盐胁迫对海滨木槿幼苗生长及光合特性的影响
2015-12-30李贺鹏张晓勉岳春雷高智慧
卢 刚,李贺鹏*,张晓勉,陈 燕,岳春雷,高智慧
(1. 浙江省林业科学研究院,浙江 杭州 310023;2. 浙江长征职业技术学院,浙江 杭州 310023;3. 浙江省林业技术推广总站,浙江 杭州 310020)
盐胁迫对海滨木槿幼苗生长及光合特性的影响
卢 刚1,李贺鹏1*,张晓勉1,陈 燕2,岳春雷1,高智慧3
(1. 浙江省林业科学研究院,浙江 杭州 310023;2. 浙江长征职业技术学院,浙江 杭州 310023;3. 浙江省林业技术推广总站,浙江 杭州 310020)
通过人工模拟的方法研究了NaCl胁迫(0、0.3%、0.6%、0.9%)对海滨木槿幼苗生长和光合特性的影响。结果表明:除了0.9%处理组1株第44 d死亡,盐胁迫对其他幼苗的生长影响不大;海滨木槿的光合指标随盐胁迫浓度、胁迫时间的变化而动态变化,至30 d,所有处理的叶绿素含量(Chl)恢复至正常水平,0.3%处理组的各指标中除了Rd和LCP外均恢复至正常水平,0.6%和0.9%处理组的各指标也得到不同程度的恢复;胁迫40 d,除了0.6%处理组的Rd和LCP显著高于对照,以及0.9%处理组的Pnmax显著低于对照(p < 0.05)外,各指标基本恢复至对照水平;研究表明海滨木槿不仅对盐胁迫具有一定的耐性,且对胁迫伤害具有较强的自我恢复能力。
海滨木槿;盐胁迫;光合特性;耐盐;生理特性;光响应
土壤盐渍化是世界性的资源环境和生态问题,由于人类不断开发和不合理利用土地,致使盐渍土和土壤次生盐渍化在世界各地不断加重,已成为制约人类社会发展和进步的全球性难题 。目前全球约有 8×10 km的盐渍化土地[3],大约20%的可耕地以及约50%的灌溉地不同程度地受到盐分的影响[4]。目前我国拥有现代盐渍土、残余盐渍土和潜在盐渍土约9.9×108hm2,主要分布在沿海及干旱和半干旱地区[5]。土壤盐分是限制植物生长和生产的主要环境因子之一[6],盐胁迫对植物造成的伤害是多方面的,包括渗透胁迫、离子毒害和细胞内离子平衡的破坏等,使植物生长受到抑制,光合速率下降,衰老加速,植株最终死亡等[7~8]。光合作用作为绿色植物最重要的生理代谢过程之一,光合作用对逆境胁迫伤害最为敏感[9],因此研究盐胁迫对光合作用的影响,对于阐明植物的盐害机理、提高植物耐盐性、改良盐渍地等有重要意义。
随着我国医疗技术水平的提高及医疗保障制度的日益完善,临床用血量每年呈现上升趋势,血液供不应求的矛盾日益严重[1],我院也不例外的出现临床供血短缺常态化。它除直接影响失血性休克患者的抢救外,还使一些择期手术患者手术延期,贫血患者的输血治疗滞后,使患者的生命健康受到严重威胁[2],直接影响了正常的医疗活动和医疗秩序,增加了医患纠纷发生的风险。我院对后者,大多通过自助献血缓解。但2018年4月1日起,国家卫生计生委全面叫停了自助献血。那如何化解此现状,成了相关各方的当务之急。下文就可能造成临床供血短缺原因和其破解途径探讨如下:
海滨木槿(Hibiscus hamabo)是锦葵科(Malvaceae)木槿属(Hibiscus)落叶灌木或小乔木,在20世纪80年代初被鉴定命名[10],原产在我国浙江定海、镇海,舟山岛西部及长山寺等几个小岛,奉化缸山岛等亦有少量分布;江苏北部,福建云霄县,广东陆丰及徐闻的海滨,日本、朝鲜也有分布[11~12]。海滨木槿根系发达,对土壤的适应能力和抗风力强,能耐短时期的水涝,抗干旱和寒冷,能抵御冬季-10℃的低温,是华东沿海地区优良的沿海防护林树种。目前,关于海滨木槿的研究主要集中在种质资源分布情况及引种驯化[12]、种子种苗繁育研究[13~19],以及盐胁迫、水淹胁迫下生理生化特性方面的研究[20~27],另外对潮间带海滨木槿的光合特性研究也有相关报道[28~29],但关于NaCl胁迫条件下海滨木槿的光合特性和生理特性未见有系统研究报道。
2.4.2 药剂防治该病除选用上述技术原则外,在发病初期常用药剂有:50%多菌灵可湿性粉剂用种子量0.3%拌种或500~800倍液喷雾,或用70%甲基硫菌灵可湿性粉剂500倍液~800倍液。
本试验通过对海滨木槿幼苗进行不同浓度NaCl处理,研究不同盐胁迫阶段下幼苗生长、光合特性及相关生理的变化,探讨海滨木槿叶片光合特性对长期盐胁迫响应特征,为充分利用滩涂、盐碱地发展种植提供科学依据。
1 材料与方法
1.1 试验材料
