烟草PVY隐性抗病基因的分子标记及其适用性
2015-11-27刘勇宋中邦童治军李永平
刘勇,宋中邦,童治军,李永平
云南省烟草农业科学研究院,烟草行业烟草生物技术育种重点实验室,国家烟草基因工程研究中心 昆明,650021
生物技术
烟草PVY隐性抗病基因的分子标记及其适用性
刘勇,宋中邦,童治军,李永平
云南省烟草农业科学研究院,烟草行业烟草生物技术育种重点实验室,国家烟草基因工程研究中心 昆明,650021
马铃薯Y病毒(Potato virus Y,PVY)是危害烟草的重要病害,种植抗病品种是经济有效的防治措施。分子标记辅助选择可提高抗病育种效率。来源于Χ-射线诱变的Virgin A Mutante(VAM)的隐性抗PVY基因位点(va)被广泛应用于烟草抗病育种。为了提高va位点的育种利用效率,根据烟草隐性抗PVY基因(感病基因)eIF4E-1基因序列,设计特异扩增的引物CF2GR11,开发eIF4E-1基因的分子标记,并检测了该标记与抗性的遗传距离和在常见烟草资源中的适用性。CF2GR11在云烟87、红花大金元和K326等感PVY品种可扩增出500 bp产物,在NC102、NC55和K326PVY等抗病品种无扩增条带。以抗、感PVY亲本构建的101个F2单株为定位群体,遗传连锁分析表明,CF2GR11标记与烟草PVY感病基因的遗传距离为0.99 cM。对46份PVY抗性明确的栽培烟草资源的检测表明,供试资源的标记检测结果与抗性的吻合度为100%,表明CF2GR11标记适用性高。该目的基因标记可用于抗PVY育种的辅助选择和抗PVY资源的鉴定。
烟草;马铃薯Y病毒;抗性;分子标记
烟草马铃薯 Y病毒病,又称为脉斑病,是由马铃薯Y病毒(Potato virus Y,PVY)引起的蚜传病毒病,在中国北方烟区危害严重,在南方烟区的危害呈上升趋势[1-3]。缺乏抗病主栽烟草品种是导致PVY危害严重的主要原因之一[4]。为选育抗病烟草品种,国内外鉴定出多个抗PVY的种质资源,如VAM(TI1406)、V.SCR等,并育成抗病白肋烟品种TN86、TN90和烤烟品种NC55、NC102等,并对几个抗源的抗性遗传特性、抗性相关的分子标记进行了研究[5-7]。大部分PVY抗源的抗性表现为隐性基因位点(va)控制。与PVY抗性相关的分子标记,如Randomly Ampli fi ed Polymorphic DNA(RAPD)和Sequence Characterized Ampli fi ed Region(SCAR)标记也有报道[8-10]。现有分子标记与抗性的遗传距离相对较远。