体外诱导实现猪的ips细胞向神经干细胞分化
2016-01-16张鼎,张晓姗,王祎
体外诱导实现猪的ips细胞向神经干细胞分化
张鼎,张晓姗,王祎
(中国农业大学生物学院,北京100094)
摘要:项目的主要目标是实现猪的诱导多能干细胞(iPSCs)向神经干细胞(NSCs)体外诱导。在整个实验探索的过程中,我们使用了传统的诱导分化形成神经干细胞(NSCs)的方法,即通过拟胚体(EB)的形成并添加维甲酸(RA)进行诱导。猪的诱导多能干细胞(iPSCs)经历了传代培养、拟胚体(EB)时期、诱导分化时期三个重要阶段,生成一类新的细胞。最后我们使用免疫荧光法检测到新的细胞中存在神经干细胞的特异性标志物 Nestin、神经元标志物β- tubulin III以及神经胶质细胞标志物GFAP表达,从而确定这类新的细胞为神经干细胞。可见,猪的诱导多能干细胞(iPSCs)经过这种传统诱导分化方法培养可分化为具备基础特征的神经干细胞(NSCs),能够表达出神经干细胞(NSCs)特异的标志分子nestin。
关键词:猪;诱导性多能干细胞;神经干细胞;体外诱导
文章编号:1672-6758(2015)06-0059-5
中图分类号:Q28
2006年, Takahashi等将四种转录因子Oct4、Sox2、c-Myc和Klf4导入已分化的小鼠皮肤成纤维细胞,获得了类似于胚胎干细胞(ESCs)的多能性干细胞, 命名为“诱导多能干细胞”(induced pluripotent stem cells,iPSCs)。[1]小鼠和人[2]的诱导多能干细胞系(iPSCs)最先被建立起来,不同种类的畜牧类动物的诱导多能干细胞系(iPSCs)也相继被建立起来,如猪[3]、绵羊[4]。诱导多能干细胞(iPSCs)作为一种新型的多能干细胞,俨然已经成为干细胞界的宠儿,其开发的潜力也是无穷的。随着研究的不断深入,人们在对比诱导多能干细胞(iPSCs)和胚胎干细胞(ESCs)的过程中,发现了诱导多能干细胞(iPSCs)拥有很大的优势。一方面,诱导多能干细胞(iPSCs)由受体自身的细胞重新编码生成,不会诱发免疫反应。例如,在神经系统疾病治疗领域,结合不同的病情,使用诱导多能干细胞(iPSCs)诱导分化成不同类型的神经细胞,能够有效地避免免疫排斥,为临床治疗打下了基础。[5]另外一方面,诱导多能干细胞(iPSCs)能够有效地避免伦理学带来的问题。虽然诱导多能干细胞(iPSCs)依然存在着很多不稳定的因素以及争论,但是不可否认诱导多能干细胞(iPSCs)为再生医学提供了一个全新的方向。[6]
诱导多能干细胞(iPSCs)的研究在小鼠和人的身上都得到了很大的进步,但是目前体内试验依旧局限于小鼠的诱导多能干细胞(iPSCs)上。如果小鼠模型得到的实验结果直接应用到人的身上,是要承担很大的风险的。小鼠和人在生理结构和免疫特性上区别很大,主要体现在两个方面:1.干细胞系多能性维持的主要信号通路不同。经过研究发现,小鼠诱导多能干细胞(iPSCs)多能性维持因子是白血病抑制因子(leukemia inhibitor factor,LIF)、信号传导及转录激活因子(Signal transducers and activators of transcription, STAT),即LIF/STAT3信号通路。[7]人的诱导多能干细胞(iPSCs)多能性维持依赖于碱性成纤维生长因子(basic fibroblast growth factor,bFGF)和激活素(Activin)/节点信号[6]。2.小鼠生命周期过短,是一种很难实现长时间追踪的医学模型。所以我们需要另外一种模式生物,实现小鼠和人之间的过渡。猪作为传统的医学模型,在生理学和解剖学与人类具有很高的相似性。[8]同时经过研究发现,猪和人的诱导多能干细胞系(iPSCs)在维持多能性方面的信号通路具有相似性。[9]而且猪和小鼠的生命周期相比较长,容易进行长时间的追踪。