试验材料为2年生实生苗。于2012年6月上旬进行盆栽,每盆栽植1株。花盆选用规格为:上口直径30.5 cm,下口直径20.0 cm,高24.0 cm。栽培基质为苗圃熟土,其理化特性分别为全氮0.677 g/kg、全磷0.448 g/kg、速氮65.962 mg/kg、速磷7.547 mg/kg、速钾200.755 mg/kg、有机质17.430 g/kg。正常浇水培育,并进行适当管护。经过1个多月的室外炼苗,于7月中旬移入温室大棚,选取长势一致的苗木进行NaCl胁迫处理。试验设置1个对照和3个强度的胁迫处理,即对照组(浇水)(ck)、0.3%(低盐)、0.6%(中盐)和0.9%(高盐),每组试验各设置3~4个重复。
式中:Pn为净光合速率(μmol·m-2·s-1);AQY为表观量子效率(μmol·μmo1-1);PAR为光合有效辐射(μmol·m-2·s-1);Pnmax为最大净光合速率(μmol·m-2·s-1);k为曲线的曲角;Rd为暗呼吸速率(μmol·m-2·s-1)。利用在0 ~ 200 μmol·m-2·s-1的Pn对PAR进行线性回归,直线在X轴上的截距为光补偿点(LCP,μmol·m-2·s-1);当Pn= Pnmax时,PAR的值即为光合作用光饱和点(LSP,μmol·m-2·s-1)[32]。
高碳天然气捕集技术:以松南高碳天然气分离捕集装置为代表,采用MDEA胺法脱碳工艺,将松南高碳气田(二氧化碳含量23%)中的二氧化碳捕集分离,生产清洁天然气,年处理能力达50万吨。
生长痛(growing pain)是很多孩子都经历过的一种情况,常见到什么程度呢?仅从小儿骨科门诊生长痛的就诊率来看,小儿骨科门诊病人数的1/3以上都是因为这种情况。
民勤县:石羊河流域重点治理 助推绿洲生态显生机……………………………………………………… 王江文(5.67)
利用LI-6400便携式光合作用测定系统(Licor,USA)对光响应曲线进行测定。选取自顶端向下的第3、4片叶片,采用标准叶室(2 cm×3 cm)并使用红蓝光源(red blue 6400-02B),设定光合有效辐射为0、20、50、100、200、400、600、800、1 000、1 200、1 500、2 000 μmol·m-2·s-1,测定叶片净光合速率(Pn)、叶片气孔导度(Gs)、胞间CO2浓度(Ci)等指标,气孔限制值(Ls)通过下列公式计算:
Ls= 1-Ci/Ca
式中,Ca为大气CO2浓度。
选用标准荧光叶室(6400-40 Default Flurometer)进行叶绿素荧光参数的测定。首先采用配套的暗适应叶夹对各个处理相同部位功能叶进行30 min的遮光处理,立即测定其初始荧光(Fo)、最大荧光(Fm)、最大光化学效率(Fv/Fm)。选取的叶片与光合—光响应曲线测量的相同。
每隔10 d测定一次,即2012年8月4日、8月14日、8月24日和9月3日,均为晴朗天气。
1.2 光合—光响应曲线和叶绿素荧光参数的测定
1.3 叶绿素含量的测定
In Finding Nemo,we follow(跟随)Marlin as he looks for his son Nemo.On the way,he becomes good friends with Dory.Dory is a friendly and cute fish.But she has a bad memory(记忆).
1.4 数据处理与分析
采用非直角双曲线模型对光响应曲线进行拟合[31]:

随着我国新农村建设不断深入推行,大量自然村采取政治合并的手段,因此村级财务管理制度不够健全,出现了大量漏洞.在许多自然村基本上都有村主任或者是村委书记一把手独掌大权,极大影响了村级财务管理的效果。同时由于农村财务管理人才较为匮乏,因此对于村干部的财务管理行为缺乏有效的监督机制,约束机制不够健全,很容易出现村领导干部滥用职权或者是以权谋私的现象,造成严重突出的职权,监督缺失的问题。
应用SPSS13.0统计分析软件进行统计分析,并采用LSD法进行多重比较和显著性检验;应用Origin7.5进行数据图绘制。
2 结果与分析
2.1 盐分胁迫下海滨木槿幼苗的生长情况
观察发现,盐分胁迫对海滨木槿生长造成一定影响。施盐后7d,0.9%处理组中少量叶片变黄,枝条顶端2、3片叶片变枯、变干;4株幼苗叶片在第10天萎蔫,顶端叶片变枯;第16天,0.6%处理组叶片稍显萎蔫,0.9%处理组叶片萎蔫;第22天,0.6%处理组变化不大,0.9%处理组增加几片黄叶;第27天,0.6%处理组叶片显萎蔫;0.9%处理组少量萎蔫叶片脱落;第32天,0.6%处理组变化不大,0.9%处理组出现部分干枯叶片;第38天,0.6%处理组变化不大,0.9%处理组1株叶片接近全干,其它叶片萎蔫;第44天,0.6%处理幼苗生长正常,0.9%处理组1株死亡,其它变化不大;第47天,0.6%处理幼苗生长正常,0.9%处理组变化不大;第56天后,所有正常生长的植株变化不大。