Noguchis等[8](1999)认为va基因型烟草植株的抗性是由于对PVY感病的基因片段的缺失造成的,Chikara等[11](1999)认为VAM的抗性机理为抑制病毒粒子细胞间移动和胞内复制。云南省烟草农业科学院克隆了va基因位点的一个隐性抗PVY基因eIF4E-1(待发表),也就是感PVY基因,该基因属于真核生物翻译起始因子(Eucaryotic initiation factor,简写为eIF)的一种类型。利用该抗病基因序列开发紧密连锁的特异分子标记,可为抗病育种提供新的标记。本文根据烟草eIF4E-1基因及其家族基因的序列信息,开发出一个新的与PVY抗性紧密连锁的分子标记,并检验了该标记在部分品种和种质资源中的适用性。
1 材料和方法
1.1 植物材料
抗PVY烤烟种质NC55和感PVY烤烟种质Coker176为云南省烟草农业科学院保存。以抗病亲本(PR)和感病亲本(Ps)杂交得到的F2群体,用于分子标记与抗性的遗传距离分析。该群体的抗性数据和SCAR标记分型数据来源于文献[10]。验证标记适用性的烟草资源,包括常用烤烟品种见表2,PVY抗性根据文献确定,为云南省烟草农业科学院保存。
1.2 抗性鉴定
PVY坏死株系分离物ZT-5由云南省烟草农业科学研究院分离,在防虫网室内的烟草种质Samsun NN繁殖备用。防虫塑料大棚内常规方法漂浮育苗,烟苗第1次剪叶后(播种后40~45 d)移栽至直径20 cm的花盆,成活后采用高压喷枪摩擦接种[12],接种浓度为病叶汁液稀释200倍。烟苗接种后,每7 d左右调查发病情况,连续调查3次,记载单株是否表现PVY症状。挑选抗性明确的单株用于基因型分型。抗性划分标准为:3次调查均无症状,划分为抗病;第1次或第2次调查表现PVY症状,划分为感病。
1.3 烟草总DNA 的提取与质量检测
取烟草新鲜嫩叶,-80℃保存。F2群体样品采用CTAB法[13]提取烟草总基因组DNA。种质资源样品采用QIAGEN DNeasy Plant Mini试剂盒提取烟草总基因组DNA。采用紫外分光光度法(Nanodrop)和琼脂糖凝胶电泳法初步检测DNA质量,去掉DNA浓度低于8 ng/μL的样品。质量合格的DNA样品,用0.5×TE溶液稀释至30~50 ng/μL,保存备用。采用扩增烟草内参基因Actin检测DNA是否可用于PCR扩增。淘汰Actin基因扩增阴性的单株DNA。挑选Actin基因扩增阳性的单株DNA用于CF2GR11引物扩增。
1.4 标记引物设计
根据烟草eIF4E-1基因序列和eIF4E家族成员的序列,设计特异引物对CF2:5’-TTTGGTTTGATAATCCTATGGCT -3’,GR11:5’-GAAGGCAAGATATTCAGGAGCT-3’; 扩增片段大小为446bp,退火温度51℃。CF2GR3扩增片段大小为1800bp,退火温度55℃。扩增烟草内参基因Actin的特异引物为Actin-F:5′-AAGGGATGCGAGGATGGA-3′,Actin-R 5′-CAAGGAAATCACCGCTTTGG-3′,扩增片段大小为160bp,退火温度58℃。引物由大连宝生物公司合成。
