白骨壤幼苗对镉胁迫的生长及生理生态响应
2015-12-01陈昌徐陆志强郑文教
陈昌徐,陆志强,郑文教*
(1.厦门大学环境与生态学院,福建 厦门361102;2.集美大学水产学院,福建 厦门 361021)
红树林是生长在热带、亚热带海岸潮间带的木本植物群落,它不仅能保护海岸、防风固堤,也为海洋动物、鸟类等提供食物来源和栖息环境,是重要的湿地生态系统之一[1-2].作为位于陆海交界地带的生态系统,红树林的生态环境既受控于海域环境,也与陆域环境关系密切.由于近几十年来沿海地区工业化与城市化的快速发展,红树林正遭受着越来越多的污染问题[3-4].在众多污染物中,重金属污染成为破坏红树林生态环境的重要因素之一[5-6].随着沿海城市人口与经济的增长,流域工农业生产的发展,尤其是沿海城市广泛选择海湾河口作为废物污水排放地,使大量重金属污染物直接或间接进入红树林生态系统中[7].
镉(Cd)虽然不是植物的必需元素,但易于被植物吸收并富集,即使在低浓度水平也会在基因、生化以及生理层面对土壤生物的活性和植物的新陈代谢产生非常不利的影响[8],如:抑制细胞和整个植株生长,导致植株矮小、生物产量下降、干物质减轻等[9-12].红树林能够耐受高浓度的有毒金属[7,13-14],然而富集金属亦可能会导致亚细胞的生物化学变化[8,15].据报道,红树植物秋茄(Kandelia obovata)对Cd具有一定的耐受性,源于细胞壁对Cd的隔离作用[14],根部组织对Cd的吸收和转运也有一定的缓冲作用[12].白骨壤(Avicennia marina)为我国常见红树植物之一,为先锋树种,分布广泛,是组成演替最前阶段的红树群落重要树种之一[1].本文主要探讨Cd污染胁迫对白骨壤幼苗的生长及生理生态两方面的影响,为科学预测与评价Cd对红树植物的生物效应以及红树林湿地的生态环境保护提供科学的依据.
1 材料与方法
1.1 实验材料
成熟白骨壤种苗采于福建九龙江口红树林自然保护区海门岛(24°29′N,117°55′E)的白骨壤林.挑选无病虫害且大小及成熟度相近的种苗用于栽培研究.栽培用砂基经清洗及消毒,等量分装于同样规格的盆栽容器中,砂基高度15cm,直径30cm.培养液用盐度15的人工海水和CdCl2·2.5H2O配制,设置培养液Cd质量浓度分别为0,0.5,5,25,50,100和150mg/L(记为Cd0,Cd0.5,Cd5,Cd25,Cd50,Cd100和 Cd150),每个盆栽加入培养液1 500mL.每个处理组设40颗种苗,3个重复,每个重复为随机选择的12~14颗种苗.所有盆栽放置于玻璃温室中,在自然透光下栽培90d(2007年10月5日—2008年1月3日),期间每7d更换一次培养液,每天用自来水补足散失的水分.
1.2 分析样品制备
抗氧化酶促系统粗酶液制备:在栽培45和90d后分别选取叶片(顶芽下第二对叶)与根尖(1cm左右)500mg,剪碎,置于研钵中,分次加入预冷的0.05 mol/L磷酸缓冲液(pH 7.8,含0.02g/mL的聚乙烯吡咯烷酮)和少量石英砂,冰浴下研磨成匀浆,终体积分别为10mL(叶)与5mL(根尖);离心(4℃,2×104g,20min),取上清液为粗酶液,于4℃冰箱保存,用于可溶性蛋白含量测定和过氧化物酶(POD)、超氧化物歧化酶(SOD)等酶活性测定.
抗氧化非酶促系统提取液制备:分别选取叶片(顶芽下第二对叶)与根尖(1cm左右)500mg左右,剪碎,置于研钵中,分次加入预冷的0.05g/mL的三氯乙酸和少量石英砂,冰浴下研磨成匀浆,终体积为5 mL;离心(4 ℃,2×104g,20min),取上清液为提取液,于4℃冰箱保存,用于抗坏血酸(ASA)及丙二醛(MDA)含量测定.