2.2 盐分胁迫下海滨木槿幼苗叶片总叶绿素含量的响应
不同盐浓度和胁迫时间对海滨木槿幼苗叶片总叶绿素含量产生不同的影响见图1。胁迫初期(10 d),叶片叶绿素含量随着盐浓度的增加而呈递减趋势,方差分析显示处理组都显著低于对照组(p < 0.05),同时0.3%的处理组显著高于0.9%的处理组(p < 0.05),说明盐度胁迫影响叶片叶绿素含量,并且高盐度胁迫的影响更加显著;随着时间的延长(20 d),海滨木槿叶片叶绿素含量与初期相比有一定程度的恢复,0.3%的处理组与对照差异不显著(p > 0.05),但0.6%和0.9%处理组都显著低于对照(p < 0.05);随着处理时间的进一步延长(30 d和40 d),所有处理组的叶绿素含量已恢复至对照水平,并且在第30天还略高于对照组,但不同处理组之间都差异显著(p > 0.05)。这些结果说明海滨木槿能够耐受一定浓度的盐胁迫。
选取与测定光合作用和叶绿素荧光相同位置和发育阶段的叶片,用纯丙酮、无水乙醇和蒸馏水的混合液法提取叶绿素,并用紫外分光光度计测定吸光值,然后根据公式计算得出叶绿素总含量[30]。

图1 盐胁迫对海滨木槿幼苗叶片总叶绿素含量的影响Figure 1 Effect of NaCl stress on total chlorophyll content in leaves of H. hamabo seedling不同字母表示处理间在0.05水平有显著差异,下同
2.3 盐胁迫下海滨木槿幼苗气体交换参数对PAR的响应
2.3.1 Pn植物光合作用光响应曲线反映了光强与植物Pn的关系,不同盐度胁迫影响了海滨木槿的光合速率(图2)。盐度胁迫初期(图2A)海滨木槿叶片的光合效率明显下降,且随着盐度浓度的增加而明显下降,其中0.9%的处理组海滨木槿的光合速率基本在0点波动。在处理第20天(图2B),盐胁迫的抑制作用达到最强,0.3%的处理组稍高于0.6%和0.9%的处理组,而后两组处理的光合速率几乎在0点波动。随着胁迫时间的延长,海滨木槿的Pn得到一定恢复,其中0.3%处理组的Pn几乎恢复到与对照相似的水平(图2C);0.6%的处理组稍高于0.9%的处理组,其Pn最大值超过3.0,而0.9%的处理组Pn最大值也在2.0以上。随着时间的进一步延长(第40天),不同盐度的处理组都得到明显恢复,0.6%的处理组的Pn已经接近0.3%处理组的水平;而0.9%处理组的Pn最大值达到3.0。这些结果说明海滨木槿的光合作用在胁迫条件下受到了影响,但是随着时间的延长,盐度胁迫对植物光合作用的抑制作用呈减弱的趋势,证实了海滨木槿对盐度胁迫有较好的适应性;另外随着盐度浓度的增强,其对植物光合作用的影响越明显。

图2 盐胁迫对海滨木槿幼苗叶片的Pn—PAR响应Figure 2 Effect of NaCl stress on responses of Pnto PAR in H. hamabo seedling leaves
2.6 盐胁迫对海滨木槿幼苗叶片光合光响应曲线参数的影响
低年级语文教材的特点是:内容明显,事理简单,篇幅短小,易读易记。重要的一条则是多读,只有多读,才能品出味道。课堂上,教师一定要重视引导儿童进行读书训练,从读中达到理解、消化、吸收的目的。获得读书的本领——“字求其训”“句求其义”“篇求其旨”。品味语言,克服摒弃过去教师包办代替式的讲读方法。
2.3.3 Ci盐胁迫下不同处理阶段的Ci受到一定影响(图4)。胁迫初期,0.9%的处理组的Ci低于对照组,在PAR低于100 μmol·m-2·s-1时,Ci随着光合有效辐射强度的逐渐升高而呈下降的变化,但在PAR处于200 ~ 1 500 μmol·m-2·s-1时,Ci呈波动的变化(图4A);0.3%和0.6%处理组的Ci随着光合有效辐射强度的增强呈先降低后略有升高的变化,但均低于对照组。胁迫中期,低盐、中盐、高盐3个处理浓度都高于对照组(图4B)。胁迫中后期,除了0.3%处理的Ci高于对照外,其他中、高浓度处理组的Ci均低于对照(图4C)。胁迫后期,3个处理组的Ci基本与对照组有相似的变化趋势,并且Ci均接近对照组(图4D)。这表明盐分胁迫下,海滨木槿叶片的细胞间隙CO2浓度下降,表明经叶片气孔扩散进入叶肉细胞的CO2浓度下降,从而不能满足叶片进行正常光合作用的需要,导致Ci下降;盐胁迫下,细胞间隙CO2浓度有所上升,表明叶肉细胞光合能力进一步减弱,对CO2的利用减少,导致CO2过剩,光合产物相应减少,这可能是盐胁迫下叶肉细胞本身光合能力下降所致。

