大分舌蜂主要生物学特性研究
2015-12-16黄敦元苏田娟余江帆牛泽清朱朝东
黄敦元,谷 平,苏田娟,余江帆,何 波,牛泽清,朱朝东
(1.江西环境工程职业学院,江西赣州 341000;2.中国科学院动物研究所,动物进化与系统学(院)重点实验室,北京1 001013;3.江西省林业科技培训中心,南昌 330038)
大分舌蜂Colleles gigas Cockerell 隶属膜翅目Hymenoptera 分舌蜂科Colletidae 分舌蜂属Colletes Latreille。主要分布在江西、湖南、浙江、福建等中国南方地区(吴燕如,1965;Kuhlmann,1999;Niu et al.,2013)。在长期的进化过程中,大分舌蜂成虫的活动期与油茶的开花物候相适应,是油茶花期主要传粉昆虫之一(吴燕如,2001;赵延会等,2010)。该蜂羽化后即可完成交配、筑巢,访问油茶花采集花粉和花蜜制作蜂粮并产卵繁育后代,根据其访花的习性,参照Gauld & Bolton(1988)的蜜蜂分类方法,大分舌蜂为典型寡访花性类群(Oligolectic)。
分舌蜂科是蜜蜂总科Apoidea 中较原始的类群,是典型的独栖性种类(Wafa et al.,1972;Litt,1988)。世界范围内共有56 属700 余种(Michener,2007;Kuhmann,2009),到目前为止,世界范围内已研究了大约24种分舌蜂属物种的巢穴(赵延会等,2010)。我国在分舌蜂科研究方面相对比较滞后,到目前为止,仅记录了分舌蜂属和叶舌蜂属Hylaeus 共4种(吴燕如,1965;赵延会等,2010)。
油茶Camellia oleifera Abel 系山茶科Theaceae山茶属Camellia 植物,是我国重要的木本油料树种之一,也是我国基本的木本食用油料树种。油茶种植遍布我国中南、西南16个省区及越南等国(Zhuang,1998),目前我国的油茶种植总面积达300 万hm2(吕翠萍等,2011)。早在20 世纪80年代,国内已有学者报道可通过增加油茶地蜂Andrena camellia Wu、大分舌蜂、纹地蜂Andrena striata Wu 等野生蜜蜂的种群密度来提高油茶的座果率(庄瑞林,1998)。
1 材料与方法
1.1 试验地点
野外调查的时间从2011年10月到2014年12月。调查地点主要在江西省新余市渝水区水北样地(E:115°04',N:28°04'),江西省宜春市袁州区巫家样地(E:114°12',N:27°45')和广东省河源市东源样地(E:114°98',N:24°19'),其中新余市水北样地坡度很小,土质为红壤(第四纪红色粘土),地表少有浅草,裸土较多,树龄为12年的高产油茶人工林,大分舌蜂巢穴主要集中分布在油茶林区周边的杉树林里;宜春巫家样地和广东东源样地有一定坡度,油茶为成年老林且缺乏管理,大分舌蜂巢穴主要分布在山体斜坡上。
1.2 研究方法
1.2.1 生活史的观察
通过对野外标记好的大分舌蜂巢穴进行定期挖掘(幼虫期每10 d 挖掘2个巢穴,卵期及蛹期每2 d 挖掘3-5个巢穴),观察、拍照并用游标卡尺测量记录相关数据完成大分舌蜂的生活史观察。
1.2.2 蜜源植物观察
在大分舌蜂成虫活动期通过野外跟踪观察该蜂(包括雌蜂和雄蜂)到访植物种类,同时通过对巢穴中蜂粮花粉成份的镜检(100×)来确定大分舌蜂的蜜源植物。