因为猪的诱导多能干细胞(iPSCs)相对小鼠和人的诱导多能干细胞(iPSCs)是一种新兴的诱导多能干细胞(iPSCs),细胞系的建立自身也不是十分完善,很多地方存在缺陷。由于缺少相关文献的支持,我们只能依照着胚胎干细胞(ESCs)定向诱导分化成为神经干细胞的传统方法,即通过拟胚体的形成并添加维甲酸(RA)进行诱导。
1试剂与材料
(1)ES培养基的配制: KnockoutTM DMEM 培养基(Gibco), ES FBS(15%),Glutamin(100x),NEAA(100x),β-mer(1000x),Lif(1000x);
(2)ES培养基的配制: DMEM/F12(1︰1) 培养基(Gibco), ES FBS(15%),Glutamin(100x),NEAA(100x),β-mer(1000x);
(3)神经样细胞分化培养基的配置: DMEM/F12(1︰1)培养基(Gibco), ES FBS(15%),Glutamin(100x),IBMX(1M),β-mer(1000x),RA(4mM);
(4)抗体:
一抗:mouse anti-Synaptophysin (BM0125) abbit anti-nestin(BA0275) 、rabbit anti-GABA(BA0601) 、rabbit anti-Serotonin (BA0121) ; Mouse anti-GFAP (Sigma, G3893), mouse anti-β-tubulin isotypeⅢ (Sigma, T8660), mouse anti-O4(MAB345);
二抗:标记山羊抗兔IgG(ZF0316), FITC 标记山羊抗鼠IgG(ZF0312)。
2方法
2.1 小鼠的胚胎干细胞、猪的ips细胞的传代培养。
得到小鼠胚胎干细胞系以及猪的ips细胞系, 接种在饲养层细胞上。饲养层细胞经γ射线照射灭活有丝分裂活性,接种ES培养基上培养, 每天更换培养液。
2.2 拟胚体(EB)培养。
在ES培养皿中加入适量的Tryple(酶),37℃消化5min,用移液枪吹打细胞,把细胞悬浮起来。将细胞吸入离心管中,1000r离心5min后,再倒掉上清液,用1-2mlEB培养基把细胞吹起来。再将细胞加到培养皿中,用分化培养液重悬细胞。在培养皿中加入PBS,将每滴悬液滴至平皿上盖表面,迅速扣在下盖上,放入培养箱。培养4 d,隔天换液。
2.3 向神经干细胞的诱导分化。
收集悬浮培养4 d的EB拟胚体,用已经配好的神经样细胞分化培养基进行培养4 d,每2 d换液。
2.4 神经干细胞鉴定。
(1)取出培养皿,抛弃培养体系,将培养液吸去,用PBS洗2次;
(2)固定:用4% PPA处理细胞,在摇床上晃动30 min,接着用PBS洗3次, 5 min/次;
(3)对核表达的蛋白:0.2% Triton-100于培养皿,在室温下30 min,接着用PBS洗3次,5 min/次;
(4)加入3% BSA封闭,室温1 h;
(5)3% BSA稀释的一抗覆盖所染部分,室温1-2h。再用PBS洗上3次,5 min/次;
(6)3%BSA(1:500)稀释的二抗覆盖所染部分,室温1h。弃二抗,PBS洗3次,5 min/次;
(7)加入 1 μg/mL DAPI(1:10000)室温下2 min。再用PBS洗上3次,5 min/次;
(8)封片:在盖玻片上加防淬灭剂,避光保存;
(9)利用荧光显微镜观察并拍照,检测Nestin、GFAP、tublin的表达。(免疫细胞化学荧光染色法)
3结果
3.1 猪的iPSCs以及小鼠的ESCs形态变化过程。
小鼠的ESCs和猪的iPSCs的诱导分化形成NSCs的过程中,形态上的变化是十分明显的。从表观上来看,小鼠的ES细胞和猪的iPS细胞形态上的变化基本是一致的。按照3个阶段进行分析:
3.1.1 传代培养。
复苏后第1天,拨开饲养层细胞,可以观察到细胞小、亮、圆,细胞核大, 核仁明显,细胞核/质比高。细胞排列致密, 边界清晰,呈集落形状生长。
复苏后第3天, 细胞克隆进一步增大, 中央逐渐突起, 显微镜下观察立体感更强, 细胞排列更紧密。

(a)生长旺盛的MES细胞(×40)

(b)生长旺盛的Pips细胞(×40)
3.1.2 拟胚体(EB)的形成。
去除饲养层细胞后,进行悬浮培养,细胞能够逐步地形成拟胚体(EB)。拟胚体(EB)呈球形,胚体饱满。内细胞间黏附紧密,边界清楚。

(a)MES的拟胚体 (×40)
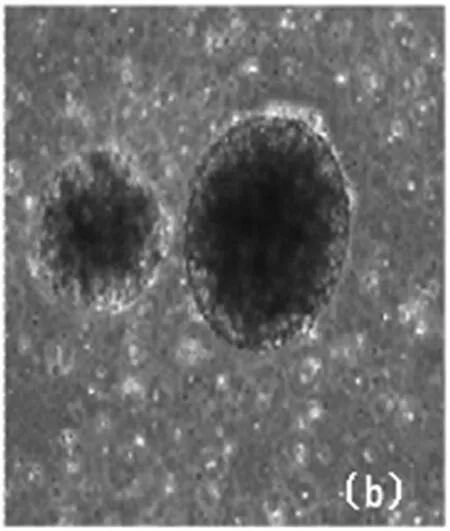
(b)Pips的拟胚体 (×40)
3.1.3 诱导分化。
在加入了维甲酸(NA)后,ES细胞获得了向某一特定方向诱导分化的趋势。
加入了诱导剂后第1天细胞的形态发生改变,有产生区域化模式的趋势,但不是十分清晰。
加入了诱导剂后3天,细胞的形态发生很大的改变,同时有细胞出现死亡现象。区域化模式十分明显,很清晰地看到了同一类型的细胞逐步靠拢。
3.2 猪iPS细胞以及小鼠ES细胞来源的神经样细胞的相关标志物的鉴定。
免疫荧光结果显示诱导7d后的神经球样克隆表达NSCs标志物nestin,贴壁培养后的神经球样克隆表达神经元标志物β-tubulin III和星形胶质细胞标志物GFAP。

小鼠:(a)单层神经干细胞nestin阳性 (×100)

小鼠:(b)β-tubulin III阳性(×100)

小鼠:(c)GFAP阳性(×100)
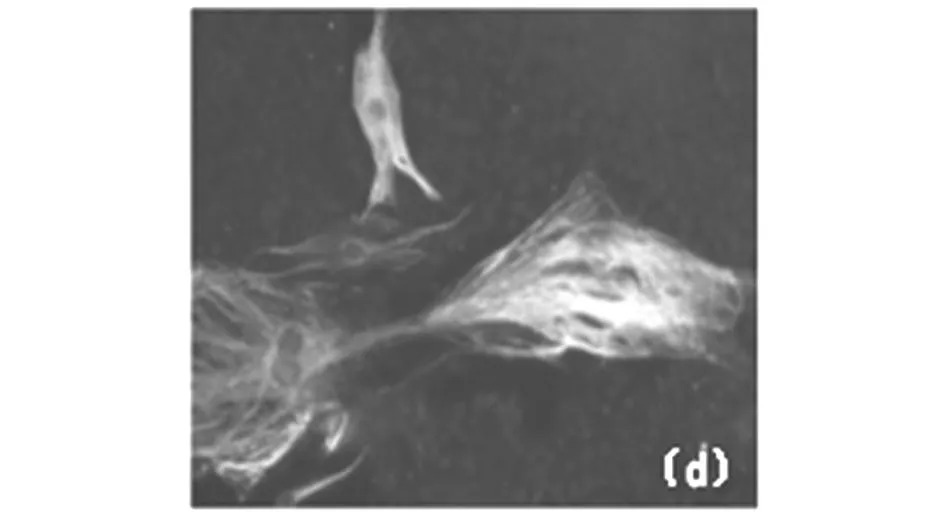
猪:(d)单层神经干细胞nestin阳性 (×100)

猪:(e)β-tubulin III阳性(×100)

猪:(f)GFAP阳性(×100)
4讨论
诱导多能干细胞(iPSCs)是一种新型的多能干细胞,具有类似于胚胎干细胞(ESCs)的特性。所以本实验借鉴了胚胎干细胞(ESCs)向神经干细胞(NSCs)诱导分化的方法。目前胚胎干细胞(ESCs)向神经干细胞(NSCs)诱导分化方法主要有以下几种: 维甲酸(RA)诱导, PA6基质细胞诱导,生长因子诱导,直接分化法。为何选择维甲酸(RA)进行诱导时基于3个方面进行考虑:
(1)维甲酸(RA)作为一种传统的分化诱导剂,它可诱导类胚体分化成多种细胞类型,其分化方向以及程度取决于维甲酸的剂量及作用阶段。[10]
(2)生长因子诱导、PA6基质细胞诱导这两种方法涉及的诱导成分过于复杂,而且对于培养的要求也是极高的。
(3)直接分化法操作简单,效率很高,是现在主流的分化方法。但是,缺少了拟胚体(EB)向神经干细胞(NSCs)过渡这一步,不利于结果分析。[11]
实验的过程中,猪的诱导多能干细胞(iPSCs)在经历传代培养、拟胚体(EB)的形成、维甲酸(RA)诱导分化以后,得到了悬浮培养的神经球。神经球能够表达NSCs标志物, 这说明猪的诱导多能干细胞(iPSCs)已经分化为NSCs。但是,对比小鼠的胚胎干细胞(ESCs)以及猪的诱导多能干细胞(iPSCs)在分化过程中的区别还是十分明显的。分化的第3d,我们在小鼠神经样细胞分化培养基中发现贴壁生长的拟胚体(EB)中央出现特有的环状空腔结构,我们推测是神经祖细胞特有的环状rosette空腔结构,一种类似于早期的神经管结构。[12]但是,猪的胚胎干细胞(ESCs)却没有出现这种rosette空腔结构。
