海洋酸化对鱼类感觉和行为影响的研究进展
2015-10-09王晓杰宋佳坤范纯新张旭光郭弘艺
王晓杰,宋佳坤,范纯新,张旭光,郭弘艺
1. 上海海洋大学海洋生物系统与神经科学研究所,上海 201306 2. 上海海洋大学国际海洋研究中心,上海 201306 3. 上海海洋大学省部共建水产种质资源发掘与利用教育部重点实验室,上海 201306
海洋酸化对鱼类感觉和行为影响的研究进展
王晓杰1,2,3,*,宋佳坤1,2,3,范纯新1,2,3,张旭光1,2,3,郭弘艺1,2,3
1. 上海海洋大学海洋生物系统与神经科学研究所,上海 201306 2. 上海海洋大学国际海洋研究中心,上海 201306 3. 上海海洋大学省部共建水产种质资源发掘与利用教育部重点实验室,上海 201306
海洋酸化是指大气增多的二氧化碳(CO2)溶解于海水而导致海水pH值降低的过程。海洋酸化将改变海水碳酸盐平衡体系,使依赖于原化学环境的多种海洋生物乃至生态系统面临巨大威胁。海洋酸化对钙质生物影响的研究最早引起大家关注,而海洋鱼类具有较完善的酸碱调节机制,大家普遍认为酸化对其影响不大。但在过去的5年中,不少实验证明海洋酸化会影响海洋鱼类仔稚鱼的感觉和行为,减弱其野外的生存能力及增加被捕食率,很可能将威胁自然种群补给量和影响全球的渔业资源量。本文从嗅觉、听觉、视觉及高级意识和相关行为角度,综述近几年海洋酸化对鱼类感觉和行为影响的研究进展,介绍了鱼类神经行为生物学的研究,为全面了解和预测海洋酸化的生态、经济和社会效应提供科学依据。
海洋酸化;海洋鱼类;感觉系统;行为;综述
海洋酸化指的是海水溶解更多的大气二氧化碳(CO2)而导致的海水pH值降低的过程。自工业革命以来,由人类活动产生CO2总量的1/3被海洋吸收,导致海洋快速酸化,表层海水pH值已经降低了0.1。如果按照目前化石燃料消耗量和大气CO2浓度升高的趋势发展(目前浓度是390 ppm),到21世纪末CO2浓度会增加至720~1 020 ppm[1],pH值可能会下降0.3~0.4[2]。这就会降低海水中碳酸钙的饱和度,使珊瑚虫、双壳类等钙化生物难以形成钙质骨骼[3-5]。
海洋酸化对钙质生物影响的研究最早引起大家关注,海洋鱼类具有较完善的酸碱调节机制,大家普遍认为酸化对其影响不大。与体内脏器不同,鱼类的嗅觉、味觉、视觉和听觉感受器均位于鱼体表,直接与海水接触。即使鱼类具有较强的酸碱调节能力,感觉系统也会直接面对海洋环境的变化。研究表明:感觉细胞膜上分布着大量电压门控离子通道[6-7],外界环境的离子改变可能会影响这些电压门控离子通道的活性。在过去的5年中,大量实验也证明酸化海水(CO2浓度为预计到21世纪末海洋中所能达到的浓度),会影响海洋鱼类仔稚鱼的感觉和行为,由此增加其在野外的死亡率及被捕食率,影响自然鱼类种类多样性和种群补给量。在全球渔业资源总产量中,鱼类发挥最重要作用,其捕获量占总量的85%,因此海洋酸化对鱼类资源的潜在影响将严重影响全球的渔业产量[8]。而且,海洋酸化对鱼类的影响还可能通过海洋中复杂的食物网,最终影响整个海洋生态系统的服务与产出功能[9]。
为此,本文综述海洋酸化对鱼类嗅觉、听觉、视觉及高级意识和行为影响的近几年的研究进展,从鱼类神经行为生物学的角度,为全面了解和预测海洋酸化对海洋鱼类及整个生态系统的影响提供基础。
1 海洋酸化对鱼类嗅觉相关行为的影响(Impacts of ocean acidification on olfactory system and its related behaviour in marine fish )
扰乱水生生物嗅觉系统及化学感受系统的功能,是海洋酸化引起的一个主要的不利影响。在鱼类行为决策形成过程中,尤其是在环境复杂、水体浑浊或者低光照情况下,嗅觉和味觉感受器对化学因子和化学信号的感知发挥主要作用[10-11]。鱼类的诸多行为,包括归巢行为、微生境选择、食物定位、配偶选择以及避敌行为等,都需要或部分需要正确探知周围水环境中的化学信号。
大部分近岸鱼类在浮游生活阶段结束后能否找到适宜栖息地定居,是决定其种群能否维持的重要条件。在外海经过浮游生活阶段,珊瑚礁鱼类幼鱼在靠近近岸珊瑚礁后,通过辨别周围水体中化学信号找到适宜的栖息地。已有室内研究发现,海洋酸化能够影响珊瑚礁鱼类对栖息地气味的嗅觉辨别力。小丑鱼(Amphiprion percula)喜欢生活在有植物生长的海岛周围的珊瑚礁区域。当对水体CO2加富酸化(1 000 ppm, pH7.8),显著影响了发育至定居阶段小丑鱼仔鱼的嗅觉辨别能力,幼鱼被正常情况下回避的植物叶子的气味强烈的吸引[12]。在对珊瑚礁天竺鲷(Cheilodipterus quinquelineatus)成鱼研究中发现,对照组鱼在对本地同种鱼栖息地气味和外地同种栖息地气味的选择实验中,偏好前者;酸化(550 ppm、700 ppm、950 ppm)处理4 d后,实验鱼失去了对本地同种鱼栖息地气味的偏好[13]。同样在对虾虎鱼(Paragobiodon xanthosomus)成鱼的酸化实验中也发现,处理组鱼失去了对正常情况下喜欢的珊瑚种类气味的偏好[14]。
在自然栖息地进行野外实验,进一步观察酸化对鱼类嗅觉行为影响带来的生态后果。经过酸化处理后,将天竺鲷(C. quinquelineatus)成鱼放置在距离其白天休息地200 m远的海域,3 d后,对照组有62%鱼回到自己的栖息地,而酸化处理组(550 ppm、700 ppm、950 ppm)有40%,40%和32%的鱼回到栖息地。而且观察酸化处理后,实验鱼回到自然栖息地白天活动率明显高于对照区,且远离栖息地的探险行为也更为明显[15]。酸化处理虾虎鱼(P. xanthosomus),然后将其放置在海区死亡珊瑚礁块上,观察在24 h内虾虎鱼对栖息地选择情况。对照组鱼全部离开了死亡珊瑚礁块,转移到喜欢的活珊瑚礁上,而处理组或者仍留在放置地,或者移到正常不偏好的珊瑚礁上[14]。
探知和避让敌害能力是保证鱼类存活的一个重要机制,尤其是在仔稚鱼阶段[16]。而在海水生态系统中,对敌害的识别和避让主要依赖对敌害化学气味的探测[17-18]。小丑鱼(A. percula)幼鱼能够利用嗅觉信号探测敌害,并能分辨敌害生物和非敌害生物产生的化学信号的异同。但当海水酸化,幼鱼失去了对敌害生物的嗅觉辨别能力,甚至产生与敌害避让行为完全相反的行为[19]。具有重要经济价值和生态价值的珊瑚礁鳟鱼(Plectropomus leopardus)稚鱼,在酸化水体中养殖4 d或4周后,与对照组相比,均表现出明显被敌害生物的气味所吸引,并且活动性增强,呆在庇护处的时间缩短,更加大胆[20]。海洋酸化对鱼类捕食者-猎物相互关系的影响,将会增加仔稚鱼的死亡率,导致种群补给量的下降。