1.2.3 羽化出巢及性比的观察
随机选择100个巢穴进行套杯处理,观察每个巢穴中大分舌蜂的羽化出巢规律。通过对20个不同样地的巢穴进行挖掘并将采集到的蛹带回实验室进行观察,统计出羽化出巢的个体数目及该蜂的性比。
1.2.4 成虫交配行为的观察
制作规格为3 m×3 m×3 m 的网罩安置在蜂巢附近,将羽化出巢的雌雄个体放置在网罩内观察该蜂的交配行为。
1.3 数据统计与分析
实验数据存储和图形制作采用SPSS 13.0 和Excel 2003 软件进行处理。
2 结果与分析
2.1 生活史
大分舌蜂在江西新余、宜春等中国南方地区一年发生1 代,主要以滞育状态下的老熟幼虫在虫室(zooecium)中越夏,成虫通常在每年的10月上旬开始羽化并于11月中旬陆续出巢活动,一般雄性大分舌蜂较雌性早4-6 d 出巢,同一个巢穴中成虫的出巢历期在15 d 左右。油茶的盛花期(一般每年的11月中旬)是该蜂成虫的活动高峰期。12月中旬成蜂数量开始减少,12月底基本不见其活动。雄蜂寿命约25 d,雌蜂寿命约38 d。卵期约8 d,幼虫活动期约24 d。幼虫取食完花粉球后开始进入滞育期,滞育期较长,历时约230 d。于翌年9月中旬开始化蛹,蛹期约35 d。成虫于11月中旬陆续羽化出巢,在羽化后当天即可开始交配,交配后的雌蜂多数在羽化地点附近选址筑巢。大分舌蜂巢穴为单支近竖直的主道或是近水平的主道,筑巢过程中,成蜂在挖掘到一定深度后(一般在40 cm 左右)开始在主道附近修筑虫室,修筑好一个虫室,然后开始外出采集山茶属植物的花粉花蜜到虫室中制作花粉球(蜂粮)。大分舌蜂虫室内有该蜂唾液腺分泌物和杜氏腺分泌物混合并凝固而成的玻璃纸状膜包裹蜂粮(Torchio et al.,1988;Michener,2007),这种膜具有防水保湿的功能(赵彦会等,2010),待一个花粉球制作完成后,雌蜂在其上产卵1 粒并封闭包裹膜和虫室通道,然后开始继续挖掘并修筑下一个虫室(赵延会等,2010;黄敦元等,2014)。大分舌蜂的年生活史见表1。
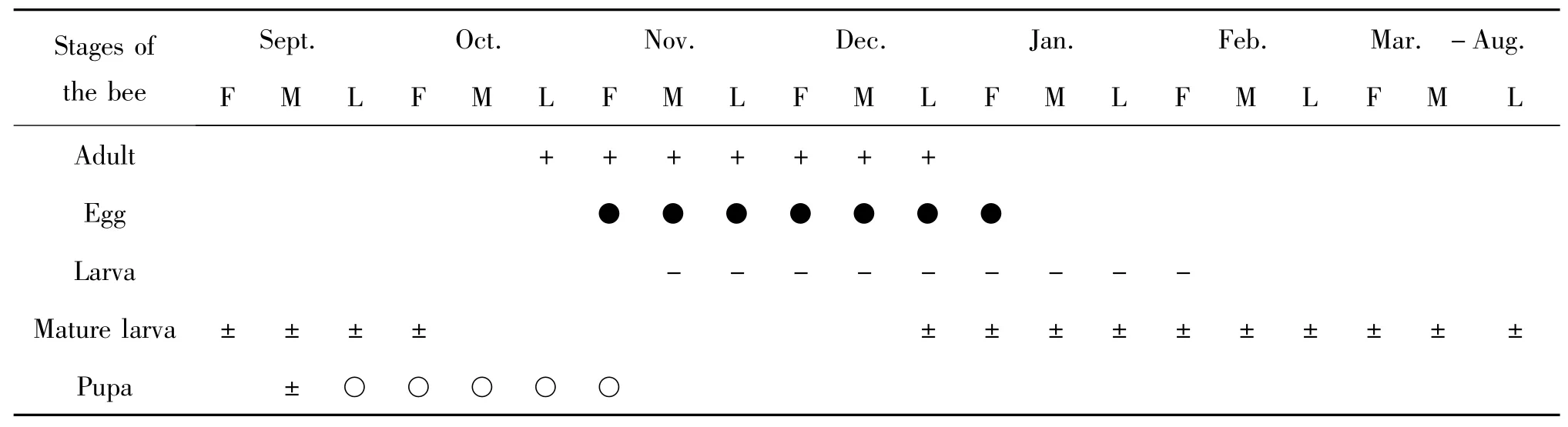
表1 大分舌蜂的生活史(江西新余)Table 1 The life history of Colleles gigas Cockerell(Xinyu,Jiangxi)
2.1.1 卵
大分舌蜂卵近似微弯曲的长圆柱形,两端略钝尖,其中一端有气室结构(air space),长约2.88±0.26 mm(n=10),直径0.93±0.09 mm(n=10)。产在糊状蜂粮的上表面。刚产的卵无色透明略带乳白色,卵壳膜状,表面光滑。大约8 d后,卵壳内幼虫的形态发育完成,虫体缓慢蠕动,然后以头部破卵而出,卵壳从背中线逐渐开裂。
2.1.2 幼虫
由于大分舌蜂幼虫在土壤虫室中的玻璃纸状膜包裹中发育且幼虫早期蜕皮现象不明显。在本研究中,我们主要根据幼虫的发育情况和对花粉球的消耗情况将大分舌蜂的幼虫大致分成三个阶段:低龄幼虫(前3 龄),高龄幼虫(4、5 龄)和老熟幼虫。
初孵幼虫以腹面与卵壳相连,呈半透明的浅乳白色,长3.45±0.24 mm(n=10),宽1.23±0.08 mm(n=10);取食时,头部没入蜂粮,身体不停地收缩和吞食蜂粮。前3 龄的低龄幼虫生长缓慢,对蜂粮的消耗量不大,大约消耗蜂粮的1/8,体表光滑且分节现象不明显。
