阳离子脂质体介导RNA干扰的效果评价
2015-12-27韩文琦甄宇红张树彪赵轶男孙永郭鑫王恩霞刘姿孙耀庭
韩文琦,甄宇红,张树彪,赵轶男,孙永,郭鑫,王恩霞,刘姿,孙耀庭
阳离子脂质体介导RNA干扰的效果评价
韩文琦1,甄宇红1,张树彪2,赵轶男2,孙永1,郭鑫1,王恩霞1,刘姿1,孙耀庭1
1 大连医科大学药学院,辽宁 大连 116044 2 大连民族大学生物技术与资源利用国家民委-教育部重点实验室,辽宁 大连 116600

韩文琦, 甄宇红, 张树彪, 等. 阳离子脂质体介导RNA干扰的效果评价. 生物工程学报, 2015, 31(8): 1239–1246.Han WQ, Zhen YH, Zhang SB, et al. Efficacy of RNA interference mediated by cationic liposomes. Chin J Biotech, 2015, 31(8): 1239–1246.
考察自制的肽型阳离子脂质体CDO14作为RNA转染载体的细胞毒性及其运载siRNA进行RNA干扰的效果。通过MTT法检测脂质体对稳定表达荧光素酶的肺癌A549 (Luc-A549) 细胞的毒性。以脂质体为载体将荧光素酶siRNA (Luc-siRNA) 转染至Luc-A549细胞内,用发光仪检测转染细胞内荧光素酶含量,BCA法检测细胞内总蛋白含量。在裸鼠腋下接种Luc-A549细胞,成瘤后尾静脉注射Luc-siRNA和脂质体的复合物,利用活体成像系统检测模型小鼠体内荧光素酶的表达量。细胞毒性实验表明,自制脂质体的毒性与商品脂质体DOTAP相近,低于商品脂质体Lipo2000;细胞转染实验表明自制脂质体作为基因转染载体的转染效率高于DOTAP;体内转染实验表明CDO14作为载体转染效果优于DOTAP。结果表明,肽型阳离子脂质体CDO14具有毒性小、转染效率高等优点,有望作为转染载体用于基因治疗。
阳离子脂质体, 荧光素酶, A549细胞, RNA干扰
RNA干扰 (RNA interference,RNAi) 是生物体内双链RNA介导的同源mRNA降解现象,是一种序列特异性的转录后基因沉默机制[1-2]。RNAi为特异性抑制目的基因的表达提供了有效的方法,目前主要应用于调节目的基因功能,抗病毒和抗肿瘤的基因治疗等领域[3-4]。RNAi研究近年发展迅速,但如何高效转运siRNA,尤其是以较低浓度的siRNA高效沉默目的基因仍是一个急需解决的问题。
目前用于基因转染的载体可分为病毒载体和非病毒载体。虽然病毒作为转运载体具有效率高的优点[5-7],但存在免疫原性高、载体容量小和致癌性等缺点[8-10],限制了它在临床治疗上的应用。非病毒载体中最典型的是阳离子脂质体 (Cationic liposomes),具有可自然降解、无免疫原性、可重复转染等优点[11-15],近年来备受研究者的重视,至今已有数十种阳离子脂质体被研制出来[16]。本文将考察自制的肽型阳离子脂质体CDO14 (图1) 的毒性及其介导RNA干扰的效果。