2 海洋酸化对鱼类内耳耳石及听力的影响(Impacts of ocean acidification on the otoliths and the hearing ability of marine fish)
海水鱼类的内耳中有石灰质的耳石,已有研究表明海洋酸化对耳石的形状、密度、日增长等产生影响,并进而可能影响听觉敏感度以及听觉相关行为。在高CO2分压在993到2 558 μatm酸化海水处理,多种海水鱼的仔稚鱼耳石的大小、表面积、密度以及日增长显著增加[21-25],但其影响机制还不清楚,可能是在高CO2分压的海水中鱼体进行酸碱调节,使得内耳淋巴液中碳酸钙饱和度增加有关[21]。内耳中的耳石与内耳的感觉毛细胞相接触,当外界声音传至鱼内耳后,内淋巴液振荡,这些振荡会震动耳石和刺激毛细胞,将信号通过听神经传达到脑,产生听觉。因此当耳石的大小、密度和质量等变化将会直接影响鱼类的听力及听觉功能。利用数值模型推算在一定水流下耳石的运动,发现随着耳石的增大,军曹鱼(Rachycentron canadum)仔鱼耳石与感觉毛细胞间的相对运动增加,即鱼的听力敏感度增加,使得在高CO2分压水体中的鱼能听到在低CO2分压水体中所听不到的声音,但听力改变对鱼类生存是利或是弊很难判断[25]。当利用听觉信息判断远处猎物时,敏感度增加是有利的,但是,敏感度的增加同时可能使鱼类获得更多的背景噪音,因此会影响鱼类从大量听觉信息中对有用信息的提取。也有研究发现在低CO2分压(600~900 μatm)酸化实验中,小丑鱼稚鱼耳石的形态和成分没有明显变化,但是其与听觉相关的行为却受到影响,对照组鱼对珊瑚礁噪音有明显的避让,但是酸化处理组实验鱼对噪音并不表现明显避让[22]。
3 海洋酸化对鱼类视觉及相关行为影响(Impacts of ocean acidification on visual system and its related behaviour in marine fish)
视觉系统是动物捕捉猎物和逃避敌害的最有效的感觉系统之一,海洋酸化对鱼类的视觉生理及视觉相关的行为也产生不利影响。酸化影响安邦雀鲷(Pomacentrus amboinensis)的视网膜反应,通过检测闪烁视网膜电图(flicker electroretinogram, fERG)阈值,发现海水酸化降低视网膜最大闪烁频率,可能降低鱼体对快速事件的反应能力[26]。850 μatm CO2酸化处理后,再给予雀鲷稚鱼的捕食者多刺光鳃鱼(Acanthochromis polyancanthus)的视觉刺激,与对照组相比,雀鲷降低了抵抗捕食者的反应,不但没有表现出觅食行为、活动性以及区域利用的下降,而且它们更靠近其捕食者,缺乏在危险情况下雀鲷典型的上下左右来回游动(bobbing)行为[27]。
酸化对鱼类不同感觉系统影响的程度不同,通过多感觉系统之间的调节和补偿,最终降低酸化对鱼体感觉系统的不利影响[13]。海洋鱼类在近岸珊瑚礁定居后,主要依赖视觉和化学感觉获取信息。在夜晚及复杂环境中,化学信息因子包括捕食者气味和同种的警戒素发挥重要作用,但是视觉可以提供危险的时空方面最准确的信息[28]。酸化能够使得雀鲷稚鱼完全失去对同种警告信号的反应,并部分降低其对捕食者的视觉反应。当给予多重捕食者信号(化学和视觉)时,酸化处理的鱼比对照组呆在遮挡物下的时间短,没有表现出危险情况下的游动行为,但是,他们确实降低了摄食率和活动水平,表明酸化情况下,视觉反应可以部分补偿化学反应的缺失[29]。
4 海洋酸化对鱼类高级认知及焦虑的影响(Impacts of ocean acidification on the advanced consciousness and anxiety in marine fish)
越来越多的研究表明,海洋酸化不仅影响海洋鱼类仔稚鱼的嗅觉、听觉和视觉,还对其大脑功能以及认知行为等产生影响。
行为侧向化是大脑功能非对称性的一种表现,在复杂情况下利于缩短决策时间,与逃跑反应[30]、空间定位[31]、集群[32]等行为有关。在热带鱼黄尾新雀鲷(Neopomacentrus azysron)和温水性三棘刺鱼(Gasterosteus aculeatus)行为侧向化实验中,用T型迷宫水槽观察实验鱼向左或向右转弯的喜好,对照组表现出明显的向左或向右的喜好性,但经过酸化处理的仔鱼,行为侧向化受到影响,实验鱼不表现出方向偏好[33-34]。学习是动物的一种高级认知,对于动物获得新知识、技巧、行为以及交流等都很重要,其中与捕食者之间的经历是识别捕食者的最有效方法。对于自然界中的水生生物,通过2种学习方式识别捕食者,一种是通过化学感觉探知水体中同物种受伤气味[28],另一种是通过社交学习,即初学者观察学习有经验的同种应对捕食者过程[35-36]。研究发现,850 μatm CO2分压的酸化海水处理,对雀鲷幼鱼的化学感觉和视觉识别捕食者的学习能力都有负影响[37],然后将学习能力受损的鱼放置野外珊瑚礁区,发现其死亡率比对照组高2.5~3倍[38]。利用光暗偏好以及物体靠近的行为实验评估加利福尼亚鲉(Sebastes diploproa)的焦虑程度的结果表明,经1 125 μatm CO2分压酸化处理1周,实验鱼的焦虑程度显著高于对照组[39]。也有报道,酸化处理后的三棘刺鱼对水槽中新放入物体的好奇性或者勇敢探索意识降低,但该行为改变的生态意义不确定[34]。
另外,有报道推测,鱼类的生物电场与电(磁)感受能力也可能会受水中CO2分压升高的影响,可能会影响鱼类利用电场的正常生理及行为,如洄游、摄食和繁殖等行为[40-41],但证实该推测的相关研究及结果,还未见报道。
5 海洋酸化对鱼类感觉和行为影响的可能机制(The possible mechanism of ocean acidification on the sensory system and behaviour in marine fish)