随着虫体的长大,到4、5 龄时幼虫明显变大,节间褶皱逐渐由浅变深,粘在身体两侧卵壳也碎裂成条状,幼虫的颜色变成了乳白色。大约5 d后,有1 次明显的蜕皮过程,刚蜕皮的幼虫褶皱明显,身体逐渐呈“C”型,且对花粉的需求量开始增大,幼虫生长迅速,消耗完所有的蜂粮后进入老熟幼虫阶段并滞育。
老熟幼虫体呈粗壮“C”型,体表光滑无毛,呈乳白色,体长18.65±0.43 mm(n=10),宽7.83±0.25 mm(n=10)。头部宽圆,有明显的触角乳突,上颚具2 齿(下口式),下颚须及下唇须明显可见,唾泵开口处为一大缝,且缝的外缘具唇状边缘。大分舌蜂以滞育的老熟幼虫越夏,滞育期约230 d。
2.1.3 蛹
离蛹。雌雄个体蛹的大小差异明显,雌性蛹体长18.45±0.83 mm(n=10),宽7.22±0.42 mm(n=10);雄性蛹体长15.91±0.15 mm(n=10),宽6.88±0.12 mm(n=10)。体色初期乳白色,逐渐由乳白色-淡黄色-橘黄色-红褐色-黑色;头胸部的颜色先于腹部变暗;腹部节间处随头胸部同时变黑,其余部位由红褐色逐渐至黑色。复眼1 对,单眼3 只,位于颅顶两复眼中间,呈倒三角形排列。初期单、复眼均无色,约3 d 后变为浅粉色,并逐渐由浅粉色-红色-深红-黑色。在整个蛹期,复眼的颜色变化早于腹部节间早于头胸部早于腹部。在腹部节间变成黑色的时候开始出现一对前翅(开始是无色透明的),7 d 后长出一对后翅。整个蛹期约38 d,羽化完成后变成成虫,自己打通虫室通道并从主道爬出巢口。
2.1.4 成虫
雌成虫:体长17-18 mm;黑色体被黄褐色毛。头宽于长;颅顶后缘凹陷;唇基刻点较粗,呈纵排排列,中央者稀疏;颜面刻点较密,颅顶上的细小;中胸背板刻点密;腹部较长,背板刻点不明显。翅浅褐色,翅脉及翅基片褐色。体被黄褐色毛;颜面、颅顶、颊、胸部背板及侧板、并胸腹节及腹部第1-2 节背板基部均密被黄褐色毛;腹部第1-4 节背板后缘具黄色宽毛带,第3-6 节背板被短的黄褐色毛,其中以5-6 节者较密;足被浅黄色毛,尤以后足转节及腿节者最长,可黏着花粉,起花粉刷(scopa)的作用。
雄成虫:体长14-15 mm。与雌蜂的主要区别是:个体较雌性明显小,体毛长而密;唇基及颜面被长毛;腹部较短,其中1-6 节背板后缘被黄色宽毛带。
雄蜂个体较小,基本不具有采粉器官(花粉篮和腹毛刷)(Pollen Basket and Pollen Press),专司交尾,访问油茶花主要是为吸食花蜜,体毛少,也会沾少许花粉,但是会经常停留在叶片上通过第一对胸足来回上下摩擦胸部(Thorax)和腹部(Abdomen)卸粉,基本不具有传粉作用。
雌蜂个体较大,具特有采粉器官(花粉篮和腹毛刷)采集花粉并吸食花蜜而不伤害花朵,体表多毛、易沾花粉。雌蜂承担筑巢、制作蜂粮、产卵等任务。
2.2 大分舌蜂的蜜源植物
