片口自然保护区不同海拔团竹克隆生长研究
2015-08-20刘美张涛谭永燕刘兰娇马李红
刘美 张涛 谭永燕 刘兰娇 马李红
摘要:通过对四川省北川县片口自然保护区不同海拔梯度下团竹的分株形态特征进行调查,研究其克隆生长特征及克隆繁殖的大小依赖关系。结果表明:母株高度、当年生立竹生物量及高度、间隔子直径、比间隔子长、间隔子分枝强度随海拔的改变均有显著性变化;母株、间隔子的生物量受海拔影响不明显。在各海拔环境中,母株大小均显著影响间隔子大小,并通过间隔子的作用显著影响当年生立竹的大小。团竹的克隆生长不仅受海拔梯度的影响,也受自身母株大小的影响,因此可通过改善母株质量而影响克隆分株。
关键词:片口自然保护区;团竹;不同海拔;克隆生长
中图分类号:S795.201 文献标志码: A 文章编号:1002-1302(2015)07-0197-03
团竹(Fargesia obliqua)为禾本科箭竹属植物,是我国濒危物种大熊猫(Ailuropoda melanoleuca)的主食竹之一。该品种近似缺苞箭竹(F.denudata),秆丛生,叶片宽大,基部常不对称,次脉常为4对,易于区分。团竹分布于北川、平武、松潘、茂县4县交界的高山地区,海拔2 500~3 300 m,并为该区垂直分布最高的竹种,其下依次为缺苞箭竹(F.denudata)、青川箭竹(F.rufa)、油竹子(F.angustissima)[1]。克隆生长是指自然条件下,生物通过营养方式自发形成遗传上相同,并具有潜在独立性的新个体或分株的能力或过程[2]。克隆繁殖使得克隆植物具备了较强的水平扩展能力,占据一个大的水平空间,并通过克隆分株的可塑性适应异质性环境[3-9]。植物个体大小与繁殖的关系是植物生活史理论研究的基本内容[6-7,10],有助于了解植物生态适应机制。团竹是多年生一次性开花结实的木质草本克隆植物,自1986年易同培于北川发现以来,对团竹的后续研究报道较少,目前尚无其生态习性、克隆生长构型与形态可塑性、繁殖及适应对策、资源调查等方面的报道。本研究以2004年片口自然保护区团竹开花死亡后新长成的团竹为对象,调查不同海拔环境中团竹的分株形态特征,探讨海拔变化对团竹分株形态特征的影响,以及团竹母株个体大小对克隆分株、间隔子生长的影响,以揭示其克隆繁殖生态适应机制,为保护和培育团竹提供理论依据。