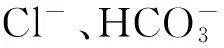
鳃是鱼类重要的气体交换器官,其上有大量的O2化学感受器[45],后来发现鳃部神经表皮细胞不仅能够感知氧分压变化,同时能够感知低浓度的CO2[46-47]。这些神经表皮细胞受到广泛的神经支配,富含多种神经递质,特异性的对CO2敏感而非对H+敏感[48]。当水中CO2分压升高时,会引起神经表皮细胞去极化,进而导致胞内Ca2+浓度快速升高,使突触前膜释放大量的神经递质[49]。但这些神经递质作用的神经网络,如何引起鱼类感觉和行为的变化目前还不清楚。
利用实时定量PCR方法研究海水酸化对青鳉鱼(Oryzias latipes)发育影响,酸化通过下调糖酵解、三羧酸循环和呼吸链等主要代谢通路的基因从而延缓青鳉鱼的胚胎发育,并发现酸化使得青鳉鱼仔鱼、成鱼体内的酸碱代谢基因表达量升高[50]。目前开展的有关海洋酸化对鱼类影响的分子机理的研究很少,但是对其他钙化海洋生物的相关研究,可以为深入研究酸化对鱼类感觉和行为影响的分子机理提供思路。例如,利用高通量测序技术,在酸化处理珊瑚虫(Acropora millepora)9 d后,珊瑚虫最明显的分子响应是抗细胞凋亡的Bcl-2族的5个基因表达量上调,并且热休克蛋白相关基因表达量也有所增加,因此推测珊瑚虫之所以能够快速适应酸化环境与这些基因表达量的上调有关[51]。
目前对于海洋酸化对鱼类感觉和行为影响机制还不了解,需要利用神经行为、电生理、分子生物学等方法,从分子到系统对海洋酸化对鱼类感觉和行为影响的机制进行深入的研究。
6 研究展望(Research prospects)
(1)海洋酸化与其他环境因子协同作用对鱼类感觉和行为的影响
全球气候变化效应的研究中,温度与CO2是紧密相联的。自工业革命以来,空气中人为排放CO2量的增加,导致地球表面平均温度升高了大约0.7 ℃,以此CO2排放速率计算,到21世纪末地球表面温度将升高2~4 ℃,海水pH值下降0.3~0.4[52]。伴随全球变暖,在近岸和大洋生态系统中低氧区分布也在增加,目前大家普遍认为海洋中低氧区海水的CO2分压较高,海水酸化[53]。在海洋生态系统中,温度升高、缺氧与酸化等环境胁迫因子可能同时存在,它们之间共同作用对海洋生物的影响可能比单一海洋酸化的影响更为严重[53-54]。并且,海洋酸化改变海水pH,影响海水碳酸盐平衡,也可能改变海洋中污染物的理化性质和生物毒性[55]。这些因子之间的相互作用对鱼类生理,包括感觉和行为的影响,都需深入的研究。
(2)海洋酸化沿食物链的上行和下行生态效应
生物之间的捕食作用是群落结构形成的一个重要机制,任何影响捕食者-猎物相互关系的因素都会影响到作用双方[16]。目前研究发现,海洋酸化能够通过影响鱼类的感觉和行为,显著增加在自然栖息地的被捕食率[19,56],虽然大家都在强调这些潜在威胁会对海洋种群动力和生态系统过程造成影响,但这其中捕食者-猎物相互作用的动态机制未知。而且,已有研究发现海洋酸化还能够影响海洋中初级生产力和消费者关系,比如,酸化改变海洋藻类的蛋白浓度和次级代谢产物,从而改变不同素食性生物对其喜好,进而改变海洋中藻类丰度和珊瑚礁群落结构[57]。也有报道,在同样酸化程度下,重要的经济种奥林匹亚牡蛎(Ostrea lurida)面临对环境耐受力强的入侵种大西洋钻蚝螺(Urosalpinx cinerea)的更强的摄食压力[58]。因此,在进行海洋酸化对鱼类感觉和行为影响的研究中,也应沿着从初级生产力到高级消费者的食物链,研究海洋酸化对研究捕食者、猎物及两者之间相互作用关系,才能更好的预知酸化生态、经济和社会效应。
(3)鱼类对海洋酸化的适应性进化
要了解海洋酸化对海洋生物多样性的影响,不仅要研究酸化如何影响生物种群,同时也应研究生物对环境变化的适应(acclimation)以及适应性进化(adaptive evolution)情况。目前绝大多数研究是通过短期、单世代实验,模拟未来100~200年海洋酸化对海洋生物的一些表型特征影响,而海洋生物对海洋酸化的长期的适应及适应性进化的研究则非常缺乏。数量遗传学和基因组学方法可用于评估具有生态、社会或系统进化意义的生物对环境变化的适应潜能[59]。无性繁殖、世代时间短的苔藓虫(Celleporella hyalina)[60]、颗石藻(Emiliania huxleyi)[61]实验中多世代的酸化适应性研究已有报道,但对于如海洋鱼类等较大型、世代时间长对酸化适应潜能的研究则亟需开展。
近10年,海洋酸化引起了科学家广泛的关注,相关的基础科学资料快速增加,但关于制定海洋酸化方面的法律法规的分析和评论却很少。在过去的40年间国际社会围绕大气CO2排放争论不止,海洋酸化是继温室效应之后,又一个全球性的“CO2问题”。目前全球就控制空气CO2的排放量尚难达成一致意见,但是可在小范围如地方政府,通过为工厂排污制定更严格的排水标准、水质检测标准中增加海水pH和其他相关参数的标准、沿海地区采用经济能源减少氮硫化合物排放等政策和措施,减缓CO2排放引起的海水酸化。这些自下而上的措施可能并不能解决全球CO2问题,但是可以作为一个较直接的手段应对快速变化的海洋的挑战,较好的保护海洋鱼类的种群繁衍和海洋渔业资源的可持续发展。
[1]Meehl G A, Stocker T F, Collins W D, et al. Global Climate Projections [M]//Solomon S, Qin D, Manning M, et al. eds. Climate Change 2007: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge, UK: Cambridge University Press, 2007: 747-846
[2]Raupach M R, Marland G, Ciais P, et al. Global and regional drivers of accelerating CO2emissions [J]. Proceedings of the National Academy of Sciences, 2007, 104(24): 10288-10293