虽然我们最终得到了神经干细胞(NSCs),但是我们并没有检测分化最终的效率,这是实验的一个遗憾之处。如果要进行更加深入的实验,我们有必要对于这种分化方法的效率进行探索。现在主流的检测分化效率的方法:RNA的分离和定量PCR分析——首先使用RNeasy Mini Kit这种提取试剂盒去实现对这个体系中所有细胞RNA的提取,再利用SuperScript III第一链合成系统进行逆转录得到细胞的互补DNA(cDNA)。在完成这些准备工作后,我们再利用SYBR Green mix对cDNA进行染色,最终可以利用Roche LightCycler 5480等仪器进行数据分析。[13]
经过以上的研究表明,通过诱导分化传统的方法即通过拟胚体的形成并添加RA进行诱导,的确可以使得猪的诱导多能干细胞(iPSCs)形成神经干细胞(NSCs)。但是,这得到的仅仅是初步的结果,至于得到的神经干细胞(NSCs)是否具有终末分化能力、是否有成瘤性、能否发挥相应的功能等, 都需要进一步深入研究。
5展望
随着iPS细胞技术的发展,人们逐步建立起了家畜类动物的iPS细胞系,如猪、绵羊。对于家畜类动物而言,iPS细胞技术的出现为这些很难建立ES细胞系的物种建立多能干细胞系带来了希望。因为家畜类动物ES细胞系长期培养后易出现分化现象,所以建立的ES细胞系难度大。如果成功建立家畜iPS细胞系,利用其独特的优势,将iPS细胞诱导技术与相关技术相结合,如嵌合体技术,在家畜动物的异种器官移植、品种改良以及人源性疾病动物模型的制作[14]等方面都具有广阔的开发前景。
虽然部分家畜类动物的iPS细胞系得到了建立,但是依旧存在很多不足。由于iPS细胞是通过体外重新编码形成的,即使猪作为最早建立iPS细胞系的家畜类动物,依旧很难确定重新编码的基因是否继续表达从而能使得细胞定向分化的过程变得复杂化。比如猪ID6细胞系[15]已被用于产生视网膜前体细胞,可以用来恢复视网膜的能力。[16]但是通过使用诱导人类ES细胞系的方法,在驱动猪ID6细胞系定向分化的过程中很难出现内胚层或中胚层细胞。这与各大实验室得到的结论:由iPSC形成的拟胚体中必定存在这样的结构,相违背。[17]
实验表明,通过在神经退行性疾病动物模型中移植神经干细胞(NSCs),神经干细胞(NSCs)可以通过分泌神经营养因子(Neurotrophin,NT),如神经生长因子(Nerve Growth Factor, NGF)、脑源神经营养因子(brain derived neurotrophic factor, BDNF)等,保护神经元并引起神经再生效应。[18]Parkinson可见神经干细胞(NSCs)在临床治疗方面具有很大的开发价值。随着iPS细胞技术的发展,诱导多能干细胞(iPSCs)定向诱导分化成神经干细胞(NSCs)必将成为神经干细胞(NSCs)的主要来源。
2014年,一名罹患退行性眼病的日本患者成为全球使用诱导多能干细胞(iPSCs)进行治疗的第一人,[19]这意味着通过iPS技术所代表的克隆技术治疗人类疾病的设想将首次付诸实践。虽然还需要后续的研究,才能确定这一实践的最终效果,但是不可否认这是一步很大的跨越,为干细胞疗法的临床实践打开了一个新的篇章。由于iPS细胞系的建立方式以及遗传稳定性依然存在着很多不确定性的因素,iPS细胞技术还需要更深入研究和探索。
参考文献
[1]Takahashi, K.;Yamanaka, S., Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors.cell2006, 126 (4), 663-676.
[2]Yu, J.;Vodyanik, M. A.;Smuga-Otto, K.;Antosiewicz-Bourget, J.;Frane, J. L.;Tian, S.;Nie, J.;Jonsdottir, G. A.;Ruotti, V.;Stewart, R., Induced pluripotent stem cell lines derived from human somatic cells.Science2007, 318 (5858), 1917-1920.