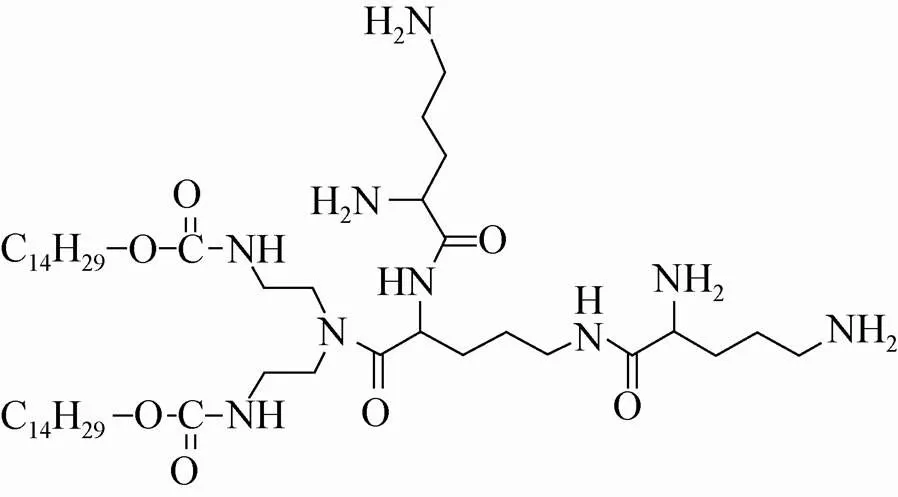
图1 阳离子脂质体CDO14的结构
1 材料与方法
1.1 材料
1.1.1 siRNA与细胞株
siRNA购自Invitrogen公司;表达荧光素酶的人肺癌细胞A549 (Luc-A549) 由美国北卡罗莱纳大学药学院惠赠。
1.1.2 检测试剂及化学试剂
脂质体DOTAP和Lipo2000购自Life公司;CDO14自制;Luciferase Assay Kit购自Promega公司;D-luciferin Potassium Salt购自上海科远迪生物科技有限公司。
1.1.3 实验动物
4−6周雌性BALB/c-nude小鼠购自大连医科大学SPF动物中心,实验动物使用许可证号:SCXK (辽) 2010-0002,于无特异致病原 (SPF) 的环境下饲养。动物饲养和使用符合实验动物管理条例,动物实验经大连医科大学动物伦理委员会批准。
1.1.4 实验仪器
活体成像仪IVIS200购自美国Xenogen公司;发光仪GioMaxTM 96 Microplate Luminometer购自Promega公司;CO2培养箱购自上海智城分析仪器制造有限公司;酶标仪FC购自美国Thermol公司。
1.2 方法
1.2.1 MTT法检测脂质体毒性
将Luc-A549细胞接种于96孔细胞培养板中,每孔加入200 μL含10% FBS不含双抗的DMEM培养基,接种4×103−5×103个细胞。将细胞置于37 ℃、5% CO2培养箱中孵育16−24 h,使细胞密度达80%−90%左右。将DOTAP、Lipo2000、CDO14三种脂质体分别用无血清DMEM稀释,每种脂质体均稀释为0.06、0.006、0.000 6 mg/mL,室温放置5 min,吸弃孔内培养上清液,每孔加200 μL脂质体稀释液,在5% CO2培养箱中37 ℃孵育4−5 h后弃除脂质体稀释液,加入含FBS的DMEM继续培养,48 h后吸弃孔内培养上清液,每孔加入100 μL 0.5 mg/mL MTT,4−6 h后每孔加入100 μL三联液继续培养,12−20 h后用酶标仪检测各孔在570 nm下的吸光值。
1.2.2 siRNA的合成
siRNA由Invitrogen公司合成,Luc-siRNA序列:正义5′-CUUACGCUGAGUACUUCGAT T-3′,反义3′-TTGAAUGCGACUCAUGAAGC U-5′;阴性对照siRNA序列:正义5′-UUCUCCG AACGUGUCACGUTT-3′,反义3′-ACGUGACA CGUUCGGAGAATT-5′,此序列不与任何人类基因序列同源,命名为N-siRNA。