1.5 PCR 扩增和电泳检测
PCR 反应体系总体积均为20μL,其中30~50 ng/μL DNA 样品 2.5μL、10×PCR buffer 2.0μL,dNTPs 1.2μL,引 物 各 1.5μL,rTaq DNA 酶 0.3μL,ddH2O 12.6μL。所用试剂购自宝生物公司。PCR反应在Ependorff梯度扩增仪上(Master Cycler )上进行。扩增的程序为:1)94℃预变性5 min;94℃变性30 s,退火温度退火30 s,72℃延伸1 min,共28个循环,72℃延伸10 min,4℃保存;采用2%的琼脂糖凝胶进行检测。
1.6 数据统计与分析
PCR扩增的带型统计方法参考文献[14,15]。对首次CF2GR11扩增结果显示为交换单株的样品,则重复扩增1-2次,以验证交换单株的结果。保留2-3次扩增中有两次结果一致的样品数据,用于标记的遗传距离计算。卡方检验采用SPSS 12.0 软件。遗传图距(D)的计算方法为:(1)计算交换值,交换值(r)=(交换配子数/总配子数)×100%;(2) 根据Kosambi 函数计算遗传图距[16](D,单位cM),D=0.25×ln((1+2r)/(1-2r))×100。
2 结果与分析
2.1 候选分子标记的确定
根据eIF4E-1的基因组序列和eIF4E家族成员的序列,设计特异引物CF2GR11和CF2GR3。检测已知抗性的烟草材料的基因组DNA,验证抗性是否与eIF4E-1的目标片段吻合。结果表明:感病品种K326、云烟87和红花大金元的eIF4E-1目标片段检测为阳性,而抗病品种NC102、NC55、K326PVY的eIF4E-1目标片段检测为阴性(图1)。6个品种对PVY的抗性与CF2GR11的PCR检测结果吻合。引物CF2GR3在感病品种云烟87中多次扩增结果为阴性(未列出数据)。因此,将CF2GR11特异扩增的eIF4E-1基因片段作为候选分子标记,用于在抗性分离群体中验证。

图1 引物CF2GR11在抗感品种间的多态性Fig.1 Polymorphism of primer CF2GR11 among resistance cultivars and susceptible cultivars
2.2 标记与抗性的遗传距离分析