1.3 分析方法
生长形态指标测定:胁迫栽培15与30d,分别统计各处理组的幼苗萌发率(以第一对叶展开为萌发标准),在栽培过程中观测幼苗的生长状况,对各处理组受害叶片进行计数,并在胁迫栽培45和90d统计各处理组幼苗的成活率(以地上部分枯萎为死亡标准).在栽培45和90d后用直尺测定每株幼苗主根长(统计40根以上)、茎高、叶长和叶宽,叶面积采用剪纸衡重法[16];生物量测定:分别测量植株根、茎和叶各器官的鲜质量及全株总鲜质量;叶绿素含量的测定:每次取下各处理组幼苗的顶芽下第二对叶,去中脉,剪碎,称取100mg左右以混合液法进行叶绿素含量的测定[17];MDA含量的测定:采用硫代巴比妥酸(TBA)显色法进行测定[18];ASA含量的测定参考汤章城[19]的方法;可溶性蛋白含量测定:以牛血清白蛋白为标准蛋白,用考马斯亮蓝G-250染色后,在波长为595 nm处进行比色[20];SOD活性的测定:采用氮蓝四唑(NBT)还原法,以抑制NBT光化学还原的50%为一个酶活性单位U[20];POD活性的测定:采用愈创木酚法,以1min内反应液在470nm处吸光值增加0.01为一个酶活性单位U[21].Cd150处理组的幼苗已有近一半死亡,在45d时采集完用于检测各项指标,因此90d该质量浓度下的各项检测数据缺失.叶面积每个处理组统计所有未受害的健全的第一对叶,统计数为20;其他测试均为3个实验重复,各处理组数据间差异显著性的LSDt检验分析由SPSS 20.0完成.
2 结果与分析
2.1 Cd对白骨壤幼苗生长状况的影响
2.1.1 Cd对白骨壤种苗萌芽及成活率的影响
在Cd胁迫下,白骨壤萌芽仅受一定程度的滞后影响:胁迫栽培15d,不同质量浓度Cd的胁迫使处理组的萌芽率在65%~77.5%之间,显著低于对照组(92.5%)(p<0.05);胁迫栽培30d,各处理组的萌芽率均达到100%.胁迫栽培45d,Cd50、Cd100和Cd150处理组中出现死亡植株,成活率分别为92.5%,75.0%和60.0%;至胁迫栽培90d,Cd胁迫质量浓度≤25mg/L的处理组植株成活率均为100%,而Cd50和Cd100处理组植株的成活率进一步降低,分别为87.5%和57.5%.
这一结果表明:在Cd胁迫下,白骨壤萌芽有一定程度的延迟,成活率随着胁迫质量浓度提高而降低.
2.1.2 Cd对白骨壤幼苗叶生长及子叶宿存率的影响
胁迫栽培19d时,在Cd质量浓度≥25mg/L的处理组,白骨壤幼苗的叶片上出现红褐色近黑色坏死斑点,且叶片受害的数量随着Cd胁迫质量浓度的上升而增加(25~150mg/L).至胁迫栽培45d统计显示Cd25、Cd50、Cd100与Cd150处理组的受害叶片总数 分 别 占 叶 片 总 数 的 10.6%,17.5%,35.0%和52.0%.
白骨壤的叶片大小总体上呈现随Cd胁迫质量浓度的上升而递减的趋势,且胁迫栽培45d的趋势较90 d明显,即后期较为迟缓.Cd对白骨壤幼苗叶长及叶宽影响不明显,各处理组与对照组的差异不显著(p>0.05),在高质量浓度Cd胁迫下,叶形由原来的卵圆盾形逐渐向镰刀形转变.白骨壤在生长过程中,最初提供幼苗生长所需物质与能量的两片子叶会逐渐变黄、萎缩,与茎的结合变松懈,最终凋落与幼苗分离.随Cd胁迫质量浓度增加,子叶凋落进度加快,且随时间推移,此趋势愈发显著;中高质量浓度(25~100 mg/L)处理组的子叶宿存率均不及对照组的40%.
上述结果表明:在Cd胁迫下,白骨壤幼苗叶片面积减小并出现坏死斑点,子叶提早与幼苗分离,且这一趋势随着Cd胁迫质量浓度的增加而加剧.