[3]Esteban, M. A.;Xu, J.;Yang, J.;Peng, M.;Qin, D.;Li, W.;Jiang, Z.;Chen, J.;Deng, K.;Zhong, M., Generation of induced pluripotent stem cell lines from Tibetan miniature pig.JournalofBiologicalChemistry2009, 284 (26), 17634-17640.
[4]Bao, L.;He, L.;Chen, J.;Wu, Z.;Liao, J.;Rao, L.;Ren, J.;Li, H.;Zhu, H.;Qian, L., Reprogramming of ovine adult fibroblasts to pluripotency via drug-inducible expression of defined factors.Cellresearch2011, 21 (4), 600-608.
[5]Uemura, T.;Takamatsu, K.;Ikeda, M.;Okada, M.;Kazuki, K.;Ikada, Y.;Nakamura, H., Transplantation of induced pluripotent stem cell-derived neurospheres for peripheral nerve repair.Biochemicalandbiophysicalresearchcommunications2012, 419 (1), 130-135.
[6]Takahashi, K.;Tanabe, K.;Ohnuki, M.;Narita, M.;Ichisaka, T.;Tomoda, K.;Yamanaka, S., Induction of pluripotent stem cells from adult human fibroblasts by defined factors.cell2007, 131 (5), 861-872.
[7]Hall, J.;Guo, G.;Wray, J.;Eyres, I.;Nichols, J.;Grotewold, L.;Morfopoulou, S.;Humphreys, P.;Mansfield, W.;Walker, R., Oct4 and LIF/Stat3 additively induce Krüppel factors to sustain embryonic stem cell self-renewal.Cellstemcell2009, 5 (6), 597-609.
[8]Brandl, U.;Michel, S.;Erhardt, M.;Brenner, P.;Burdorf, L.;J ckle, H.;Bittmann, I.;R ssle, M.;Mordstein, V.;Baschnegger, H. InTransgenicanimalsinexperimentalxenotransplantationmodels:orthotopichearttransplantationinthepig-to-baboonmodel, Transplantation proceedings, Elsevier: 2007;pp 577-578.
[9]Alberio, R.;Croxall, N.;Allegrucci, C., Pig epiblast stem cells depend on activin/nodal signaling for pluripotency and self-renewal.Stemcellsanddevelopment2010, 19 (10), 1627-1636.