图3 盐胁迫对海滨木槿幼苗叶片Gs—PAR 的影响Figure 3 Effect of NaCl stress on responses of Gsto PAR in H. hamabo seedling leaves图4 盐胁迫对海滨木槿幼苗叶片Ci—PAR的影响Figure 4 Effect of NaCl stress on responses of Cito PAR in leaves of H. hamabo seedling

图3 盐胁迫对海滨木槿幼苗叶片Gs—PAR 的影响Figure 3 Effect of NaCl stress on responses of Gsto PAR in H. hamabo seedling leaves图4 盐胁迫对海滨木槿幼苗叶片Ci—PAR的影响Figure 4 Effect of NaCl stress on responses of Cito PAR in leaves of H. hamabo seedling
吴玉梅不得已,只得如实告诉他们:说杨力生和杨秋香是已谈过恋爱的。老两口一边仍然表示不同意,一边暗暗埋怨自己儿子眼光低下。
盐胁迫下海滨木槿幼苗叶片Ls随着处理时间和处理浓度的不同而变化(图5)。胁迫初期(图5A),各处理随着PAR的增强,Ls不断增大,PAR达到200 μmol·m-2·s-1左右时,随着PAR的增强,Ls趋于稳定;0.3% 和 0.6%处理组的 Ls基本上高于对照组,而 0.9%处理组则相对低于对照,并且在 PAR处于 200 ~ 2 000 μmol·m-2·s-1范围时,Ls处于波动的变化;胁迫中期(图5B),各处理组的Ls均低于对照,并且随盐胁迫浓度的增加,Ls下降越明显;同时,在PAR处于200 ~ 1 500 μmol·m-2·s-1范围时,0.6%和0.9%处理组的Ls呈波动的变化。胁迫中后期(图5C),除了0.3%处理Ls低于对照外,其他中、高浓度处理组均高于对照。胁迫后期(图5D),随着PAR的变化,其气孔限制值与胞间CO2浓度的变化一致。由此可见,盐胁迫在不同时间阶段对植物Ls影响存在差异,并且随着处理浓度的变化呈差异性的变化。
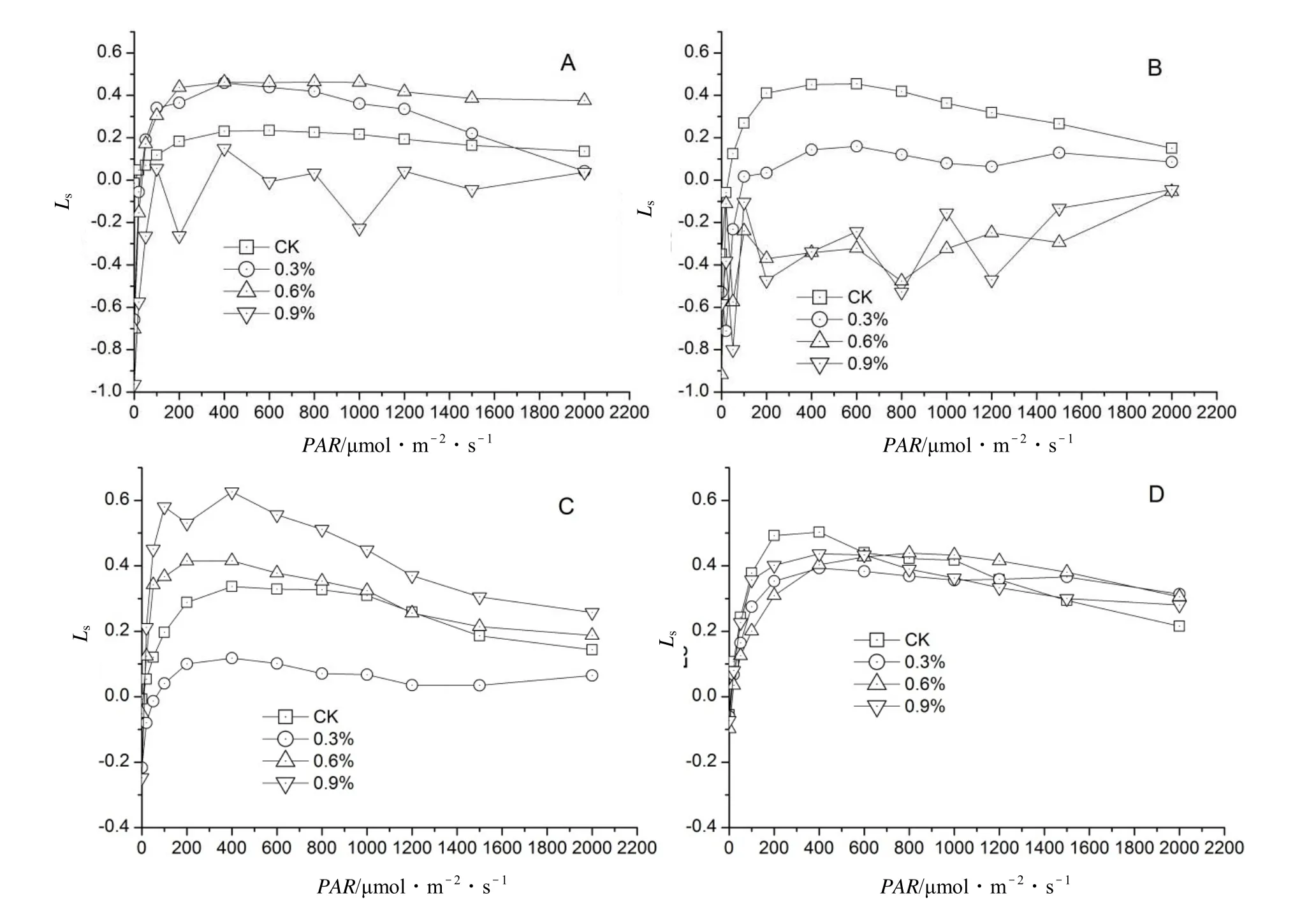
图5 盐胁迫对海滨木槿幼苗叶片Ls—PAR的影响Figure 5 Effect of NaCl stress on responses of Lsto PAR in leaves of H. hamabo seedling
2.5 盐胁迫对海滨木槿幼苗叶片叶绿素荧光的影响
Fv/Fm、Fv/Fo分别反映了光系统Ⅱ(PSⅡ)反应中心原初光能转化效率和PSⅡ潜在的光化学活性,常用来表征植物是否处于被胁迫状态。在试验初期(10 d),处理组的Fv/Fm随着盐度浓度的增加呈逐渐下降的变化趋势(图6),且显著低于对照组(p < 0.05),0.3%的处理组显著高于0.9的处理组(p < 0.05),同时高于0.6%的处理组,但差异不显著(p > 0.05);随着处理时间的延长,Fv/Fm得到明显恢复,除了0.9%的处理组在20 d 和30 d以及0.6%的处理组在30 d时有明显差异(p < 0.05)外,其他已经无显著差异(p > 0.05)。上述结果表明盐胁迫初期对海滨木槿幼苗的光合器官发生严重的光抑制,高度盐度的胁迫更加明显,但随时间的延长,其抑制作用逐渐减弱。
2.4 盐分胁迫下海滨木槿幼苗Ls对PAR的响应
从图7可以看出,在盐度胁迫初期(10 d)处理组的Fv/Fo逐渐降低,都显著低于对照组(p < 0.05),但处理组之间都差异不显著(p > 0.05);随着处理时间的延长(20 d),0.3%和0.6%的处理组已恢复至对照组的水平(p > 0.05),但0.9%处理组显著低于对照组(p < 0.05);30 d的Fv/Fo变化趋势与20 d的相似,其中0.6%处理组还明显低于20 d的处理组(p < 0.05);第40天,各处理组都与对照组的Fv/Fo无显著差异(p > 0.05)。这说明在盐度胁迫下,叶片光能转化效率在初期受到抑制作用,但到试验末期基本得以恢复,叶片光合机构的功能随着时间延长也逐渐显现。

图6 盐胁迫对海滨木槿幼苗叶片PSⅡ最大光化学效率的影响(P < 0.05)Figure 6 Effect of NaCl stress Fv/Fmin leaves of H. hamabo seedling

图7 盐胁迫对海滨木槿幼苗叶片PSⅡ潜在光化学效率的影响(P < 0.05)Figure 7 Effect of NaCl stress on Fv/Foin leaves of H. hamabo seedling