[3]Hoegh-Guldberg O, Mumby P J, Hooten A J, et al. Coral reefs under rapid climate change and ocean acidification [J]. Science, 2007, 318(5857): 1737-1742
[4]Kleypas J A, Feely R A, Fabry V J, et al. Impacts of Ocean acidification on coral reefs and other marine calcifiers: A guide for future research [R]. Seattle: NOAA/Pacific Marine Environmental Laboratory, 2006
[5]Orr J C, Fabry V J, Aunont O, et al. Anthropogenic ocean acidification over the twenty-first century and its impact on calcifying organisms [J]. Nature, 2005, 437(7059): 681-686
[6]Wässle H. Parallel processing in the mammalian retina [J]. Nature Review Neuroscience, 2004, 5(10): 747-757
[7]Gillespie P, Müller U. Mechanotransduction by hair cells: Models, molecules, and mechanisms [J]. Cell, 2009, 139(1): 33-44
[8]Branch T A, DeJoseph B M, Ray L J, et al. Impacts of ocean acidification on marine seafood [J]. Trends in Ecology and Evolution, 2013, 28(3): 178-186
[9]唐启升, 陈镇东, 余克服, 等. 海洋酸化及其与海洋生物及生态系统的关系[J]. 科学通报, 2013, 58(14): 1307-1314
Tang Q S, Chen Z D, Yu K F, et al. The effects of ocean acidification on marine organisms and ecosystem [J]. Chinese Science Bulletin (Chinese Version), 2013, 58(14): 1307-1314 (in Chinese)
[10]Derby C D, Sorensen P W. Neural processing, perception, and behavioral responses to natural chemical stimuli by fish and crustaceans [J]. Journal of Chemical Ecology, 2008, 34(7): 898-914
[11]Leduc A O H C, Munday P L, Brown G E, et al. Effects of acidification on olfactory-mediated behavior in freshwater and marine ecosystems: A synthesis [J]. Philosophical Transactions of the Royal Society B, 2013, 368(1627): 20120447
[12]Munday P L, Dixson D L, Donelson J M, et al. Ocean acidification impairs olfactory discrimination and homing ability of a marine fish [J]. Proceedings of the National Academy of Sciences, 2009, 106(6): 1848-1852
[13]Devine B M, Munday P L, Jones G P. Rising CO2concentrations affect settlement behavior of larval damselfishes [J]. Coral Reefs, 2012, 31(1): 229-238
[14]Devine B M, Munday P L. Habitat preferences of coral-associated fishes are altered by short-term exposure to elevated CO2[J]. Marine Biology, 2013, 160(8): 1955-1962
[15]Devine B M, Munday P L, Jones G P. Homing ability of adult cardinalfish is affected by elevated carbon dioxide [J]. Oecologia, 2012, 168(1): 269-276
[16]Lima S L, Dill L M. Behavioral decisions made under the risk of predation: A review and prospectus [J]. Canadian Journal of Zoology, 1990, 68(4): 619-640
[17]Wisenden B D. Olfactory assessment of predation risk in the aquatic environment [J]. Philosophical Transactions of the Royal Society of London, Series B, 2000, 355:1205-1208
[18]Kelley J L, Magurran A E. Learned predator recognition and antipredator responses in fishes [J]. Fish and Fisheries, 2003, 4(3): 216-226