2.1.3 Cd对白骨壤幼苗茎高与根长的影响
从图1中可以看出,随着Cd胁迫质量浓度的升高,白骨壤的茎高递减,从Cd25处理组开始,与对照组差异显著(p<0.05),但随胁迫时间增加,白骨壤幼苗的茎高降幅减小,如Cd100处理组45d时较对照组降低20.8%,90d时则仅降低14.8%.白骨壤根长有随着Cd胁迫质量浓度的升高而下降的趋势,但对照组与各胁迫组并无显著差异(p>0.05).从Cd25处理组开始,白骨壤须根数目减少,主根根尖呈红褐色,且此现象随Cd质量浓度的升高而愈发显著;但在Cd150处理组也未发现主根有明显死亡的迹象.
这一结果表明:Cd胁迫能抑制白骨壤幼苗茎与主根的生长,该抑制作用对茎的影响较为显著.
2.1.4 Cd对白骨壤幼苗生物量的影响
从图2可以看出,不同胁迫时期,Cd胁迫对白骨壤幼苗各器官及全株生物量均有不同程度的抑制作用.白骨壤幼苗各器官及全株生物量表现出类似的规律:胁迫栽培45d,其生物量总体上表现为随Cd胁迫质量浓度上升而下降;胁迫栽培90d,在0.5mg/L的Cd胁迫下,其生物量相比对照组稍有提高(p>0.05),而后随Cd胁迫质量浓度的上升而减少.
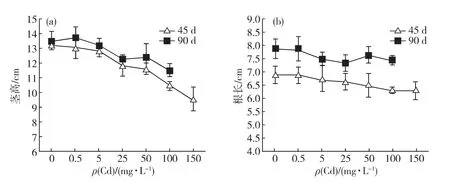
图1 Cd对白骨壤幼苗茎高(a)与根长(b)的影响Fig.1 Effect of Cd on the height of stem (a)and the length of main root(b)of A.marinaseedlings

图2 Cd对白骨壤幼苗生物量生长的影响Fig.2 Effect of Cd on the biomass of A.marinaseedlings
根生物量变化与其他稍有不同.胁迫栽培45d,仅在150mg/L的Cd胁迫下,根生物量与对照组差异显著(p<0.05),相对降低26.6%;而在25~150mg/L的Cd胁迫下,茎、叶及全株的生物量均显著低于对照组(p<0.05).胁迫栽培90d,当Cd胁迫质量浓度≥5mg/L时,根生物量便有显著的降低(p<0.05);而此时茎、叶及全株生物量仅在100mg/L的Cd胁迫下有显著降低(p<0.05).
这一结果表明:不同胁迫时期,Cd对白骨壤各器官生物量均有不同程度的抑制作用.其中,根对Cd的反应在胁迫前期较其他器官迟缓,而到胁迫后期根部所受到的抑制作用较其他器官更为严重.
2.2 Cd对白骨壤幼苗生理生态的影响
2.2.1 Cd对白骨壤幼苗叶绿素含量的影响
如图3所示,胁迫栽培45d,白骨壤幼苗叶片叶绿素a及叶绿素b的含量变化表现基本一致:当Cd胁迫质量浓度≤25mg/L时,叶绿素含量随Cd胁迫质量浓度上升而升高,当胁迫质量浓度继续升高,叶绿素含量则呈下降趋势;而从叶绿素a/b可以看出,叶绿素a与叶绿素b变化幅度相近.胁迫栽培90d,Cd胁迫质量浓度为0.5和5mg/L时,叶绿素a及叶绿素b的含量与对照组相当;Cd胁迫质量浓度≥25mg/L时,叶绿素含量随胁迫质量浓度上升呈下降趋势,且从叶绿素a/b可以看出,叶绿素b降幅大于叶绿素a.
上述结果表明:低质量浓度的Cd(≤25mg/L)在胁迫前期对白骨壤叶绿素的合成具有促进作用,而高质量浓度的Cd会抑制白骨壤叶绿素的合成,且对叶绿素b的抑制作用较叶绿素a大.