2.3.2 Gs盐胁迫明显抑制了海滨木槿叶片Gs,并且呈现明显的阶段性变化,但在总体上是随着盐胁迫浓度的增加而Gs呈逐渐降低的变化(图3)。在胁迫初期(10 d)受到的抑制作用稍强(图3A),在胁迫中期(20 d)受到抑制作用最强(图3B);在胁迫中后期(30 d)Gs得到一定的恢复,在PAR低于1 000 μmol·m-2·s-1时,0.3%处理组的Gs高于对照组(图3C);在胁迫后期(40 d),0.3%和0.6%处理组的Gs恢复到较高水平,并且在PAR低于1 500 μmol·m-2·s-1时,0.3%和0.6%处理组的Gs高于对照组(图3D)。这表明盐胁迫下不同阶段的Gs呈现不同的变化,随着时间的延长海滨木槿在低、中浓度盐胁迫下的Gs得到明显升高,而高浓度则明显降低了Gs。
从表1可以看出,处理组的AQY在前30天中都显著低于对照(p < 0.05),而在胁迫后期(40 d)时与对照差异不显著,但略低于对照;随着胁迫浓度的增加,AQY基本上呈显著或者明显下降的变化。暗呼吸速率(Rd)在各处理时间段随着胁迫浓度的变化呈不同的变化,胁迫初期(10 d)的Rd呈先升高、下降再升高的变化,其中0.3%处理组的Rd显著高于中盐、高盐浓度的处理组(p < 0.05);胁迫中期(20 d)的Rd随着胁迫浓度的升高呈逐渐下降的变化,并且显著低于对照(P < 0.05);胁迫中后期(30 d)的Rd都显著低于对照(p < 0.05),但随着胁迫浓度的升高而Rd呈逐渐升高的变化;盐分胁迫后期(40 d),各处理组的Rd基本恢复正常水平,甚至0.6%处理组的Rd显著高于其他各试验组(p < 0.05)。光补偿点(LCP)在胁迫初期和中期(10 d和20 d)都呈显著升高的变化(p < 0.05),随着胁迫浓度的逐渐升高初期的LCP呈先升高、后降低、再升高的变化,而中期的LCP呈逐渐升高,最后下降的变化,说明在胁迫条件下海滨木槿对弱光的利用率提高;胁迫中后期的随着胁迫浓度的升高而呈逐渐升高的变化,但是低盐、中盐浓度的LCP显著低于对照(p < 0.05),说明该时期的盐分胁迫降低了海滨木槿的对弱光的利用率;胁迫后期各处理组的LCP在都明显恢复,除了0.6%处理的LCP显著高于对照(p < 0.05)外,其他都明显高于对照但差异不显著(p > 0.05),说明该阶段盐分胁迫下的海滨木槿对弱光的利用率明显提高。盐分胁迫不同程度的降低了植物的最大净光合速率(Pnmax),并且随着胁迫浓度的增加而呈逐渐降低的变化,说明盐分胁迫不同程度地抑制了植物的Pnmax。
我们初见时的玩笑并没有说错。胖子的确是一只功夫熊猫,只是这一生只使出唯一的必杀一招,救回了一个最想拯救的人。爱那么沉重,又轻似尘埃,都化成最后的一句话。

表1 盐分胁迫对海滨木槿幼苗叶片的光合生理参数的影响Table 1 Effect of NaCl stress on photosynthetic parameters in leaves of H. hamabo seedling
2.7 盐分胁迫下海滨木槿幼苗叶绿素含量和光合生理特性指标的相关性分析
盐分胁迫下海滨木槿叶片中总叶绿素含量与表观量子效率呈显著正相关关系(表 2),并且分别与最大净光合速率和暗呼吸速率呈正相关关系,但差异不显著,其中与Pnmax接近正相关关系(其相关系数为0.493,p = 0.052);另外AQY与光补偿点(LCP)呈负相关关系。AQY与Pnmax呈极显著正相关关系,而与 LCP呈极显著负相关关系。Pnmax与LCP呈极显著负相关关系。当盐分胁迫降低了海滨木槿叶片的总叶绿素含量时,显著降低了植物的净光合速率,各光合生理指标间呈显著或极显著的正相关关系。
近5年AJPE的研究趋势为主动学习、跨职业教育、模拟患者以及人文关怀。在这些研究中主要采用定量分析、STATA以及SPSS数据分析软件应用最多。t检验、卡方检验、方差分析、回归分析最为常见。

表2 盐胁迫对海滨木槿叶片叶绿素含量和光合生理指标的相关性分析(n=48)Table 2 Correlation analysis on total chlorophyll content and photosynthetic characteristics under salt stress in H. hamabo leaves
3 讨论与结论
3.1 盐分胁迫下海滨木槿幼苗生长
海滨木槿是海岸带基干林带、盐碱地绿化的优良造林树种[33]。研究证实海滨木槿对盐碱有很强的耐受性,它不仅能在pH 8.0 ~ 8.6、含盐量达1.1% ~ 1.5%的滩涂泥土中正常生长,也能在pH 5.5 ~ 6.5的酸性土中旺盛生长,对土壤的适应能力强,是典型的盐生植物[21]。本研究结果与上述研究结果相一致,但是本试验中死亡1株,可能是短期内高盐环境影响植物的正常代谢水平导致死亡。
3.2 盐分胁迫下海滨木槿幼苗叶片的光合特性