对抗病品种NC55与感病品种Coker176配制的F2群体进行抗性鉴定与基因分型。前期结果表明,该群体苗期接种后21 d的统计结果显示,在309个单株中,感病株和抗病株的实际比例为230:79,理论分离比为232:77。卡方检验表明该群体代的实测分离值与理论分离值无显著差异性(P=0.8696)[10]。根据DNA的浓度与内参Actin基因的扩增结果,挑选出42份感病单株DNA和59份抗病单株DNA,用于CF2GR11基因分型。对感病单株第1次扩增阴性的样品,重复扩增1次。将2次扩增结果一致的样品数据作为有效数据,用于遗传距离分析。获得F2代遗传群体101个单株的CF2GR11扩增的有效数据(表1)。利用3.6中的公式计算重组率及遗传距离,结果表明标记CF2GR11与PVY感病性的遗传距离为0.99 cM(图 2 )。

表1 标记与抗性在F2群体中的独立性检验Tab.1 Independence test between marker and resistance in the F2 population
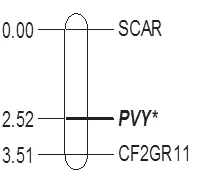
图2 基于CF2GR11标记的Va基因位点的遗传连锁图(cM)Fig.2 Genetic linkage map of Va locus based on SCAR markers(cM)
2.3 标记在抗感PVY烟草资源中的适用性
根据文献挑选对PVY抗性明确的栽培烟草资源46份,检测CF2GR11标记的适用性。结果表明(表2)。NC102、NC55和TN86等9个抗病品种的标记检测为阴性。云烟87和K326等37个感病品种的CF2GR11的标记检测为阳性,供试品种的标记检测与抗性的吻合度为100%。表明CF2GR11标记在供试烟草资源中的适用性高,可用于转育va基因位点抗性的辅助选择。
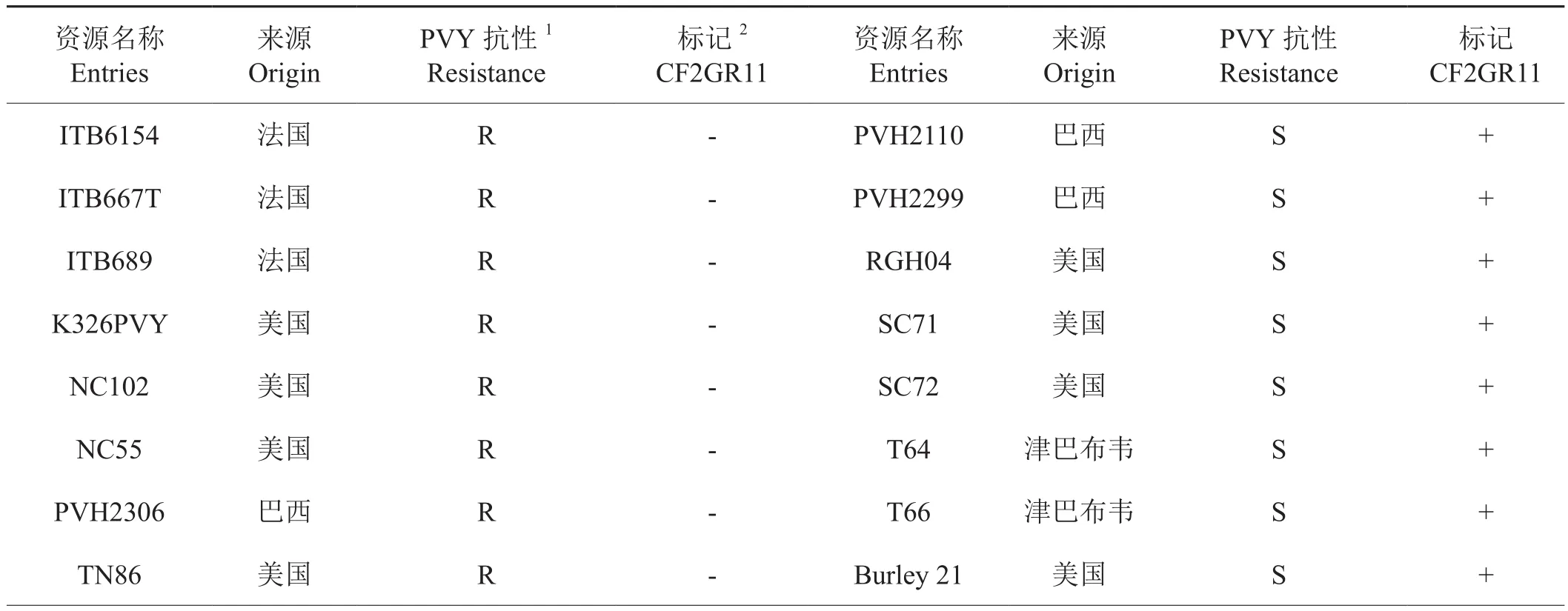
表2 CF2GR11标记检测与PVY抗性比较Tab.2 CF2GR11 marker detection compared to PVY resistance of germplasm
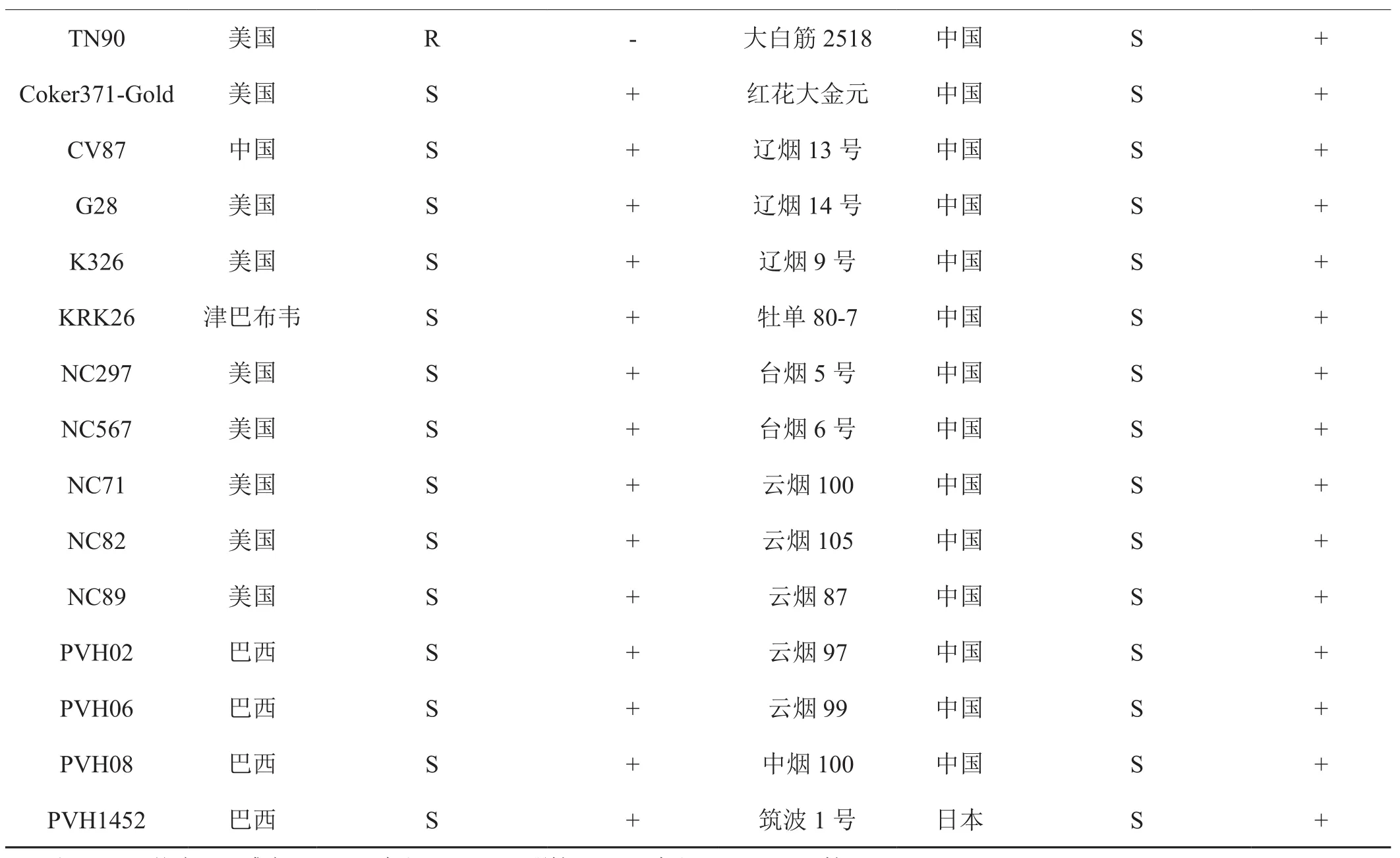
续表2
3 讨论
3.1 基于eIF4E-1基因的标记设计
前期研究表明eIF4E-1缺失可赋予烟草对PVY的抗性。栽培烟草为异源四倍体,eIF4E-1的家族成员较多,序列相似率高。设计特异扩增与PVY抗性相关的eIF4E-1分子标记难度较大。eIF4E-1的基因组大小约5.3kb。本文通过在eIF4E-1基因的第1个外显子内设计上游引物,在内含子区域设计下游引物,获得在抗病品种中无扩增条带,而在感病品种中有特异条带的引物对。