2.2.2 Cd对白骨壤幼苗根尖与叶片中 MDA与ASA含量的影响
从图4可以看出,胁迫栽培90d,根尖MDA含量随胁迫质量浓度上升呈下降趋势,从Cd5处理组开始极显著低于对照组(p<0.01).叶的 MDA含量在Cd0.5处理组稍高于对照组,但二者差异并不显著(p>0.05);而后,随胁迫质量浓度上升呈下降趋势且均显著低于对照组(p<0.05).与根尖相比,叶的MDA含量变化幅度较小.如Cd25处理组叶的MDA含量相比对照组降低22.0%,而根尖的MDA含量相对降低57.4%.

图3 Cd对白骨壤幼苗叶片叶绿素含量的影响Fig.3 Effect of Cd on the chlorophyll contents in leaves of A.marinaseedlings

图4 Cd对白骨壤幼苗根尖与叶片中MDA与ASA含量的影响Fig.4 Effect of Cd on the MDA and ASA contents in roots and leaves of A.marinaseedlings
胁迫栽培90d,根尖与叶片的ASA含量总体上随Cd胁迫质量浓度上升而增加,所有胁迫组均显著或极显著高于对照组(p<0.05或0.01).同时根尖的ASA含量变化幅度高于叶片.
上述结果表明:与叶相比,Cd胁迫能较显著地影响根尖的MDA及ASA含量,在0.5~50mg/L的Cd胁迫范围内,能显著减少根尖中MDA含量及提高ASA含量,且此作用与Cd胁迫质量浓度呈一定的正相关.
2.2.3 Cd对白骨壤幼苗根尖与叶片可溶性蛋白的影响
由图5可以看出,胁迫栽培45d,根尖可溶性蛋白含量在0.5~100mg/L的Cd胁迫质量浓度范围内,随着Cd质量浓度增加而上升,显著高于对照组,且在Cd100处理组达到最高值,较对照组提高98.3%;当Cd胁迫质量浓度达到150mg/L时,含量虽略低于Cd100处理组,但仍极显著高于对照组(p<0.01).胁迫栽培90d,根尖可溶性蛋白含量在0.5~25mg/L的Cd胁迫范围内,与对照组相当(p>0.05);当Cd胁迫质量浓度高于25mg/L时,根尖可溶性蛋白含量随胁迫质量浓度上升而增加,但幅度不大,Cd100处理组较对照组仅提高16.6%.
Cd胁迫对叶片可溶性蛋白含量的影响较弱.胁迫栽培45d,叶可溶性蛋白含量随Cd胁迫质量浓度的上升先增加后减少,且在Cd胁迫质量浓度为25mg/L时最高;仅Cd5、Cd25及Cd50处理组与对照组存在显著差异(p<0.05).胁迫栽培90d,叶可溶性蛋白含量除在Cd25处理组有波动外,总体趋势与45d一致.
上述结果表明:Cd胁迫对根尖可溶性蛋白的影响较叶显著,随着Cd胁迫质量浓度的上升,能显著地提升根尖可溶性蛋白含量,而随着胁迫时间的推进,该提升作用减弱.
2.2.4 Cd对白骨壤幼苗根尖与叶片抗氧化酶促系统的影响
如图6所示,胁迫栽培45d,在0~100mg/L的Cd胁迫范围内,根尖SOD活性随Cd胁迫质量浓度上升而增强,Cd150处理组根尖的SOD活性虽开始下降,但仍极显著高于对照组(p<0.01).胁迫栽培90 d,各胁迫组根尖SOD活性虽有所波动,但与对照组无显著差异(p>0.05).