叶绿体是植物光合作用的场所,也是对盐胁迫最敏感的细胞器[34],叶绿素含量不仅是直接反映植物光合能力的重要指标,也是衡量植物耐盐性的重要生理指标之一[35]。在盐胁迫条件下不同植物叶绿素合成与分解的反应机制各异[36~37]。本研究结果中试验初期盐胁迫下总叶绿素含量均呈现下降趋势,且随着胁迫强度的增加而下降,但随着处理时间的延长,总叶绿素含量呈明显上升的趋势,并在试验末期基本恢复,这可能是作为盐生植物,在 NaCl胁迫下海滨木槿耐盐机制启动,致使叶片叶绿素含量表现出波动的变化趋势[25]。不同浓度和处理时间的盐胁迫下海滨木槿的叶绿素含量变化规律不明显,这与其他的研究相一致[22,25]。这进一步证实海滨木槿在盐胁迫条件下,胁迫初期海滨木槿叶片中叶绿素的含量变化与胁迫浓度变化相一致,可能是初期胁迫条件造成了叶绿素的破损,但随着时间的延长盐生植物的耐盐机制启动,叶绿素合成逐渐增加,进而恢复至正常水平。
叶绿素是植物进行光合作用的主要化学物质,不仅是吸收和传递光能的载体,同时也是电子传递过程中必不可缺的电子传递体,是光合作用的必要条件,它的增加可以明显提高植物的光合能力[38~39]。盐胁迫条件下海滨木槿光响应曲线的变化趋势基本上与植物叶片叶绿素含量的变化一致,胁迫条件下不同程度地抑制了海滨木槿叶片的光合速率,这与自然滩涂消浪林[28]和模拟潮汐水淹[29]条件下海滨木槿的研究结果一致。另外相关性分析发现,不同处理时间和处理浓度的总叶绿素含量与最大净光合速率呈正相关关系(相关系数0.493,p = 0.052),进一步证实了叶绿素含量的变化影响着植物的光合速率。
光合系统Ⅱ的光化学效率Fv/Fm和Fv/Fo可反映逆境胁迫对植物光合系统的破坏程度。在盐胁迫下枸杞[40]、葡萄[41]等植物的Fv/Fm和Fv/Fo均不同程度地降低。本研究中海滨木槿在中、高盐胁迫下Fv/Fm和Fv/Fo均在中后期(30 d)时低于对照,表明NaCl胁迫破坏叶绿体光合机构,PSⅡ反应中心受损,抑制光合作用的原初反应,阻碍光合电子传递的过程,明显降低 PSⅡ原初光能转换效率,抑制 PSⅡ潜在活性[42~43],最终导致净光合速率的显著下降,这是植物在逆境下以降低光合速率等为代价维持自身生长的需要。
一般认为胁迫使Pn降低的因素包括气孔限制和非气孔限制,如果胁迫使Gs降低而叶肉细胞仍在活跃地进行光合,Ci降低而 Ls增加,Pn降低是典型的气孔限制所致;如果叶肉细胞本身光合能力降低,使其利用 CO2的能力降低,Ci增加而Gs、Ls降低,Pn降低是非气孔限制所致[44]。本研究中胁迫初期,低盐(0.3%)、中盐(0.6%)浓度处理的海滨木槿的Gs和Ci同时下降而Ls增加,其Pn降低是气孔限制;而高盐(0.9%)浓度处理组的Ci增加而Gs和Ls同时下降,其Pn降低属于非气孔限制。胁迫中期,各处理组的Ci均增加,而Gs和Ls则同时下降,此时各处理组的Pn降低均属于非气孔限制。胁迫中后期,中盐、高盐浓度胁迫的Ci和Gs同时下降而Ls增加,属于气孔限制;低盐度胁迫浓的Ci增加,而Ls下降,而Gs在PAR高于1 000 μmol·m-2·s-1时是下降的,Pn降低属于非气孔限制,但是在PAR低于1 000 μmol·m-2·s-1时Gs是增加的,此时的Pn降低既不属于非气孔限制,又不属于气孔限制。胁迫后期,除了高盐浓度处理组的Gs稍低外,其他处理的Gs、Ci、Ls等均接近对照,并且随着PAR可能略有升高或者降低,并不是严格属于气孔限制或者非气孔限制。这些结果证实,限制光合作用的气孔因素和非气孔因素二者之间并不是相对独立的,而是随着胁迫时间的长短和胁迫浓度的变化而处于动态变化中[45];另外长期胁迫过程中每个时期植物所反表现的光合生理生态特性,可能会更好的反映出植物的耐受特性。
盐胁迫对海滨木槿幼苗的AQY、Rd、Pnmax、LCP等具有明显影响。海滨木槿作为盐生植物,各光合参数对盐胁迫的响应在不同胁迫阶段表现明显差异,并且随着时间的延长,各光合生理参数逐渐恢复,进一步证实海滨木槿对NaCl胁迫具有一定的耐性,且对胁迫伤害具有一定的修复能力[22]。
综上所述,海滨木槿对NaCl胁迫具有较强的耐受能力,并且各光合生理参数与盐有着密切关系;盐胁迫条件下,光合生理参数的光响应呈现出一定的适应性,对胁迫伤害具有较强的自我恢复能力,可以作为滨海盐碱地绿化的优良树种。另外,胁迫条件下不同时期的光合生理参数不仅能更好地反映植物的生态特性,对于了解植物对胁迫环境的适应策略与机制,以及耐性树种的选育等方面都有重要意义。
[1] Munns R. Comparative physiology of salt and water stress [J]. Plant Cell Environ,2002,25(2):239-250.