[19]Dixson D L, Munday P L, Jones G P. Ocean acidification disrupts the innate ability of fish to detect predator olfactory cues [J]. Ecology Letters, 2010, 13(1): 68-75
[20]Munday P L, Pratchett M S, Dixson D L, et al. Elevated CO2affects the behavior of an ecologically and economically important coral reef fish [J]. Marine Biology, 2013, 160(8): 2137-2144
[21]Checkley D M Jr, Dickson A G, Takahashi M, et al. Elevated CO2enhances otolith growth in young fish [J]. Science, 2009, 324(5935): 1683
[22]Simpson S D, Munday P L, Wittenrich M L, et al. Ocean acidification erodes crucial auditory behaviour in a marine fish [J]. Biology Letters, 2011, 7(6): 917-920
[23]Hurst T P, Fernandez E R, Mathis J T, et al. Resiliency of juvenile walleye pollock to projected levels of ocean acidification [J]. Aquatic Biology, 2012, 17(3): 247-259
[24]Bignami S, Sponaugle S, Cowen R K. Response to ocean acidification in larvae of a large tropical marine fish, Rachycentron canadum [J]. Global Change Biology, 2013, 19(4): 996-1006
[25]Bignami S, Enochs I C, Manzello D P, et al. Ocean acidification alters the otoliths of a pantropical fish species with implications for sensory function [J]. Proceedings of the National Academy of Sciences of the United States of America, 2013, 110(18): 7366-7370
[26]Chung W S, Marshall N J, Watson S A, et al. Ocean acidification slows retinal function in a damselfish through interference with GABAA receptors [J]. The Journal of Experimental Biology, 2014, 217(3): 323-326
[27]Ferrari M C O, McCormick M I, Munday P L, et al. Effects of ocean acidification on visual risk assessment in coral reef fishes [J]. Functional Ecology, 2012, 26(3): 553-558
[28]Ferrari M C O, Wisenden B D, Chivers D P. Chemical ecology of predator-prey interactions in aquatic ecosystems: A review and prospectus [J]. Canadian Journal of Zoology, 2010, 88(7): 698-724
[29]Lönnstedt O M, Munday P L, McCormick M I, et al. Ocean acidification and responses to predators: Can sensory redundancy reduce the apparent impacts of elevated CO2on fish? [J]. Ecology and Evolution, 2013, 3(10): 3565-3575
[30]Dadda M, Koolhaas W H, Domenici P. Behavioural asymmetry affects escape performance in a teleost fish [J]. Biology Letters, 2010, 6(3): 414-417
[31]Sovrano V A, Dadda M, Bisazza A. Lateralized fish perform better than nonlateralized fish in spatial reorientation tasks [J]. Behavioural Brain Research, 2005, 163(1): 122-127
[32]Bisazza A, Dadda M. Enhanced schooling performance in lateralized fishes [J]. Proceeding of Royal Society B, 2005, 272(1573): 1677-1681