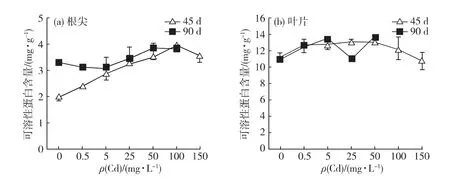
图5 Cd对白骨壤根尖(a)和叶片(b)可溶性蛋白含量的影响Fig.5 Effect of Cd on the soluble protein content in roots(a)and leaves(b)of A.marinaseedlings
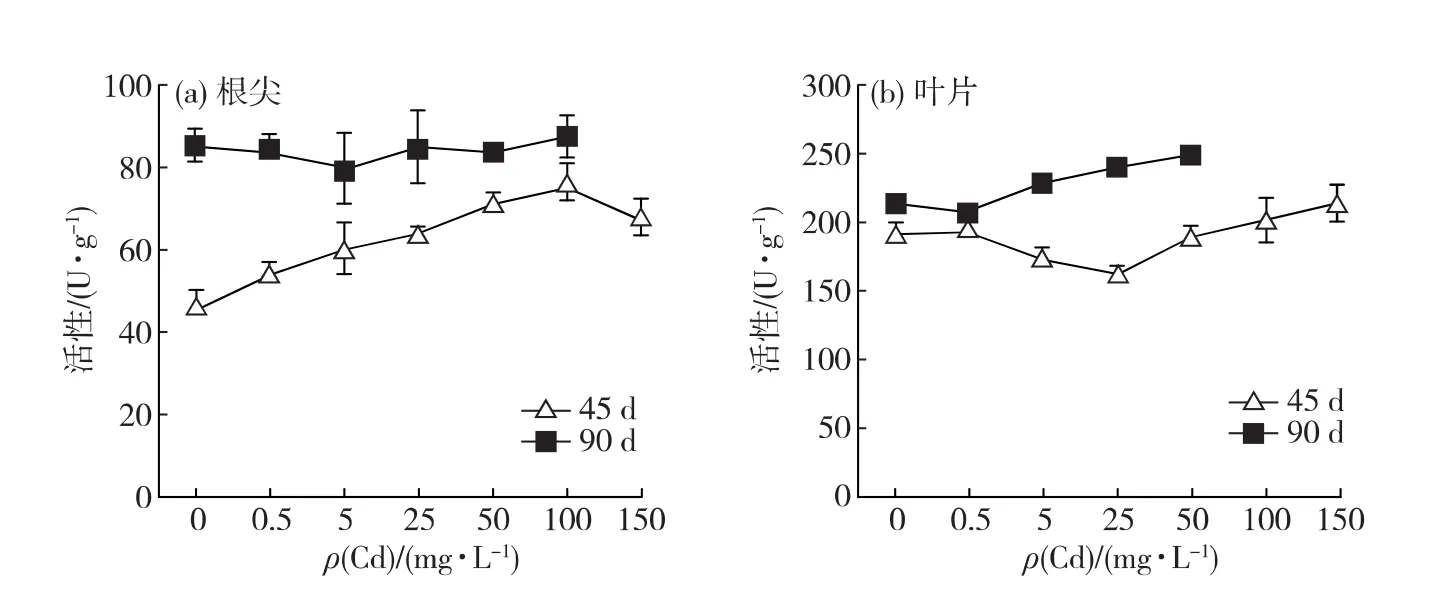
图6 Cd对白骨壤幼苗根尖(a)与叶片(b)SOD活性的影响Fig.6 Effect of Cd on the SOD activity in roots(a)and leaves(b)of A.marinaseedlings
叶片SOD活性随Cd胁迫质量浓度上升先减弱后增强,胁迫栽培45d,Cd25处理组的活性最低且与对照组差异显著(p<0.05).胁迫栽培90d,Cd0.5处理组的叶片SOD活性稍低于对照组,但差异不显著(p>0.05);而后,叶片SOD活性随Cd胁迫质量浓度升高而显著高于对照组并随之增强(p<0.05).
从图7可以看出,根尖POD活性总体上表现为随Cd胁迫质量浓度上升而增强.胁迫栽培45d,根尖POD活性从Cd5处理组开始极显著高于对照组(p<0.01);胁迫栽培90d,根尖POD活性从Cd25处理组开始极显著高于对照组(p<0.01).叶片POD活性变化表现随Cd胁迫质量浓度上升呈先增强后减弱的规律.胁迫栽培45d,叶POD活性在Cd25处理组具最强活性,较对照组增强38.6%,除Cd150处理组与对照组具相当的活性水平外,其余处理组均显著或极显著高于对照组(p<0.05或0.01).胁迫栽培90d,最高活性出现在Cd5处理组,其中Cd5、Cd25与Cd50处理组的POD活性极显著高于对照组(p<0.01).