1 试验地区与方法
1.1 试验地区自然概况
北川片口自然保护区地处四川盆地西缘、青藏高原东南缘岷山中段、龙门山西侧,位于北川县片口、小坝、开坪3乡境内,地理坐标为104°10′~104°26′E,32°02′~32°12′N,占地面积8 253 hm2。自然保护区内为北亚热带湿润季风气候,年均气温7.2~11.2 ℃,年均降雨量800 mm。区内土壤呈明显的垂直带谱分布,森林(含灌木林地)覆盖率达997%,植被垂直地带性分布明显。该保护区是以保护大熊猫、金丝猴(Rhinopithecus roxellana)、扭角羚(Budorcas taxicolor)等珍稀野生动物及其自然生态系统为主的自然保护区,是岷山山系自然保护区的典型代表之一[11]。
1.2 调查方法
于2012年10月中旬,在保护区内皮棚子处海拔2 700~2 900 m的团竹种群中开展野外调查。其乔木层主要由岷江冷杉(Abies faxoniana)构成,伴生树种主要有红桦(Betula albo-sinensis)、糙皮桦(Betula utilis)等;灌木层为团竹、花楸(Sorbus pohuashanensis)、紫花杜鹃(Rhododendron amesiae)等;草本层植物稀少。区内团竹种群无明显虫害及大熊猫采食痕迹。
在海拔2 700~2 900 m范围内,每隔100 m分别随机设置3个10 m×10 m的样地。在各样地内随机选择20株(包括一级母株[12]、间隔子、当年生竹笋长成的立竹)完整团竹分株对,分别记录母株和当年生立竹的高度和基径、间隔子的长度和直径、地下茎分蘖数(分枝强度)。用自封袋装好带回实验室,洗尽泥沙后将母株、间隔子、当年生立竹进行分装,于65 ℃恒温烘干至恒质量,冷却后使用万分之一电子天平称质量并记录。
1.3 数据分析
采用SPSS 18.0软件分析所有试验数据;采用One-way ANOVA软件分析不同海拔环境下团竹分株形态特征的差异性,并作LSD多重比较;采用Regression软件分析克隆分株的生物量模型、母株和克隆生长参数的回归关系,并检验显著度。其中,比间隔子长为间隔子长与间隔子生物量的比值[7];间隔子分枝强度为地下茎分蘖数。