[33]Domenici P, Allan B, McCormick M I, et al. Elevated carbon dioxide affects behavioural lateralization in a coral reef fish [J]. Biology Letters, 2012, 8(1): 78-81
[34]Jutfelt F, Bresolin de Souza K, Vuylsteke A, et al. Behavioural disturbances in a temperate fish exposed to sustained high-CO2levels [J]. PloS ONE, 2013, 8(6): e65825. doi:10.1371/journal.pone.0065825
[35]Galef B G, Giraldeau L A. Social influences on foraging in vertebrates: Causal mechanisms and adaptive functions [J]. Animal Behaviour, 2001, 61(1): 3-15
[36]Griffin A S. Social learning about predators: A review and prospectus [J]. Learning & Behavior, 2004, 32(1): 131-140
[37]Ferrari M C O, Manassa R P, Dixson D L, et al. Effects of ocean acidification on learning in coral reef fishes [J]. PloS ONE, 2012, 7(2): e31478. doi:10.1371/journal.pone.0031478
[38]Chivers D P, Mccormick M I, Nilsson G E, et al. Impaired learning of predators and lower prey survival under elevated CO2: A consequence of neurotransmitter interference [J]. Global Change Biology, 2014, 20(2): 515-522 doi: 10. 1111/gcb.12291
[39]Hamilton T J, Holcombe A, Tresguerres M. CO2-induced ocean acidification increases anxiety in rockfish via alteration of GABAAreceptor functioning [J]. Proceedings of the Royal Society, 2014, 281(1775): 20132509
[40]张旭光, 宋佳坤, 张国胜, 等. 食蚊鱼的生物电场特征[J]. 水生生物学报, 2011, 35(5): 823-828
Zhang X G, Song J K, Zhang G F, et al. The bioelectric field to Gambusia affinis [J]. Acta Hydrobiologica Sinica, 2011, 35(5): 823-828 (in Chinese)
[41]Zhang X G, Herzog H, Song J K, et al. Response properties of the electrosensory neurons in hindbrain of the white sturgeon, Acipenser transmontanus [J]. Neuroscience Bulletin, 2011, 27(6): 422-429
[42]Nilsson G E, Dixson D L, Domenici P, et al. Near-future carbon dioxide levels alter fish behavior by interfering with neurotransmitter function [J]. Nature Climate Change, 2012, 2(3): 201-204
[43]Ishimatsu A, Hayashi M, Kikkawa T. Fishes in high-CO2, acidified oceans [J]. Marine Ecology Progress Series, 2008, 373(1): 295-302
[44]Brauner C J, Baker D W. Patterns of acid-base regulation during exposure to hypercarbia in fishes [M]// Cardio-Respiratory Control in Vertebrates. Berlin: Springer, 2009: 43-63
[45]Smith F M, Jones D R. Localization of receptors causing hypoxic bradycardia in trout (Salmo gairdneri) [J]. Canadian Journal of Zoology, 1978, 56(6): 1260-1265
[46]Gilmour K M, Perry S F. Branchial chemoreceptor regulation of cardiorespiratory function [M]//Hara T J, Zielinski B. ed. Sensory Systems Neuroscience. San Diego: Academic Press, 2007: 97-151
[47]Perry S F, Reid S G. Cardiorespiratory adjustments during hypercarbia in rainbow trout (Oncorhynchus mykiss) are initiated by external CO2receptors on the first gill arch [J]. The Journal of Experimental Biology, 2002, 205(21): 3357-3365