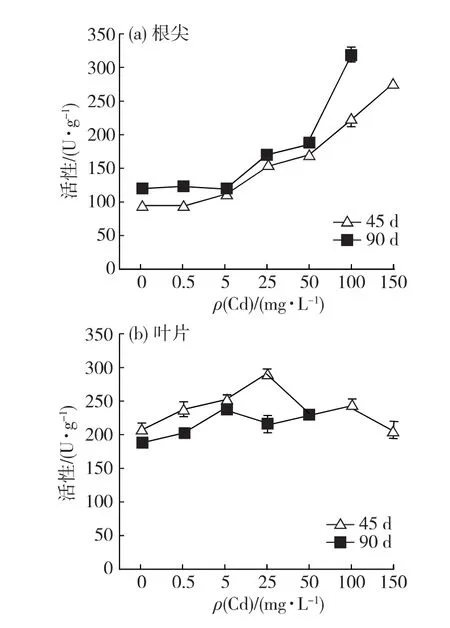
图7 Cd对白骨壤幼苗根尖(a)与叶片(b)POD活性的影响Fig.7 Effect of Cd on the POD activity in roots(a)and leaves(b)of A.marinaseedlings
上述结果表明:胁迫栽培45d,Cd能显著提高白骨壤根尖的SOD和POD活性,到胁迫后期,该促进作用减弱;Cd胁迫对白骨壤叶片的SOD和POD活性也有一定程度的促进作用,但相对于根尖而言,变化幅度较小.
3 讨 论
通过不同质量浓度Cd的胁迫栽培,本文以45和90d为时间节点,研究白骨壤幼苗生长和生理生态所受到的影响及各指标变化的过程.白骨壤幼苗在≥5 mg/L的Cd胁迫下,根生物量虽开始有显著降低,但除细根略有减少外,主根的生长并未受到显著抑制且未出现明显的受害症状,对白骨壤各组分Cd含量进行分析,根尚未有明显的富集Cd(另文发表).
本研究推测,高质量浓度Cd胁迫下白骨壤幼苗叶形的变化是在叶长及叶宽没有明显变化的情况下,叶面积随胁迫质量浓度上升而减小的主要原因.白骨壤幼苗的死亡主要是由于叶片出现坏死斑点,直至叶片全部坏死脱落.MacFarlane等[22]的研究表明,在Zn和Cu污染下,白骨壤叶片在分泌掉体内多余盐分的同时,也可将Zn和Cu排除体外,减少重金属对植物的毒害.本研究在胁迫栽培后期,于中高质量浓度(25~100mg/L)Cd处理组的白骨壤幼苗叶分泌的盐粒收集液中检测出Cd,推测白骨壤借由正常的泌盐活动将Cd排出体外,这尚待进一步研究.
与其他红树植物类似[23-24],白骨壤幼苗在 Cd胁迫下也表现出主根生长、茎高受抑制,各器官生物量减少,叶绿素含量降低等现象.生物量分析显示,Cd胁迫能减缓地上部分生物量的增长,胁迫时间的延长可使抑制作用减弱;相反,Cd胁迫对根生物量的抑制作用随胁迫时间增加而加强,这可能是由于根吸收Cd后,较少向上运输而在根部累积[11],导致Cd对其影响随胁迫时间增加而增强.Cd在根部的含量虽高于其他组分,但未有明显的受害,这可能是因为Cd在根尖中的生物有效性被降低.细胞壁截留Cd在根部、大液泡的分隔以及与细胞配位体的络合反应都可能是白骨壤幼苗降低Cd生物有效性的手段[14],从而减低Cd对原生质的毒害.Cd能够通过抑制叶绿素酸酯还原酶活性、影响氨基-γ-酮戊二酸的合成、直接破坏叶绿素结构及功能并干扰植物对Fe和Zn的吸收和转移,导致叶绿素含量的下降[25].本研究中,胁迫栽培45d,叶绿素a和叶绿素b的含量随Cd胁迫质量浓度的上升均呈现先上升后下降的趋势,这与Cd对桐花树的处理结果类似[23,26],表明低质量浓度的Cd对白骨壤叶绿素的合成有促进作用;但随着胁迫时间的延长,促进作用消失,且Cd对叶绿素b含量的抑制作用增强,不同的是Cd对桐花树叶绿素a的破坏作用更为明显[26].