2 结果与分析
2.1 不同海拔团竹分株的个体特征
团竹分株个体形态特征中,母株高度、当年生立竹生物量和高度、间隔子直径均随海拔的升高而不断增大,且差异显著。比间隔子长随海拔的升高呈先减小后增大的趋势,且差异极显著。间隔子分枝强度随海拔的升高呈先增大后减小的趋势,且差异极显著。母株生物量和基径、间隔子長度和生物量均无明显差异(表1)。
2.2 不同海拔团竹分株的生物量模型
团竹母株生物量和基径、当年生立竹生物量和基径、间隔子生物量和直径的回归关系见表2。随着海拔由低到高,母株生物量随基径的增大分别呈指数增长、三次曲线增长、线性增长趋势,总体呈二次曲线增长趋势,各增长均达极显著水平;当年生立竹生物量随基径的增大分别呈对数增长、二次曲线增长、线性增长趋势,总体呈指数增长趋势,各增长均达极显著水平;间隔子生物量和直径分别呈指数增长、线性增长、指数增长趋势,总体间隔子生物量随直径的增大呈指数增长趋势,各增长均达极显著水平。
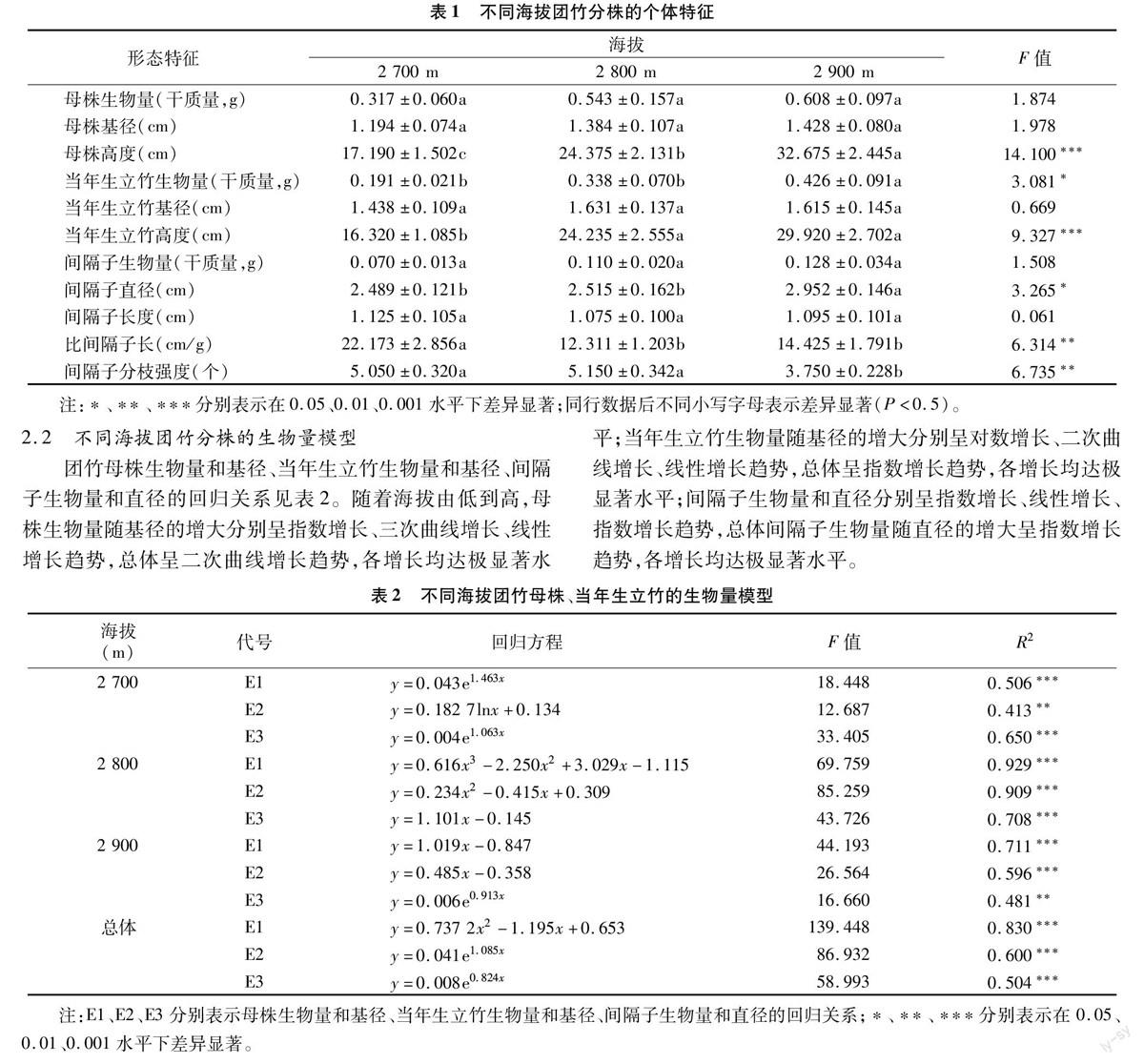
2.3 不同海拔母株与间隔子的回归关系
在海拔2 700、2 900 m处,间隔子直径随母株基径的增大均呈显著的二项式增长;在海拔2 800 m处,间隔子直径随母株基径的增大呈极显著的线性增长。间隔子直径随母株生物量的增大呈极显著的线性增长(图1)。
2.4 不同海拔母株与当年生立竹的回归关系
在海拔2 700 m处,当年生立竹基径与母株基径、母株生物量均呈显著的线性增长;在海拔2 800 m处,当年生立竹基径随母株基径的增长呈极显著的线性增长,随母株生物量的增长呈极显著的对数增长;在海拔2 900 m处,当年生立竹基径与母株基径呈极显著的指数增长,与母株生物量呈极显著的线性增长(图2)。
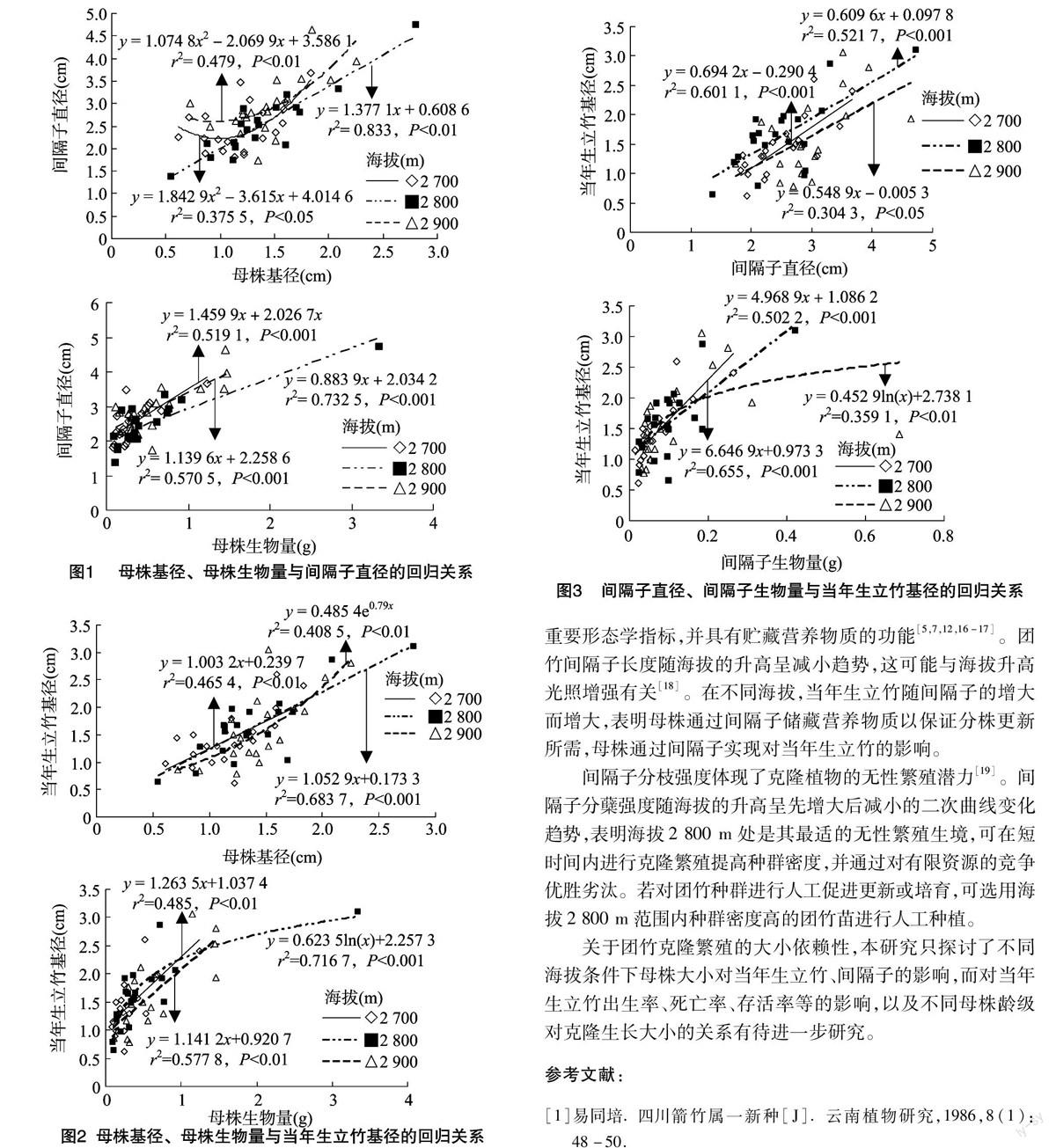
2.5 间隔子对当年生立竹的影响
在海拔2 700、2 800 m处,当年生立竹基径随间隔子直径、间隔子生物量的增大均呈极显著的线性增长;在海拔2 900 m处,当年生立竹基径与间隔子直径呈极显著的线性增长,与间隔子生物量呈极显著的对数增长(图3)。