[48]Qin Z, Lewis J E, Perry S F. Zebrafish (Danio rerio) gill neuroepithelial cells are sensitive chemoreceptors for environmental CO2[J]. The Journal of Physiology, 2010, 588(5): 861-872
[49]Perry S F, Abdallah S. Mechanisms and consequences of carbon dioxide sensing in fish[J]. Respiratory Physiology and Neurobiology, 2012, 184(3): 309-315
[50]Tseng Y, Hu M Y, Stumpp M, et al. CO2-driven seawater acidification differentially affects development and molecular plasticity along life history of fish (Oryzias latipes) [J]. Comparative Biochemistry and Physiology, Part A, 2013, 165(2): 119-130
[51]Moya A, Huisman L, Foret S, et al. Rapid acclimation of juvenile corals to CO2-mediated acidification by upregulation of heat shock protein and Bcl-2 genes [J]. Molecular Ecology, 2015, 24(2): 438-452
[52]IPCC. Summary for Policymakers [M]// Solomon S, Qin D, Manning M, et al. ed. Climate Change 2007: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge: Cambridge University Press, 2007
[53]Gobler C J, DePasquale E L, Griffith A W, et al. Hypoxia and acidification have additive and synergistic negative effects on the growth, survival, and metamorphosis of early life stage bivalves [J]. PLoS ONE, 2014, 9(1): e83648
[54]Enzor L A, Zippay M L, Place S P. High latitude fish in a high CO2world: Synergistic effects of elevated temperature and carbon dioxide on the metabolic rates of Antarctic notothenioids [J]. Comparative Biochemistry and Physiology, Part A, 2013, 164(1): 154-161
[55]Nikinmaa M. Climate change and ocean acidification-interactions with aquatic toxicology [J]. Aquatic Toxicology, 2013, 126: 365-372
[56]Munday P L, Dixson D L, McCormick M I, et al. Replenishment of fish populations is threatened by ocean acidification [J]. Proceedings of the National Academy of Sciences, 2010, 107(29): 12930-12934
[57]Borell E M, Steinke M, Fine M. Direct and indirect effects of high pCO2on algal grazing by coral reef herbivores from the Gulf of Aqaba (Red Sea) [J]. Coral Reefs, 2013, 32(4): 937-947
[58]Sanford E, Gaylord B, Hettinger A, et al. Ocean acidification increases the vulnerability of native oysters to predation by invasive snails [J]. Proceedings of the Royal Society B, 2014, 281(1778): 2013-2681
[59]Sunday J M, Calosi P, Dupont S, et al. Evolution in an acidifying ocean [J]. Trends in Ecology and Evolution, 2014, 29(2): 117-125
[60]Pistevos J C A, Calosi P, Widdicombe S, et al. Will variation among genetic individuals influence species responses to global climate change? [J]. Oikos, 2011, 120(5): 675-689
[61]Langer G, Nehrke G, Probert I, et al. Strain-specific responses of Emiliania huxleyi to changing seawater carbonate chemistry [J]. Biogeosciences, 2009, 6(11): 2637-2646