重金属胁迫和其他形式的氧化胁迫相似,能导致大量的活性氧自由基产生,引起膜脂过氧化.MDA则是膜脂过氧化的产物[4],在植物逆境生理研究中,MDA被普遍地作为一种表示伤害的指标.本研究中,在0.5~50mg/L的Cd胁迫范围内,MDA含量总体上呈下降趋势,表明植物细胞清除自由基能力有所提高.植物的抗氧化系统由酶促和非酶促抗氧化剂构成[24],其中ASA是植物体内普遍存在的一种非酶促类抗氧化剂,可以直接清除O2-、H2O2和·OH等活性氧自由基,在抵御植物体内活性氧损伤方面具有重要的作用.本研究中,胁迫栽培90d,在0.5~50mg/L的Cd胁迫范围内,根尖和叶的ASA含量显著上升,进而提高了白骨壤对Cd胁迫的耐受性.
从可溶性蛋白的分析可以看出,一定质量浓度的Cd胁迫能提高可溶性蛋白在根尖和叶片中的含量,这可能是细胞通过产生植物络合素(PC)等重金属结合多肽或蛋白,结合进入细胞的Cd,以降低Cd的生物有效性,也可能通过产生更多的抗氧化酶来减轻胁迫所带来的自由基对细胞的伤害.SOD和POD就是植物体内活性氧自由基清除系统中重要的保护酶,其活性的提高是一种急性解毒措施,是使细胞免受毒害的调节反应,但其调节能力是临时和有限的,当细胞长时间受到胁迫,细胞内的活性物质包括酶亦会受到损伤而活性下降[27].本研究中,胁迫栽培45d,在5~150mg/L的Cd胁迫下,根尖SOD、POD活性显著提高,但随胁迫时间增加,Cd的促进作用减弱,到胁迫栽培90d,根尖POD活性的变化幅度变小,SOD活性则与对照组相当.Cd胁迫对叶片的SOD、POD活性也有一定促进作用,但相对根尖而言变化幅度较小.
4 结 论
本文通过胁迫栽培,揭示了不同质量浓度的Cd对白骨壤幼苗生长及生理生态的影响,结果表明:Cd胁迫会导致白骨壤幼苗子叶与植株提早分离,叶面积变小,生物量及叶绿素含量减少;白骨壤幼苗对Cd的适宜抗性生长质量浓度范围在25mg/L以下,中高质量浓度Cd胁迫(25~150mg/L)导致叶出现受害坏死斑点,茎高度生长显著受抑制,50~150mg/L的Cd胁迫导致植株死亡;胁迫后期,白骨壤根部的特性使其在Cd胁迫下受害程度较低,中低质量浓度(0.5~50mg/L)Cd胁迫下白骨壤根尖及叶的膜脂过氧化作用有不同程度的减弱,这与白骨壤根尖和叶中ASA含量的上升有关,同时Cd胁迫对白骨壤根尖和叶中的可溶性蛋白含量、SOD及POD活性具有不同程度的促进作用,但随着胁迫时间的增加,促进作用减弱.在自然环境中,污染物往往不是单独存在的,研究复合污染物的胁迫是下一步的研究方向,同时红树植物对重金属的耐受机理以及重金属在环境和植物体内的存在形态还须进一步研究.
[1]林鹏.中国红树林生态系[M].北京:科学出版社,1997:297-316.
[2]Caregnato F F,Koller C E,MacFarlane G R,et al.The glutathione antioxidant system as a biomarker suite for the assessment of heavy metal exposure and effect in the grey mangrove,Avicennia marina (Forsk.)Vierh[J].Marine Pollution Bulletin,2008,56(6):1119-1127.
[3]Mahdavi E S,Rahimi A E K,Amini H V.Pb and Cd accumulation in Avicennia marinafrom Qeshm Island,Persian Gulf[J].Iranian Journal of Fisheries Sciences,2012,11(4):867-875.
[4]Yan Z,Tam N F Y.Differences in lead tolerance between Kandelia obovata and Acanthus ilicifolius seedlings under varying treatment times[J].Aquatic Toxicology,2013,126:154-162.