[2] 杨劲松. 中国盐渍土研究的发展历程与展望[J]. 土壤学报,2008,45(5):835-837.
[3] Munns R. Genes and salt tolerance: bringing them together [J]. New Phytol,2005(167):645-663.
[4] Sun J,Chen S,Dai S,et al. NaCl-induced alternations of cellular and tissue ion fluxes in roots of salt-resistant and salt-sensitive poplar species[J]. Plant Physiol,2009,149(2):1 141-1 153.
[5] 张建锋,张旭东,周金星,等. 世界盐碱地资源及其改良利用的基本措施[J]. 水土保持研究,2005,12(6): 28-30.
[6] Allakhverdiev SI,Sakamoto A,Nishiyama Y,et al. Ionic and osmotic effects of NaCl-induced inactivation of photosystems I and II in Synechococcus sp[J]. Plant Physiol,2000,123(3),1 047-1 056.
[7] Munns R. Physiological processes limiting plant growth in saline soils: Some dogmas and hypotheses [J]. Plant cell Environ,1993,16(1):15-24.
[8] 秦景,贺康宁,谭国栋,等. NaCl胁迫对沙棘和银水牛果幼苗生长及光合特性的影响[J]. 应用生态学报,2009,20(4):791-797.
[9] Yeo A R,Caporn S J M,Flower T J. The effect of salinity upon photosynthesis in rice (Oryza saliva L.) gas exchange by individual leaves in relation to their salt content [J]. J Exp Bot,1985,36(8):1 240-1 248
[10] 范文涛. 海滨木槿——中国锦葵科植物一新记录[J]. 浙江大学学报(农业与生命科学版),1986,12(4):454-455.
[11] 浙江植物志编辑委员会. 浙江植物志(第四卷)[M]. 杭州:浙江科学技术出版社,1993.
[12] 俞慈英,徐树华. 海滨木槿的驯化及开发利用前景[J]. 林业科学研究,1999,12(2):210-213.
[13] 薄鹏飞,孙秀玲,宋杰,等. NaCl胁迫对海滨木槿种子萌发及Na+、K+含量的影响[J]. 安徽农业科学,2008,36(8):3 098-3 100.
[14] 商宏艳,张士花,杜希华. 不同盐处理对海滨木槿种子离子含量及抗氧化酶活性的影响[J]. 山东师范大学学报(自然科学版),2009,24(4):117-120.
[15] 徐森富,汪成丽. 不同温度浸种对海滨木槿种子发芽率的影响[J]. 中国园艺文摘,2010(10):51, 193.
[16] 崔大练,马玉心,俞兴伟,等. Zn2+对海滨木槿种子萌发及根伸长抑制效应的研究[J]. 种子,2011,30(1):45-48.
[17] Li Junmin,Liao Jingjing,Guan Ming,et al. Salt tolerance of Hibiscus hamabo seedlings: a candidate halophyte for reclamation areas [J]. Acta Physiol Plant,2012(34):1 747-1 755.
[18] 於朝广,殷云龙,芦治国,等. 海滨木槿耐寒良种嫩枝扦插繁殖技术[J]. 现代农业科技,2012(24):180, 191.
[19] 王小雪,孙海菁,刘芸,等. 浓硫酸处理对海滨木槿10个家系种子萌发的影响[J]. 应用生态学报,2012,23(11):2 968-2 974. [20] 杨华,杜国坚,汪奎宏. 胁迫环境下海滨木槿生理特性的研究[J]. 浙江林业科技,2008,28(3):43-47.
[21] 薄鹏飞,孙秀玲,孙同虎,等. NaCl胁迫对海滨木槿抗氧化系统和渗透调节的影响[J]. 西北植物学报,2008,28(1):113-118.
[22] 李会欣,吴明,方炎明,等. NaCl胁迫对海滨木槿叶片生理特性的影响[J]. 植物资源与环境学报,2010,19(3):55-61.
[23] 王秀丽,张获,刘红梅,等. 海滨木槿耐盐性的初步研究[J]. 上海交通大学学报,2010,28(3):248-254.
[24] 黄超群,屠娟丽,周金. 盐胁迫对海滨木槿叶片生理指标的影响[J]. 浙江农业科学,2010(4):773-774, 778.
[25] 芦治国,殷云龙,於朝广,等. NaCl胁迫条件下木槿嫁接苗和扦插苗及其砧木海滨木槿的生理差异[J]. 植物资源与环境学报,2011,20(4):49-57.
[26] 孙宏丽,商宏艳,姚叶,等. NaCl与KCl处理对海滨木槿生长特性的影响[J]. 山东林业科技,2011(6):13-16,20.
[27] 王宗星,吴统贵,虞木奎,等. 模拟海平面上升对海滨木槿渗透调节的影响[J]. 生态科学,2011,30(4):389-392.
[28] 周和锋,房聪玲,李会欣,等. 潮间带消浪林海滨木槿光合作用的初步研究[J]. 福建林业科技,2009,36(2):255-258.