◆
Impacts of Ocean Acidification on Sensory System and Behavior in Marine Fish
Wang Xiaojie1,2,3,*, Song Jiakun1,2,3, Fan Chunxin1,2,3, Zhang Xuguang1,2,3, Guo Hongyi1,2,3
1. Institute for Marine Biosystem and Neurosciences, Shanghai Ocean University, Shanghai 201306, China 2. International Center of Marine Study, Shanghai Ocean University, Shanghai 201306, China 3. Key Laboratory of Exploration and Utilization of Aquatic Genetic Resources, Shanghai Ocean University, Ministry of Education, Shanghai 201306, China
6 February 2015accepted 5 May 2015
Ocean acidification is the lowering of ocean pH due to increasing levels of CO2in the atmosphere. This process has resulted in a drop of oceanic pH by 0.1 pH units since pre-industrial times and a further decrease of 0.3-0.4 pH units is predicted by 2100 if maintain current CO2emissions trajectories. Ocean acidification affects its carbonate chemistry, which will threaten many marine organisms and even the whole ecosystem that depend on the stability of chemical environment. Previous studies have focused mainly on calcifying organisms, and marine fish drew less attention because of being have better acid-base regulation system. However, over the past 5 years, this assumption has been challenged by quite a few experiment results on fish larval sensory system and behavioral studies. These studies documented that ocean acidification affected the early life of fish, and the slight changes in early life stage could increase mortality rates of fish in large. That probably will affect recruitment rates to natural population and species diversity, further impact on global fishery catches. This review focused on the impact of ocean acidification on sensory system and behavior in marine fish, including olfactory, hearing, visual systems with their related behavior, and the advanced consciousness studies. It provides perspectives from sensory biology of fish to prediction of ecological, economic and social effect of ocean acidification.
ocean acidification; marine fish; sensory system; behavior; review
国家自然科学基金项目(41306097);中国科学院海洋生态与环境科学重点实验室开放课题(KLMEES2013);上海高校水产学一流学科建设资助项目;上海海洋大学国际海洋研究中心项目
王晓杰(1979-),女,讲师,研究方向为海洋生态和鱼类神经生物学,E-mail: xj-wang@shou.edu.cn
10.7524/AJE.1673-5897.20150206001
2015-02-06 录用日期:2015-05-05
1673-5897(2015)6-013-08
X171.5
A
简介:王晓杰(1979—),女,海洋生物学博士,讲师,主要研究方向为海洋生态学、鱼类神经生物学。
王晓杰, 宋佳坤, 范纯新, 等. 海洋酸化对鱼类感觉和行为影响的研究进展[J]. 生态毒理学报,2015, 10(6): 13-20
Wang X J, Song J K, Fan C X, et al. Impacts of ocean acidification on sensory system and behavior in marine fish [J]. Asian Journal of Ecotoxicology, 2015, 10(6): 13-20 (in Chinese)