[5]Bodin N,N′Gom-KâR,KâS,et al.Assessment of trace metal contamination in mangrove ecosystems from Senegal,West Africa[J].Chemosphere,2013,90(2):150-157.
[6]Usman A R A,Alkredaa R S,Al-Wabel M I.Heavy metal contamination in sediments and mangroves from the coast of Red Sea:Avicennia marina as potential metal bioaccumulator[J].Ecotoxicology and Environmental Safety,2013,97:263-270.
[7]郑文教,连玉武,郑逄中,等.广西英罗湾红海榄林重金属元素的累积及动态[J].植物生态学报,1996,20(1):20-27.
[8]Xie X,Weiss D J,Weng B,et al.The short-term effect of cadmium on low molecular weight organic acid and amino acid exudation from mangrove (Kandelia obovata (S.,L.)Yong)roots[J].Environmental Science and Pollution Research,2013,20(2):997-1008.
[9]张金彪,黄维南.镉对植物的生理生态效应的研究进展[J].生态学报,2000,20(3):514-523.
[10]Zhang F Q,Wang Y S,Lou Z P,et al.Effect of heavy metal stress on antioxidative enzymes and lipid peroxidation in leaves and roots of two mangrove plant seedlings(Kandelia candel and Bruguiera gymnorrhiza)[J].Chemosphere,2007,67(1):44-50.
[11]Huang G Y,Wang Y S.Physiological and biochemical responses in the leaves of two mangrove plant seedlings(Kandelia candel and Bruguiera gymnorrhiza)exposed to multiple heavy metals[J].Journal of Hazardous Materials,2010,182(1):848-854.
[12]Du J,Yan C,Li Z.Formation of iron plaque on mangrove Kandalar obovata (S.,L.)root surfaces and its role in cadmium uptake and translocation[J].Marine Pollution Bulletin,2013,74(1):105-109.
[13]Ye J,Yan C,Liu J,et al.Effects of silicon on the distribution of cadmium compartmentation in root tips of Kandelia obovata(S.,L.)Yong[J].Environmental Pollution,2012,162:369-373.
[14]Weng B,Xie X,Weiss D J,et al.Kandelia obovata(S.,L.)Yong tolerance mechanisms to cadmium:subcellular distribution,chemical forms and thiol pools[J].Marine Pollution Bulletin,2012,64(11):2453-2460.
[15]程皓,陈桂珠,叶志鸿.红树林重金属污染生态学研究进展[J].生态学报,2009,29(7):3893-3900.
[16]乔富廉.植物生理学实验分析测定技术[M].北京:中国农业科学技术出版社,2002:78-83.
[17]陈福明,陈顺伟.混合液法测定叶绿素含量的研究[J].林业科技通讯,1984(2):4-8.
[18]刘祖祺,张石城.植物抗性生理学[M].北京:中国农业出版社,1994:371-372.
[19]汤章城.现代植物生理学实验指南[M].北京:科学出版社,1999:356-358.
[20]李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社,2000:167-185.
[21]张志良,瞿伟菁.植物生理学实验指导[M].北京:高等教育出版社,2003:121-124.
[22]MacFarlane G R,Burchett M D.Znic distribution and excretion in the leaves of the grey mangrove,Avicennia marina (Forsk.)Vierh[J].Environmental and Experimental Botany,1999,41(2):167-175.
[23]石贵玉,康浩,段文芳.重金属镉对红树植物白骨壤和桐花树幼苗生理特性的影响[J].广西植物,2009,29(5):644-647.
[24]Weng Z X,Wang L X,Tan F,et al.Proteomic and physiological analyses reveal detoxification and antioxidation induced by Cd stress in Kandelia candel roots[J].Trees,2013,27(3):583-595.
[25]陶毅明,陈燕珍,梁士楚,等.镉胁迫下红树植物木榄幼苗的生理生化特性[J].生态学杂志,2008,27(5):762-766.
[26]杨盛昌,吴琦.Cd对桐花树幼苗生长及某些生理特性的影响[J].海洋环境科学,2003,22(1):38-42.
[27]张利红,李培军,李雪梅.镉胁迫对小麦幼苗生长及生理特性的影响 [J].生态学杂志,2005,24(4):458-460.