1.2.3 细胞转染
将A549细胞接种于24孔细胞培养板中,每孔加入500 μL含10% FBS不含双抗的DMEM,接种0.5×105−2×105个细胞,将细胞置于37 ℃、5% CO2培养箱中孵育16−24 h使细胞密度达80%−90%左右。将1.5 μg Luc-siRNA或N-siRNA用PBS稀释至50 μL,3 μg脂质体DOTAP或CDO14用PBS稀释至50 μL,将稀释后的siRNA与脂质体混合 (即质量比1︰2),制备的复合物在室温放置20 min,然后每孔加入100 μL上述复合物转染细胞。对照组不加siRNA,只加相同剂量的脂质体。转染的细胞在5% CO2培养箱中37 ℃孵育4−5 h,吸弃原培养液,重新加入500 μL含10% FBS不含双抗的DMEM,继续培养48 h。
1.2.4 发光值的测定
细胞转染48 h后,去除孔中DMEM,用DPBS润洗2次,每孔加入600 μL裂解液 (10%甘油,1% Triton-100溶于DPBS中),室温充分裂解20 min后,取细胞裂解液20 μL移至96孔白板中,加80 μL荧光素酶检测液,使用发光仪测发光值,以发光值表示荧光素酶的量。
1.2.5 总蛋白含量的测定
将250 μL蛋白检测试剂和5 μL细胞裂解液混匀,加到96孔透明板中,37 ℃孵育20 min。使用酶标仪测570 nm下吸光值,计算总蛋白 含量。
以荧光素酶含量/总蛋白含量 (RLU/mg) 表示细胞内RNAi的效果。
1.2.6 裸鼠移植瘤模型的建立
取对数生长期Luc-A549细胞,用胰蛋白酶消化,离心、收集细胞后用PBS洗两次。再次重悬后,用PBS调整体积,使浓度达到 2×107cells/mL。在无菌室内,将裸鼠常规消毒后,在前肢腋下皮下接种上述细胞,每只0.2 mL。接种后,定期观察小鼠的精神、饮食及排便等情况,称量小鼠体重。接种1周左右,可见接种部位皮下长出米粒大小的硬结,为移植瘤模型建成。接种15 d左右,瘤体积 (计算公式V=ab2/2,a和b分别代表移植瘤最大长径和横径) 达到100−200 mm3,开始实验。
1.2.7 裸鼠体内转染
将12只成瘤裸鼠随机分为4组,每组3只,分别为空白对照组、阴性对照组、DOTAP转染组、CDO14转染组。每只裸鼠用量为8 μg siRNA和24 μg脂质体,先将siRNA与脂质体分别稀释于42 μL与76 μL PBS中,然后将siRNA加到脂质体中,混匀,制备的复合物室温放置20 min后,给裸鼠尾静脉注射进行体内转染。
1.2.8 裸鼠体重、瘤体积和活体成像检测