大分舌蜂为典型独栖性野生蜜蜂,雌蜂采集蜜源植物的花粉和花蜜制作蜂粮作为繁育后代的食物,通过野外观察和室内镜检发现:雌蜂主要到访油茶和茶Camellia sinensis L.的花采集花粉和花蜜并带回巢穴制作蜂粮;偶有到访黄荆Vitex negundo Linn.等油茶同花期花,主要采集少许花蜜补充自身营养,但是不采集花粉。同时在实验室通过对巢穴内花粉球和雌蜂花粉篮中花粉在解剖镜下镜检(100×),确认大分舌蜂的蜜源植物为油茶和茶等山茶属植物,所以大分舌蜂为典型寡食性野生蜜蜂。雄性大分舌蜂不采集花粉和花蜜来繁育后代,到访开花植物仅仅是采集花蜜维持成虫个体自身营养需要,主要到访植物种类有:油茶、茶、黄荆、马兰Kalimeris indica、地桃花Urena lobata 和千里光Senecio scandens 等。
2.3 成虫羽化出巢及性比
成虫羽化出巢一般在油茶初花期,即:每年10月中旬开始羽化出巢(江西新余)。羽化出巢的蜂先在地面爬行并震动翅膀直到翅膀干燥并能飞行,同时会排泄体内粪便。一般大分舌蜂巢口直径大小是:9.62±0.45 mm(n=30)。我们通过对20个不同样地的巢穴进行挖掘并将采集到的蛹带回实验室进行统计,共收集到雌蜂蛹461 头,雄蜂163 头,雌雄比约为3∶1。
2.4 成虫交配行为
雄蜂多在晴好天气上午10∶00 左右羽化出土,刚出巢的雄蜂先寻找花朵,取食花蜜,以补充营养并返回羽化巢区附近等待雌蜂羽化出巢并完成交配,夜晚和大风低温天气躲到隐蔽的树叶下休息。雌蜂的出巢时间多数集中在晴好天气的9∶00-17∶00,雌蜂出巢后一般在当天便可与雄蜂交尾,交尾地点多在巢口附近或是蜜源植物附近。
由于雄蜂羽化较雌蜂早且大多集中在巢口附近的杂草和灌木周围,所以在巢口和蜜源植物附近都有大量企图交配的雄蜂。一旦发现雌蜂,雄蜂迅速追逐与之交尾。交尾时雄蜂用上颚(mandible)和足抓住雌蜂,并落在其背上,不断用前足拨动雌蜂的前胸背部,待雌蜂腹部末端上翘表示接受交尾时,雄蜂伸长并向下弯曲腹部,翻出阳茎,插入雌蜂的生殖孔。交尾持续约50.69±5.37 s(n=10)后,雌蜂挣脱雄蜂飞走,有时雄蜂试图与该雌蜂再次交尾,但未见成功。由此推断,1 头雄蜂可与多头雌蜂交尾,而雌蜂则一生可能只交尾1 次。交尾后的雌蜂如果遇到再次来交配的雄蜂时,一般雌蜂腹部末端下翘,振动翅膀并迅速从叶片上坠落而逃离。
交配成功的雌蜂开始选址筑巢、访花、采集花粉和花蜜制作蜂粮。当雌蜂开始筑巢后,仍有雄蜂前来试图交尾,但都遭到拒绝。完成筑巢和做好花粉球后,雌蜂将卵产在花粉球上,1 头雌蜂一生产卵22-34 粒,多数则26 粒。
3 结论与讨论
低龄幼虫蜕皮本身不易观察,而且大分舌蜂是地下营巢并在土壤的虫室中完成其发育过程,其巢穴及虫室内的适宜温度、适宜湿度等因素,故目前还无法将大分舌蜂移入实验室来饲养和观察(蛹期之前)。这样导致目前还无法具体描述大分舌蜂卵的发育过程和幼虫的具体蜕皮过程,无法精确确定幼虫龄期。
温度对昆虫的发育速率尤其是对昆虫幼虫的发育有非常明显的影响(Irlich et al.,2009;Shi et al.,2011)。据野外对其巢穴挖掘发现:通常情况下,大分舌蜂幼虫的虫室一般在60-120 cm 深度的土壤中(土质极差巢址除外)。在冬季气温较低的时候,像这样深度的土壤中,土壤温度要明显高于气温且日变化幅度较小,这对大分舌蜂卵和幼虫的发育是非常适宜的。在野外观察中发现:同一块样地,光照比较好的地方大分舌蜂成虫出巢的时间相对要比树荫下早3-5 d,这说明光照、土壤温度等因素可能对大分舌蜂的发育速率有一定的影响,具体影响规律还需要进一步的研究。