3 结论与讨论
海拔梯度包含了温度、水分、光照等环境因子的剧烈变化,因此成为研究植物环境适应性、对全球气候变化响应的理想区域[13]。本研究中各海拔条件下,团竹母株的生物量、基径、高度,当年生立竹的生物量、高度,间隔子的生物量、直径均随海拔的升高而不断增大。潘红丽等在对卧龙自然保护区华西箭竹的研究中发现,生物量随海拔的升高呈先升高后降低的趋势[13];刘兴良等在研究中发现,生物量随海拔的升高呈下降趋势[14]。以上结论均与本研究结果不一致,这可能是由于各品种的最适生长海拔不同,是物种适应异质性环境的结果。
物种的繁殖需要消耗大量营养物质,植物个体大小是衡量植物对资源积累的重要指标之一[6-7,12]。以基径和生物量衡量团竹母株大小,发现在不同海拔条件下,当年生立竹、间隔子大小均随母株的增大而增大,呈显著正相关,可见团竹的克隆生长在个体水平具有大小依赖关系,这与解蕊等、王琼等、刘左军等的研究结果[7,12,15]一致。
间隔子是研究克隆植物在资源异质性环境中觅食行为的
重要形态学指标,并具有贮藏营养物质的功能[5,7,12,16-17]。团竹间隔子长度随海拔的升高呈减小趋势,这可能与海拔升高光照增强有关[18]。在不同海拔,当年生立竹随间隔子的增大而增大,表明母株通过间隔子储藏营养物质以保证分株更新所需,母株通过间隔子实现对当年生立竹的影响。
间隔子分枝强度体现了克隆植物的无性繁殖潜力[19]。间隔子分蘖强度随海拔的升高呈先增大后减小的二次曲线变化趋势,表明海拔2 800 m处是其最适的无性繁殖生境,可在短时间内进行克隆繁殖提高种群密度,并通过对有限资源的竞争优胜劣汰。若对团竹种群进行人工促进更新或培育,可选用海拔2 800 m范围内种群密度高的团竹苗进行人工种植。
关于团竹克隆繁殖的大小依赖性,本研究只探讨了不同海拔条件下母株大小对当年生立竹、间隔子的影响,而对当年生立竹出生率、死亡率、存活率等的影响,以及不同母株龄级对克隆生长大小的关系有待进一步研究。
参考文献:
[1]易同培. 四川箭竹属一新种[J]. 云南植物研究,1986,8(1):48-50.