[29] 吴统贵,虞木奎,张翠,等. 两种消浪植物光合特征对起垄高度的响应[J]. 辽宁工程技术大学学报(自然科学版),2010,29(4):674-677.
[30] 陈福明,陈顺伟. 混合液法测定叶绿素含量的研究[J]. 浙江林业科技,1984,4(1):4-8.
[31] Farquhar GD,Van Caemmerer S. Modeling of photosynthetic response to environmental conditions[A]. Lange O L,Nobel P S,Osmond C B. Physiological Plant Ecology Ⅱ. Encyclopedia of Plant Physiology[M]. New Series, Berlin: Springer-Verlag, 1982. 549-587.
[32] 刘宇锋,萧浪涛,童建华,等. 非直线双曲线模型在光合光响应曲线数据分析中的应用[J]. 中国农学通报,2005,21(8):76-79. [33] 王奇志,陈雨,孙浩,等. 木槿属植物海滨木槿的研究进展[J]. 中国野生植物资源,2012,31(3):8-10.
[34] 刘建福. NaCl胁迫对澳洲坚果叶片生理生化特性的影响[J]. 西南师范大学学报(自然科学版),2007,32(4):25-29.
[35] 任艳芳,何俊瑜. NaCl胁迫对莴苣幼苗生长和光合性能的影响[J]. 华北农学报,2008,23(4):149-153.
[36] Strogonov BP. Structure and function of plant cell in saline habitats: new trends in the study of salt tolerance [M]. New York: John Wiley &
Sons, Inc, 1973. 78-80.
[37] 刁丰秋,章文华,刘友良. 盐协迫对大麦叶片类囊体膜组成和功能的影响[J]. 植物生理学报,1997,23(2):105-110.
[38] Nieva FJJ,Castillo JM,Luque CJ,et al. Ecophysiology of tidal and non-tidal population of the invading cordgrass Spartina densiflora: Seasonal and diurnal patterns in a Mediterranean climate [J]. Estuar Coast Shelf Sci,2003(57):919-928.
[39] Plus M,Auby I,Verlaque M,et al. Seasonal variations in photosynthetic irradiance response curves of macrophytes from a Mediterranean
coastal lagoon [J]. Aquat Bot,2005(81):157-173.
[40] 许兴,郑国琦,周涛,等. 宁夏枸杞耐盐性与生理生化特征研究[J]. 中国生态农业学报,2002,10(3):70-73.
[41] 邢庆振,郁松林,牛雅萍,等. 盐胁迫对葡萄幼苗光合及叶绿素荧光特性的影响[J]. 干旱地区农业研究,2011,29(3):96-100.
[42] Everard JD,Gucci R,Kann SC,et al. Gas exchange and carbon partitioning in the leaves of celery (Apium graveolens L.) at various levels of root zone salinity [J]. Plant Physiol,1994,106(1): 281-292.
[43] Shabala SN,Shabala SI,Martynenko AI,et al. Salinity effect on bioelectric activity growth Na+accumulation and chlorophyll fluorescence of corn leaves: a comparison survey and prospects for screening [J]. Aust J Plant Physiol,1998,25(5):609-6l6.
[44] Farquhar GD,Sharkey TD. Stomatal conductance and photosynthesis[J]. Annu Rev Plant Physiol,1982(33):317-345
[45] 郑国琦,许兴,徐兆祯,等. 盐胁迫对枸杞光合作用的气孔与非气孔限制[J]. 西北农业学报,2002,11(3):87-90.
Effect of Salt Stress on Growth and Photosynthetic Characteristics of Hibiscus hamabo Seedings
LU Gang1,LI He-peng1*,ZHANG Xiao-mian1,CHEN Yan2,YUE Chun-lei1,GAO Zhi-hui3
(1. Zhejiang Forestry Academy, Hangzhou 310023, China; 2. Zhejiang Changzheng Vocational & Technical College, Hangzhou 310023, China; 3. Zhejiang Forestry Extension Administration, Hangzhou 310020, China)
Simulated salt stress with 0, 0.3%, 0.6%, 0.9% of NaCl was conducted on 2-year seedling of Hibiscus hamabo to study effect on growth and photosynthetic characteristics. Net photosynthetic rate (Pn), stomatal conductance (Gs), intercellular CO2concentrations (Ci) and others were determined on 10th, 20th, 30thand 40thday after salt stress treatments. The results showed that salt stress had no great effect on the growth of H. hamabo seedlings, expect one treated seedling (died 44 days after treatment) with 0.9% of NaCl. Photosynthetic parameters changed with variation of concentration and the first 20 days of the stress. At the 30th days of the stress, chlorophyll content(Chl) of each treatment recovered, and photosynthetic parameters of treatment with 0.3% recovered except dark respiration rate (Rd) and light compensation point (LCP), and that of treatment with 0.6% and 0.9% recovered varying degrees. At the 40thdays of the stress, most of parameters recovered except Rdand LCP of seedlings treated with 0.6% was higher than that of the control, and maximum net photosynthetic rate (Pn max) of seedlings treated with 0.9% was higher than that of the control. The result also indicated that H. Hamabo had strong tolerance to NaCl stress and could recovered from damage of NaCl.
Hibiscus hamabo; salt stress; salt toerance; photosynthetic characteristics; chlorophyll content
S718.5
A
1001-3776(2015)03-0016-010
2014-12-12;
2015-03-25
浙江省省属科研院所专项项目(2013F50002、2014F50017);省院合作项目(2013SY14);浙江省重点科技创新团队项目(2011R50027);“台州湾围垦区植物区系及耐盐性研究与种植示范”
卢刚(1976-),男,浙江安吉人,工程师,硕士,从事森林培育研究和林业技术推广;*通讯作者。