3.2 与PVY抗性连锁的分子标记
Noguchi等[8]对只在PVY抗性上有差异的近等基因系(PVY抗性来源于Perevi)及其F2代进行RAPD分析,找到10个与Va连锁的RAPD标记,这些标记在8个感病品种中都存在,至少有1个标记在8个抗病品种中不存在。Tajima等[14]将这些RAPD标记转化为STS标记,成功用于回交后代的育种选择。Julio等[9](2006)将PVY抗性相关的AFLP标记,转化成SCAR标记,找到与TN86中va的等位基因Va连锁标记PVYME1(遗传距离为5.1cM)。王贵等[10]检测了PVY抗源NC55中RAPD标记和SCAR标记的存在情况,获得了两个与NC55的PVY抗病对应的等位基因Va连锁的标记O12V3695和PVYME1,与Va的遗传距离分别为2.10 cM和2.52Cm。上述分子标记都是通过抗性分离群体的大量差异标记分型获得的。标记与抗性的连锁距离相对较远。本文根据烟草资源NC55等的隐性抗病基因的序列开发分子标记CF2GR11,该标记与抗性紧密连锁,遗传距离为0.99cM。
3.3 CF2GR11标记在PVY抗源类型划分中的潜力
根据现有文献,烟草抗PVY的抗源主要有两大类:一类是以VAM为代表的隐性基因位点(va)控制的抗性,包括Perevi、Kerti No.1、PBD6、Wislica、NC744、NC745、TN86、TN90等种质[8-9]。另一类是以野生种N.africana为代表的对PVY免疫的抗性,包括N.africana的衍生种质NC152、K326/Naf等[17]。目前育种利用的抗源主要为va基因位点。Acosta-Leal等[18]提出VAM对PVYN的抗性由两对隐性基因va1和va2控制,va1限制病毒细胞间运动和阻断病毒进入维管束,va2抑制病毒细胞内积累。利用本文的CF2GR11可初步划分烟草PVY抗性的类型。表型鉴定为抗PVY的资源,若CF2GR11标记阴性,则表明该资源的PVY抗性与eIF4E-1基因缺失有关,属于隐性基因位点(va)控制的抗性;若CF2GR11标记阳性,则表明该资源的PVY抗性与eIF4E-1基因缺失无关,属于eIF4E-1基因突变或其他抗病机制。通过简单快速的标记划分抗性类型,有助于发现新的烟草PVY抗源。
3.3 CF2GR11标记的适用性
栽培烟草的基因组约4.5G,具有巨大而且复杂等特点。根据少数品种验证的分子标记,在多大范围内的种质资源适用,需要实验验证。本文结果表明,CF2GR11标记在供试46个品种资源中都适用,包括常见va基因座的抗病资源与常见烟草品种。可利用该标记辅助选择eIF4E-1基因介导的PVY抗性。
3.4 结论