分舌蜂属类群的巢穴结构一般是由一个主道和一些与主道直接相连的虫室道组成,但有的种类在一个巢穴内只建一个虫室,因此没有分支(Torchio,1965;Stephen et al.,1969)。分舌蜂属中也存在同一个虫室道内有2 到多个虫室的情况(Claude-joseph,1926;Michener and Lange,1957;Michener,2007)。本研究中的大分舌蜂巢穴结构基本是由一个主道和一些与主道直接相连的虫室道组成,每个虫室道内只建一个虫室。
油茶初花期的雨量及土壤湿度是地栖野生蜜蜂选择筑巢地址的主要影响因素之一(黄敦元等,2008)。大分舌蜂一般选择在平地或是坡面上筑巢且两种情况下其巢穴结构内部差异较大,具体原因有待于进一步研究。
大分舌蜂雌性成虫个体要远大于雄性,这种差异也同样体现在雌性与雄性在虫室、幼虫及蛹的大小上。这种各个性状在性别上的差异在其他分舌蜂属物种中也有过报道(Rooijakkers and Sommeijer,2009)。研究还发现雌性虫室中的蜂粮也比雄性多,因此可以提供更多的食物供其发育成较大的个体。由于较大的虫室需要的空间更大,因此大分舌蜂在挖掘巢穴前就应该已经确定该虫室中将产何种性别的卵(Rooijakkers and Sommeijer,2009),这对其在雌性与雄性后代进行相应合理的投入有重要意义。







References)
Claude-Joseph F.Recherches biologiques sur les Hymenopteres du Chile(Melliferes)[J].Annales des Sciences Naturelles,Zoologie,1926,9:113-268.
Huang DY,Ding L,Zhang YZ,et al.Life history and relevant biological features of Andrena camellia Wu(Hymenoptera:Andrenidae)[J].Acta Entomologica Sinica,2008,51(7):778-783.[黄敦元,丁亮,张彦周,等.油茶主要野生传粉地蜂(Andrena camellia Wu)生活史及相关生物学习性研究[J].昆虫学报,2008,51(7):778-783]
Huang DY,Gu P,Yu JF,et al.2014.The habitats and foraging behaviors of Colletes gigas(Hymenoptera,Colletidae)[J].Journal of Environmental Entomolgy,2014,36(3):315-321.[黄敦元,谷平,余江帆,等.大分舌蜂的栖息地环境和访花规律[J].环境昆虫学报,2014,36(3):315-321]
Irlich UM,Terblanche JS,Blackburn TM,et al.Insect rate-temperature relationships:environmental variation and the metabolic theory of ecology[J].The American Naturalist,2009,174(6):819-835.