于转染前和转染后48 h测量并记录裸鼠体重,用游标卡尺测瘤大小,并计算瘤体积。活体成像检测体内荧光素酶的表达,裸鼠腹腔注射荧光素3 mg/20 g,自由活动15 min后用异氟烷麻醉,再进入活体成像系统拍照。以转染前后小鼠瘤内的净光密度比表示体内RNAi的 效果。
1.2.9 数据分析
2 结果与分析
2.1 脂质体对Luc-A549细胞的毒性
在给药48 h后,MTT法检测细胞的存活率。由图2可见,在脂质体浓度为0.000 6 mg/mL时,不同脂质体对细胞存活率的影响无显著性差异;当脂质体浓度为0.006 mg/mL时,DOTAP组和Lipo2000组细胞的存活率分别为80%和49%,CDO14组细胞的存活率为79%,自制脂质体组细胞的存活率高于商品脂质体Lipo2000 (<0.05),与DOTAP相当;当脂质体浓度为0.06 mg/mL时,各组的细胞存活率均较低,但CDO14组与DOTAP组相当,明显高于Lipo2000组 (<0.05)。可见,自制脂质体的毒性与DOTAP相当,低于Lipo2000,在后续的实验中将采用DOTAP作为阳性对照。

图2 不同脂质体对Luc-A549细胞的毒性
2.2 脂质体介导的细胞内RNAi
将siRNA转染至Luc-A549细胞,Luc-siRNA转染组荧光素酶的表达量明显低于N-siRNA转染组及未转染siRNA组 (<0.05),N-siRNA转染组和未转染siRNA组间无明显差异,表明合成的Luc-siRNA可在转录水平上抑制荧光素酶报告基因的表达 (图3)。
两种脂质体介导的Luc-siRNA转染组荧光素酶的表达量明显低于无脂质体组 (<0.05);以CDO14为载体组Luc-A549细胞的荧光素酶表达量明显低于以DOTAP为载体组 (<0.05) (图3)。
2.3 脂质体介导小鼠体内RNAi
小鼠腋下接种Luc-A549细胞15 d后进行体内转染,分别在转染前及转染后48 h检测小鼠体内荧光素酶的表达量和小鼠的瘤体积。如图 4所示,在小鼠肿瘤接种部位能观察到荧光素酶的表达,Luc-RNAi特异性转染组荧光素酶表达量在转染后明显降低。如图5所示,随着时间的推移,各组小鼠体内肿瘤细胞增多,瘤体积增大;转染48 h后,空白对照组和N-siRNA非特异性转染组荧光素酶表达量随着肿瘤体积的增加而升高,而两种脂质体介导的Luc-RNAi特异性转染组荧光素酶表达量均显著降低,以脂质体CDO14介导的干扰效果更明显。
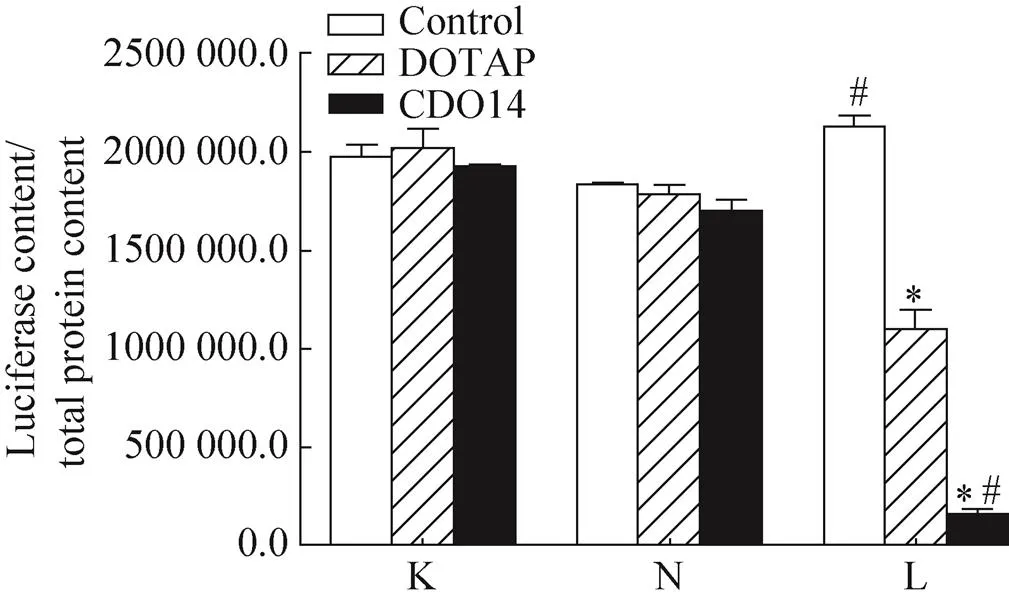
图3 不同脂质体介导Luc-A549细胞内Luc-RNAi
Luc-RNAi特异性转染组与空白组及N-siRNA非特异性转染组相比,转染前后瘤体积比例无显著差异。说明Luc-RNAi并没有影响肿瘤的大小,只是干扰了荧光素酶的表达。
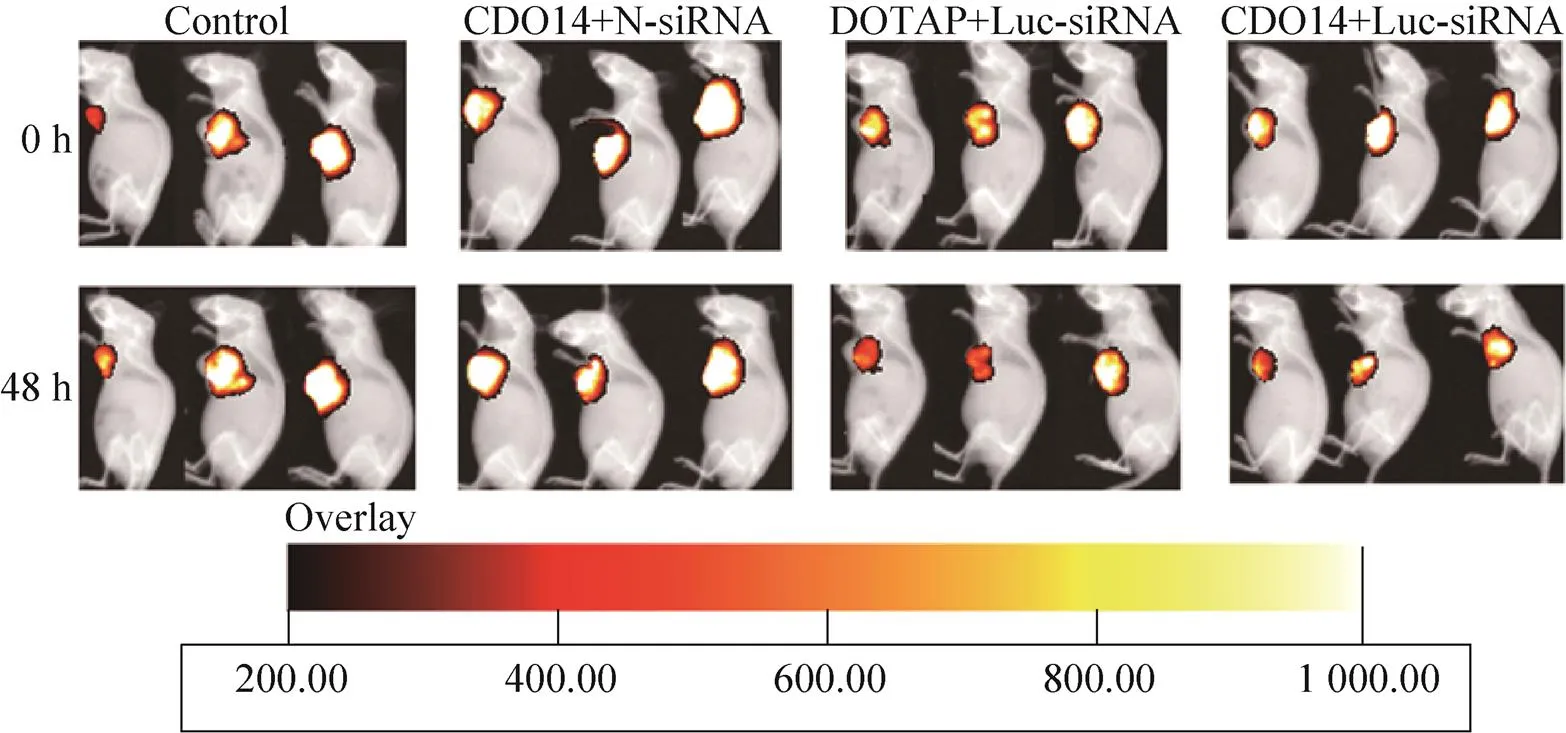