根据烟草抗PVY的隐性基因开发的分子标记未见其他文献报道,CF2GR11标记与va基因位点控制的抗性紧密连锁,检测的稳定性高,在常见烟草种质资源中的适用性高。在抗PVY育种的辅助选择和抗病资源鉴定中具有较大的应用前景。
致谢:感谢杨华兵协助完成遗传群体抗性鉴定、黄昌军博士提供数据分析支持。
[1]李淑君,王海涛,陈玉国,等.2000年烟草病毒病大发生概况及原因分析[J].烟草科技,2001(1):44-46.
[2]李义强,王凤龙,程云吉,等.黄淮烟区烟草病毒病发生种类、发病规律研究,中国烟叶学术论文集,科学技术文献出版社,北京:2004:438-442.
[3]刘勇,莫笑晗,余清,等.云南、福建、湖南烟区烟草花叶病主要病毒种类检测及黄瓜花叶病毒亚组鉴定 [J].植物病理学报,2006,36(4):310-313.
[4]Burk L.G.,Gooding G.V.,and Chaplin J.F.Reaction of Nicotiana species and cultivars or breeding lines of Nicotiana tabacum to three strains of potato virus Y[J].Tobacco Science,1982,26:85-88.
[5]陈荣平,焦庆明,兰荣利,等.烟草抗PVY育种材料的筛选与应用[J].中国烟草科学,2000(1):1-4.
[6]周显升,钱玉梅,陈德鑫,等.烟草品种对马铃薯Y病毒的抗性遗传分析[J].中国烟草学报[J].2005,13(1):31-36.
[7]陈万胜.烟草马铃薯Y病毒病抗性遗传及分子标记[D],中国农业科学院研究生院,2008.
[8]Noguchi S,Tajima T,Yamamoto Y,et al.Deletion of a large genomic segment in tobacco varieties that are resistant to potato virus Y(PVY) [J].Mol Gen Genet,1999,262:822-829.
[9]Julio E,Denoyes-Rothan B,Verrier JL,et al.Detection of QTLs linked to leaf and smoke properties in Nicotiana tabacum based on a study of 114 recombinant inbred lines[J].Mol Breeding,2006,14:114-123.
[10]王贵,刘勇,卢秀萍,等.烟草PVY抗性的遗传分析与分子标记筛选[J].分子植物育种,2012,10(1):97-103.