[2]董 鸣,于飞海. 克隆植物生态学术语和概念[J]. 植物生态学报,2007,31(4):689-694.
[3]Cook R E. Growth and development in clonal plant populations[M]. New haven:yale university press,1985:259-296.
[4]董 鸣. 异质性生境中的植物克隆生长:风险分摊[J]. 植物生态学报,1996,20(6):543-548.
[5]宋利霞,陶建平,冉春燕,等. 卧龙亚高山暗针叶林不同林冠环境下华西箭竹的克隆生长[J]. 植物生态学报,2007,31(4):637-644.
[6]解 蕊. 亚高山不同针叶林冠下大熊猫主食竹的克隆生长[D]. 北京:北京林业大学,2009:3-34.
[7]解 蕊,李俊清. 亚高山针叶林冠下缺苞箭竹的克隆生长[J]. 东北林业大学学报,2009,37(8):22-25.
[8]陶建平,宋利霞. 亚高山暗针叶林不同林冠环境下华西箭竹的克隆可塑性[J]. 生态学报,2006,26(12):4019-4026.
[9]Doust L L. Population dynamics and local specialization in a clonal perennial(Ranunculus repens).Ⅲ.Responses to light and nutrient supply[J]. Journal of Ecology,1987,75(2):555-568.
[10]Klinkhamer P L,Meelis E,Jong T J,et al. On the analysis of size-dependent reproductive output in plants[J]. Functional Ecology,1992,6(3):308-316.
[11]岳 鹏,周 勇. 四川片口自然保护区总体规划[R]. 成都:四川省林业厅,四川省林业勘察设计研究院,2005:1-75.
[12]王 琼,苏智先,雷泞菲,等. 慈竹母株大小对克隆生长的影响[J]. 植物生态学报,2005,29(1):116-121.
[13]潘红丽,田 雨,刘兴良,等. 卧龙自然保护区华西箭竹(Fargesia nitida)生态学特征随海拔梯度的变化[J]. 生态环境学报,2010,19(12):2832-2839.
[14]刘兴良,刘世荣,宿以明,等. 巴郎山川滇高山栎灌丛地上生物量及其对海拔梯度的响应[J]. 林业科学,2006,42(2):1-7.
[15]刘左军,杜国祯,陈家宽. 不同生境下黄帚橐吾(Ligularia virgaurea)个体大小依赖的繁殖分配[J]. 植物生態学报,2002,26(1):44-50.
[16]Kleiju D,Van G J M. The exploitation of heterogeneity by a clonal plant in habitats with contrasting productivity levels[J]. Journal of Ecology,1999,87:873-884.
[17]Dong M,De K H. Plasticity in morphology and biomass allocation in cynodon dactylon,a grass species forming stolons and rhizomes[J]. Oikos,1994,70:99-106.
[18]罗学刚,董 鸣. 蛇莓克隆构型对光照强度的可塑性反应[J]. 植物生态学报,2001,25(4):494-497.
[19]吴福忠,王开运,杨万勤,等. 大熊猫主食竹群落系统生态学过程研究进展[J]. 世界科技研究与发展,2005,27(3):79-84.