图4 不同脂质体介导小鼠体内Luc-RNAi

图5 转染后48 h与转染前净光密度比和瘤体积比
3 讨论
RNAi是进化上保守的转录后基因沉默途径,是一种具有巨大潜力的人类疾病治疗工具,但如何高效运载siRNA进入靶组织已成为限制RNAi应用的主要障碍[17]。阳离子脂质体是一种由多聚阳离子磷脂和胆固醇构成的表面带有正电荷的脂质体复合物,是目前公认的转染效率较高的非病毒载体之一,但有一定的细胞毒 性[18]。其介导基因转染的机制为:带正电荷的阳离子脂质体通过静电作用与带负电的基因 (DNA或RNA) 形成脂质体-基因复合物,通过细胞内吞或细胞膜融合作用进入细胞内,进一步释放基因,从而在细胞内转录和翻译[19]。
肽型阳离子脂质体与其他类型的阳离子脂质体相比具有良好的生物降解能力和生物相容性,并具有靶向性,近年来受到广泛关注[20-21]。阳离子脂质体作为基因转染载体,转染效果与脂质体的氨基酸、多肽等头部有关,而连接键决定了其稳定性和生物降解性[22-23]。本研究中考察的肽型阳离子脂质体CDO14的极性头部是由3个鸟氨酸形成的三肽;疏水尾部是两个14碳烷基链,连接键为氨基甲酸酯。细胞毒性实验表明该脂质体的细胞毒性与常用的商品脂质体DOTAP相近,其作为siRNA转染载体,在体内外的RNAi效果均优于DOTAP。DOTAP的极性头部是季铵盐,其与基因的结合域比肽型头部要小,因而其结合力可能会弱。此外,DOTAP中的连接键是酯键,而CDO14中的连接键是氨基甲酸酯键,相对而言后者具有更好的稳定性。DOTAP的季铵盐头部可能使其毒性大于肽型头部的脂质体,我们在考察脂质体对A549细胞毒性的实验中得到的结果是二者毒性相近,但对于其他细胞及在体内的毒性如何,还有待于深入研究。
为了评价体内siRNA干扰效果,我们将稳定表达荧光素酶的细胞株Luc-A549接种小鼠腋下,在裸鼠体内稳定成瘤,建立了表达荧光素酶的小鼠移植瘤模型。该模型与传统的肿瘤动物模型相比,能够通过活体成像技术在空间和时间分布上反映细胞内荧光素酶的表达,便于检测活体动物体内肿瘤的生长和转移情况,同时可以对同一个研究个体进行长时间反复跟踪成像,以便长期观察,并减少了不同实验动物之间的个体差异[24-25]。
4 结论
本实验结果表明肽型阳离子脂质体CDO14 与商品脂质体相比具有毒性小、转染效率高等优点,有望作为转染载体用于基因治疗。但此类脂质体在其他细胞中的毒性及转染效果,以及其体内毒性还需进一步研究。
[1] Halic M, Moazed D. Dicer-independent primal RNAs trigger RNAi and heterochromatin formation. Cell, 2010, 140(4): 504−516.
