地方品种waxy基因表达研究[D].雅安:四川农业大学,2009.
[29] 姚坚强,鲍坚东,朱金庆,等.中国糯玉米wx基因种质资源遗传多样性[J].作物学报,2013,39(1):43-49.
[30] 曾孟潜.我国糯质玉米的亲缘关系[J].作物品种资源,1987(3):8-10.
[31] 李艳天,杨瑞华.起源于我国的蜡质玉米[J].种子世界,1985(8):34-35.
[32] 刘坚.中国糯玉米多样性中心及胚乳突变型基因内分子标记策略研究[D].雅安:四川农业大学,2007.
[33] 游修龄.玉米传入中国和亚洲的时间途径及其起源问题[J].古今农业,1989(2):1-10.
[34] 张赞平,李懋学.玉米8个栽培亚种(类型)的核型和C-带带型的比较研究[J].遗传学报,1990,17(2):86-93.
[35] 全丽艳.中国糯玉米蜡质基因位点受到人工选择的分子证据[D].杭州:浙江大学,2007.
[36] 俞琦英.浙江省甜、糯玉米品种管理现状与对策研究[D].杭州:浙江大学,2009.
[37] 张丛卓,白建荣,张效梅,等.山西糯玉米地方资源的表型特性分析[J].山西农业科学,2015,43(2):131-133.
[38] 陈永欣,翟广谦,韩永明,等.黑(果皮)糯玉米主要标志性状及色泽遗传规律研究初报[J].山西农业科学,2009,37(1):19-26.
[39] 程宇坤,白建荣,张效梅,等.山西省糯玉米自交系的遗传多样性分析及类群划分[J].山西农业科学,2012,40(5):433-438
[40] 陈永欣,翟广谦,董立红,等.晋鲜糯玉米系列品种的选育与推广应用[J].山西农业科学,2013,41(10):1037-1041.
[41] 王晓梅,崔坤,迟全勃,等.特用玉米品种筛选与配套栽培技术研究[J].山西农业科学,2011,39(6):552-554,561.
[42] 翟广谦,陈永欣,田福海.速冻保鲜甜、糯玉米营养品质分析[J].山西农业科学,1997,25(3):49-51.
[43] 翟广谦,陈永欣,田福海.糯玉米速冻加工技术研究[J].山西农业科学,1998,26(1):76-79.
[44] 董立红,陈永欣,翟广谦,等.山西省甜糯玉米产业现状与发展对策[J].山西农业科学,2013,41(12):1405-1408.
Research and Utilization of Waxy Maize Germplasm Resources in Yunnan
LEI Yongtao,LONG Wenjie,ZHOU Guoyan,CAI Qing*,WU Shaoyun*
(Biotechnology & Germplasm Resources Institute,Yunnan Academy of Agricultural Sciences/Yunnan Provincial Key Lab of Agricultural Biotechnology/Key Lab of Southwestern Crop Gene Resources and Germplasm Innovation,Ministry of Agriculture,Kunming 650223,China)
The one of center for the origin and genetic diversity of waxy corn is in Yunnan province.In where,the germplasm resources of waxy maize are richer than elsewhere in china,and the type of landraces is obviously diverse,and the distribution of the native varieties is also very wide in 78 counties or cities of 16 states or cities.Silunuo were still planted at Menghai county in Xishuangbanna,which was recognized as waxy corn landraces with a series of primitive traits.This paper reviewed the development of study on waxy corn for the research institutions and researcher in intra-and outer-Yunnan province from ecotype or diversity of landraces,content changes of branched chain starch,bands characteristics of peroxidase isozyme,genetic diversity with molecular markers revealed.The relationship between Menghai Silunuo and Myanmar waxy corn,and Menghai Silunuo and Sikkim original corn were analyzed,and the role and main problems of waxy corn were also analyzed in the developing characteristic agriculture in Yunnan plateau.
waxy maize; germplasm resources; study and utilization; diversity; Yunnan
2015-07-13
科技部、财政部国家科技基础条件平台国家农作物种质资源平台(云南)项目(NICGR2015-030);云南省农业科学院专项(YAAS2015ZY001)
雷涌涛(1967-),女,云南个旧人,本科,主要从事农业生物资源收集保存与利用研究。E-mail:yongtaolei@126.com 隆文杰与第一作者同等贡献。
*通讯作者:蔡 青(1966-),女,云南昆明人,研究员,博士,主要从事农作物种子资源研究。E-mail:caiqingysri@163.com 伍少云(1964-),男,四川马边人,研究员,主要从事农作物资源保存与种质创新工作。 E-mail:caiyunwu205cn@aliyun.com
S513
A
1004-3268(2016)01-0001-07