[11]Chikara M,Hiroshi M,Tatsuji H,et al.A single amino acid change in viral genome-associated protein of potato virus Y correlates with resistance breeding in “Virgin A Mutant”tobacco[J].Phytopathology,1999,89(2):118-123.
[12]林志文,刘勇,李梅云,等.烟草种质资源抗马铃薯Y病毒病鉴定方法比较[J].中国农学通报2010,26(19):269-274.
[13]Murry HG,Thomspon WF.Rapid isolation of weight DNA[J].Nucleic Acids Res,1980,8:4321-4322.
[14]Tajima T,Noguchi S,Tanoue W,et al.Background selection using DNA markers in backcross breeding program for potato virus Y resistance of tobacco [J].Breeding Science,2002,52:253-257.
[15]肖炳光,高玉龙,吴为人.23 份烟草品种遗传关系的SSR 分析[J].分子植物育种(网络版),2011,4(9) :1297-1304.
[16]Kosambi D D.The estimation of map distances from recombination values[J].Ann Eugen ,1944,12:172-175.
[17]Lewis R S.Transfer of resistance to potato virus Y(PVY)from Nicotiana Africana to Nicotiana tabacum:possible influence of tissue culture on the rate of introgression[J].Theor Appl Genet,2005,110:678-687.
[18]Acosta-Leal R,Χiong Z.Complementary functions of two recessive R-genes determine resistance durability of tobacco'Virgin A Mutant'(VAM) to Potato virus Y[J].Virology,2008,379(2):275-283.
Molecular marker from recessive gene resistant to potato virus Y of tobacco and its suitability
LIU Yong,SONG Zhongbang,TONG Zhijun,LI Yongping
Yunnan Academy of Tobacco Agricultural Sciences,Key Laboratory of Tobacco Biotechnological Breeding,National Tobacco Genetic Engineering Research Center,Kunming,650021,China
Potato virus Y(PVY) is a serious tobacco disease.Molecular marker-assisted selection could improve breeding efficiency.A gene locusvaof Virgin A Mutante(VAM) induced by Χ-ray irradiation is widely used in tobacco breeding.In order to improve va locus breeding utilization ef fi ciency,speci fi c ampli fi cation primer CF2GR11 was developed based on recessive resistance gene eIF4E-1(susceptible gene) to tobacco PVY,and genetic distance between CF2GR11 and resistance gene,as well as its suitability of CF2GR11 in tobacco germplasm were evaluated.In PVY susceptible varieties Yunyan87,Honghuadajinyuan and K326,CF2GR11 can produce a 446bp product as ampli fi er,while no product in PVY resistant varieties NC102,NC55 and K326PVY.Genetic linkage analysis of F2 population from PVY resistant and susceptible parents as locus point showed that genetic distance between CF2GR11 marker and resistance gene was 0.99cM.CF2GR11 proved suitable in 46 tested tobacco germplasm.This target gene marker could be used as an assisted selection tool for PVY resistant variety breeding and germplasm identi fi cation.
tobacco; potato virus Y; resistance; molecular marker
刘勇,宋中邦,童治军,等.烟草PVY隐性抗病基因的分子标记及其适用性[J].中国烟草学报,2015,21(1)
中国烟草总公司云南省公司科技项目(2012YN02);中国烟草总公司科技项目(110201301010)
刘勇(1970—),博士,副研究员,主要从事烟草病害防治与抗病育种研究,Tel:0871-65106352,Email:yliu@yntsti.com
李永平(1966—),硕士,研究员,主要从事烟草育种研究,Email:liyongping@yntsti.com
2014-03-28
:LIU Yong,SONG Zhongbang,TONG Zhijun,et al.Molecular marker from recessive gene resistant to potato virus Y of tobacco and its suitability [J].Acta Tabacaria Sinica,2015,21(1)