[2] Sun P, Zhao W, Liu YL. Principle and application of RNA interference technology. Med Recapitul, 2011, 17(2): 164−167 (in Chinese).孙平, 赵微, 刘毅玲. RNA干扰技术的原理与应用. 医学综述, 2011, 17(2): 164−167.
[3] Ntziachristos V, Ripoll J, Wang LV, et al. Looking and listening to light: the evolution of whole-body photonic imaging. Nat Biotechnol, 2005, 23(3): 313−320.
[4] Notting IC, Buijs JT, Que I, et al. Whole-body biolum inescent imaging of human uvealmel-anoma in a new mouse model of local tumor growth and metastasis. Invest Ophthalmol Vis Sci, 2005, 46(5): 1581−1587.
[5] Crystal RG. Transfer of genes to humans: early lessons and obstacles to success. Science, 1995, 270(5235): 404−410.
[6] Chandrashekhar V, Srujan M, Prabhakar R, et al. Cationic amphiphiles with fatty acyl chain asymmetry of coconut oil deliver genes selectively to mouse lung. Bioconjug Chem, 2011, 22(3): 497−509.
[7] Xiu KM, Zhao NN, Yang WT, et al. Versatile functionalization of gene vectors via different types of zwitterionic betaine species for serum-tolerant transfection. Acta Biomater, 2013, 9(7): 7439−7448.
[8] Cockrell AS, Kafri T. Gene delivery by lentivirus vectors. Mol Biotechnol, 2007, 36(3): 184−204.
[9] Donahue RE, Kessler SW, Bodine D, et al. Helper virus induced T cell lymphoma in nonhuman primates after retroviral mediated gene transfer. J Exp Med, 1992, 176(4): 1125−1135.
[10] Sun JY, Anand-Jawa V, Chatterjee S, et al. Immune responses to adeno-associated virus and its recombinant vectors. Gene Ther, 2003, 10(11): 964−976.
[11] Nakagawa O, Ming X, Huang L, et al. Targeted intracellular delivery of antisense oligonucleotides via conjugation with small-molecule ligands. J Am Chem Soc, 2010, 132(26): 8848−8849.
[12] Bishop CJ, Ketola TM, Tzeng SY, et al. The effect and role of carbon atoms in poly (β-amino ester) s for DNA binding and gene delivery. J Am Chem Soc, 2013, 135(18): 6951−6957.
[13] Clerici AM, Faraoni P, Ferrari C, et al. Carborane derivatives loaded into liposomes as efficient delivery systems for boron neutron capture therapy. J Med Chem, 2009, 52(23): 7829−7835.
[14] Zhang S, Zhao Y, Zhao B, et al. Hybrids of nonviral vectors for gene delivery. Bioconjug Chem, 2010, 21(6): 1003−1009.
[15] Zhi D, Zhang S, Cui S, et al. The headgroup evolution of cationic lipids for gene delivery. Bioconjug Chem, 2013, 24(4): 487−519.
[16] Hyoudou K, Nishikawa M, Umeyama Y, et al. Inhibition of metastatic tumor growth in mouse lung by repeated administration of polyethylene glycol-conjugated catalase: quantitative analysis with firefly luciferase-expressing melanoma cells. Clin Cancer Res, 2004, 10(22): 7685−7691.
[17] Mandl SJ, Mari C, Edinger M, et al. Multi-modality imaging identifies key times for annexinV imaging as an early predictor of therapeutic outcome. Mol Imaging, 2004, 3(1): 1−8.