[10]Rohwedel, J.;Guan, K.;Wobus, A. M., Induction of cellular differentiation by retinoic acid in vitro.Cells,tissues,organs1999, 165 (3-4), 190-202.
[11]Kawasaki, H.;Mizuseki, K.;Nishikawa, S.;Kaneko, S.;Kuwana, Y.;Nakanishi, S.;Nishikawa, S.-I.;Sasai, Y., Induction of midbrain dopaminergic neurons from ES cells by stromal cell-derived inducing activity.Neuron2000, 28 (1), 31-40.
[12]Wang, J.;Jiao, F.;Pan, X.-h.;Xie, S.-y.;Li, Z.-l.;Niu, X.-h.;Du, L.-x., Directed differentiation of chick embryonic germ cells into neural cells using retinoic acid induction in vitro.Journalofneurosciencemethods2009, 177 (1), 168-176.
[13]Choi, H. W.;Kim, J. S.;Choi, S.;Hong, Y. J.;Kim, M. J.;Seo, H. G.;Do, J. T., Neural stem cells differentiated from iPS cells spontaneously regain pluripotency.StemCells2014, 32 (10), 2596-2604.
[14]Fujimura, T.;Takahagi, Y.;Shigehisa, T.;Nagashima, H.;Miyagawa, S.;Shirakura, R.;Murakami, H., Production of alpha 1, 3‐Galactosyltransferase gene‐deficient pigs by somatic cell nuclear transfer: A novel selection method for gal alpha 1, 3‐Gal antigen‐deficient cells.Molecularreproductionanddevelopment2008, 75 (9), 1372-1378.
[15]Roberts, R. M.;Telugu, B. P. V.;Ezashi, T., Induced pluripotent stem cells from swine (Sus scrofa): why they may prove to be important.CellCycle2009, 8 (19), 3078-3081.
[16]Zhou, L.;Wang, W.;Liu, Y.;de Castro, J. F.;Ezashi, T.;Telugu, B. P. V.;Roberts, R. M.;Kaplan, H. J.;Dean, D. C., Differentiation of induced pluripotent stem cells of swine into rod photoreceptors and their integration into the retina.StemCells2011, 29 (6), 972-980.
[17]Ezashi, T., B. Telugu, and R. Roberts, Induced pluripotent stem cells from pigs and other ungulate species: an alternative to embryonic stem cells Reproduction in Domestic Animals, 2012. 47(s4): p. 92-97.
[18]Zhou, C.;Wen, Z.-X.;Wang, Z.-P.;Guo, X.;Shi, D.-M.;Zuo, H.-C.;Xie, Z.-P., Green fluorescent protein-labeled mapping of neural stem cells migrating towards damaged areas in the adult central nervous system.Cellbiologyinternational2003, 27 (11), 943-945.
[19]Kawamata, S.;Kanemura, H.;Sakai, N.;Takahashi, M.;Go, M. J., Design of a Tumorigenicity Test for Induced Pluripotent Stem Cell (iPSC)-Derived Cell Products.JournalofClinicalMedicine2015, 4 (1), 159-171.
Inducing Porcine iPS Cells into Neural Stem Cellsinvitro
Zhang Ding,Zhang Xiaoshan,Wang Yi
(School of Biological Sciences, China Agriculture University, Beijing 10094, China)
Abstract:The major objective of our study is to induce porcine iPS cells into neural stem cells in vitro. During the whole process of experimental exploration, we induced porcine iPS cells into neural stem cells by using a traditional method: inducing embryoid body(EB) into neural stem cells by adding Retinoic acid(RA) in vitro. Porcine iPSCs underwnet subculture、embryoid body(EB)、inducing differentiation three important stages to form another kind of cells.Finally,we validated the newly formed cell is neural stem cells by using immunofluorescence assay to detect the neural stem cell specific marker Nestin、the neuronal marker β- tubulin III and glial cell marker GFAP in the new formed cell. As a result, porcine iPSCs induced by this traditional method can successfully induced into neural stem cells and express nestin.
Key words:pig;iPS cells;neural stem cells(NSCs);induceinvitro
Class No.:Q28Document Mark:A
(责任编辑:宋瑞斌)