Kuhlmann M.Colletes wolfi spec,nova from Italy,and Lectotype designation for palaearctic bees of the genus colletes LATR.,with notes on new Homonymies and Synonymies(Hymenoptera:Apidae:Colletinae)[J].Linzer Biologische Beiträge,1999,31(1):71-81.
Kuhlmann M.Katalog der paläarktischen arten der bienengattung colletes latr.,mit lectotypenfestlegungen,neuer synonymie und der beschreibung von zwei neuen arten(Hymenoptera:Apidae:Colletinae)[J].Linzer Biologische Beiträge,2000,32:155-193.
Kuhlmann M.Erster nachtrag zur kenntnis der bienengattung colletes latreille 1802 in der mongolei mit beschreibung einer neuen art(Hymenoptera,Apiformes,Colletidae)[J].Beiträge zur Entomologie,2009,59:19-32.
Lv CP,Luo ZW,Liu H,et al.Research advances of Camellia oleifera in China[J].Journal of Anhui Agricultural Sciences,2011,39(26):16177-16179.[吕翠萍,罗正伟,刘虹,等.我国油茶研究进展[J].安徽农业科学,2011,39(26):16177-16179]
Michener CD,Lange RB.Observations on the ethology of some Brazilian colletid bees[J].Journal of the Kansas Entomological Society,1957,30:72-80.
Michener CD.The Bees of the World[M].2nded.Baltimore:The Johns Hopkins University Press,2007,953.
Niu ZQ,Kuhlmann M,Zhu CD.A review of Colletes succinctus group(Hymenoptera:Colletidae:Colletinae:Colletes)from China with redescription of the male of C.gigas[J].Zootaxa,2013,3626(1):173-187.
Rooijakkers EF,Sommeijer MJ.Gender specific brood cells in the solitary bee Colletes halophilus(Hymenoptera;Colletidae)[J].Journal of Insect Behavior,2009,22(6):492-500.
Shi PJ,Ge F,Sun YC,et al.A simple model for describing the effect of temperature on insect developmental rate[J].Journal of Asia-Pacific Entomology,2011,14(1):15-20.
Stephen WP,Bohart GE,Torchio PF.The Biology and External Morphology of Bees[M].Corvallis:Oregon State University,Agricultural Experiment Station,1969,140.
Torchio PF,Trostle GE,Burdick DJ.The nesting biology of Colletes kincaidii Cockerell(Hymenoptera:Colletidae)and development of its immature forms[J].Annals of the Entomological Society of America,1988,81(4):605-625.
Torchio PF.Observations on the biology of Colletes ciliatoides(Hymenoptera:Apoidea,Colletidae)[J].Journal of the Kansas Entomological Society,1965,38(2):182-187.
Wu YR.Economic Insects of China.No.9,Hymenoptera:Apoidea[M].Beijing Science Press,1965,83.[吴燕如.中国经济昆虫志.第九册.膜翅目·蜜蜂总科[M].北京:科学出版社,1965,83]
Wu YR.The behavior of bee[J].Bulletin of Biology,2001,36:1-3.[吴燕如.蜜蜂的行为[J].生物学通报,2001,36:1-3]
Zhao YH,Ding L,Yuan F,et al.Nesting biology of Colletes gigas(Hymenoptera:Colletidae)[J].Acta Entomologica Sinica,2010,53(11):1287-1294.[赵延会,丁 亮,袁峰,等.大分舌蜂营巢生物学[J].昆虫学报,2010,53(11):1287-1294]
Zhuang RL.The Camellia oleifera of China[M].2nded.Beijing:China Forestry Publishing House,1998,1-20.[庄瑞林.中国油茶[M].北京:中国林业出版社,1998,1-20]