[18] Guo B H, Zhang J S, Xie Y J. Development of cationic liposomes in research of gene delivery. Prog Pharm Sci, 2000, 24(1): 1−4.
[19] Tresset G. The multiple faces of self-assembled lipidic systems. PMC Biophys, 2009, 2(1): 3−4.
[20] Rea JC, Gibly RF, Barron AE, et al. Self-assembling peptide-lipoplexes for substrate-mediated gene delivery. Acta Biomater, 2009, 5(3): 903−912.
[21] Rajesh M, Sen J, Srujan M, et al. Dramatic influence of the orientation of linker between hydrophilic and hydrophobic lipid moiety in liposomal gene delivery. J Am Chem Soc, 2007, 129(37): 11408−11420.
[22] Lamanna CM, Lusic H, Camplo M, et al. Charge-reversal lipids, peptide-based lipids, and nucleoside-based lipids for gene delivery. Acc Chem Res, 2012, 45(7): 1026−1038.
[23] Ashley CE, Carnes EC, Epler KE, et al. Delivery of small interfering RNA by peptide-targeted mesoporous silica nanoparticle-supported lipid bilayers. ACS Nano, 2012, 6(3): 2174−2188.
[24] Wang X, Rosol M, Ge S, et al. Dynamic tracking of human hematopoietic stem cell engraftm entusingbiolum inescene imaging. Blood, 2003, 102(10): 3478−3482.
[25] Zhang HY, Li Y, Wang Z, et al. Establishment of xenograft mouse models to study human lung cancer by usingimaging system. Chin J Biotech, 2009, 25(8): 1204−1210 (in Chinese).张海燕, 李艳, 王喆, 等. 用于活体成像的小鼠肺癌移植瘤模型的建立. 生物工程学报, 2009, 25(8): 1204−1210.
(本文责编 郝丽芳)
Efficacy of RNA interference mediated by cationic liposomes
Wenqi Han1, Yuhong Zhen1, Shubiao Zhang2, Yinan Zhao2, Yong Sun1, Xin Guo1, Enxia Wang1, Zi Liu1, and Yaoting Sun1
1,,116044,,2,,116600,,
To investigate the cytotoxicity of the homemade peptide cationic liposome CDO14 and its efficacy of RNA interference (RNAi). MTT method was used to determine the cytotoxicity of the liposome to a human lung cancer cell line Luc-A549 that can express luciferase stably. Luciferase siRNA (Luc-siRNA) was transfected into Luc-A549 cells by CDO14. Contents of luciferase in the transfected cells were detected by luminous instrument and contents of total protein in these cells were detected by BCA method. Nude mice were inoculated with Luc-A549 cells in axilla to establish xenograft tumor model. Complexes of Luc-siRNA and the cationic liposomes were injected into the modeling mice via tail vein. Contents of luciferase in the transfected mice were detected by the whole body imaging system. The cytotoxicity of the homemade cationic liposome was similar to that of commercial liposome DOTAP, and lower than that of Lipo2000. The siRNA transfection efficacy mediated by CDO14 was higher than that mediated by DOTAP. The homemade peptide cationic liposome CDO14 is expected to serve as delivery vector in gene therapy because of its low cytotoxicity and high transfection efficiency.
cationic liposome, luciferase, A549 cell, RNAi
10.13345/j.cjb.140520
October 29, 2014; Accepted: December 10, 2014
National Natural Science Foundation of China (Nos. 20876027, 21176046), National High Technology Research and Development Program of China (863 Program) (No. 2014AA020707), Fundamental Research Funds for the Central Universities (No. DC12010104), Natural Science Foundation of Liaoning Province (No. 2013023037).
Yuhong Zhen. Tel/Fax: +86-411-86110414; E-mail: zhenyhwaner@aliyun.com Shubiao Zhang.Tel/Fax: +86-411-87656141; E-mail: zsb@dlnu.edu.cn
国家自然科学基金 (Nos. 20876027, 21176046),国家高新技术研究发展计划 (863计划) (No. 2014AA020707),中央高校自主科研基金 (No. DC12010104),辽宁省自然科学基金 (No. 2013023037) 资助。
2015-02-27
http://www.cnki.net/kcms/detail/11.1998.Q.20150227.1139.005.html
