产青蒿二烯的人工酵母细胞的构建及发酵优化
2015-06-19郭睿丁明珠元英进
郭睿,丁明珠,元英进
(系统生物工程教育部重点实验室,天津大学化工学院制药工程系,天津化学化工协同创新中心,天津300072)
引 言
青蒿素及其衍生物已被世界卫生组织认定为目前世界上最有效的治疗脑型疟疾和抗氯喹恶性疟疾药物。目前主要通过青蒿类植物提取[1]、化学合成[2]以及利用合成生物学手段[3]等方式获得青蒿素。合成生物学由于快速安全绿色环保等优势逐渐替代植物提取和化学全合成[4]。青蒿二烯(amorphadiene)是青蒿素的重要前体,青蒿植物中青蒿二烯的合成分两个步骤:(1)乙酰辅酶A经MVA途径生成法尼基焦磷酸[5](FPP);(2)青蒿二烯合成酶(ADS)[6]催化 FPP 经环化反应生成青蒿二烯。FPP是倍半萜类化合物的共同前体,主要通过两条独立的生物途径合成[7]:甲羟戊酸途径(MVA途径,存在于真核生物、古生菌和部分原核生物中)和 2-C-甲基-D-赤藻糖醇-4-磷酸途径 (MEP途径,存在于细菌和光合真核生物中)。
目前,利用合成生物学方法进行青蒿二烯合成研究中应用最广泛的底盘细胞是大肠杆菌和酿酒酵母[7]。大肠杆菌中存在具有较大萜类合成潜力的MEP途径,适合萜类化合物碳骨架的合成[8]。Keasling课题组在大肠杆菌体内重构MVA途径,优化后青蒿二烯产量达到27.4 g·L−1[9-11]。但大肠杆菌缺少糖基化和翻译后修饰等功能,因此限制了后续青蒿酸合成基因高效表达。而酿酒酵母遗传背景清楚、基因操作简单、可进行大规模发酵生产,且次级代谢产物简单[12],因此较多研究者选择酿酒酵母作为青蒿二烯生物合成的底盘细胞。Kong等[13]通过在酿酒酵母体内过表达tHMGR和ERG20基因,并表达突变的ADS基因,使青蒿二烯产量达到123.2 mg·L−1。Keasling 课题组[14]通过过表达酿酒酵母MVA路径基因和ADS基因,抑制鲨烯合酶表达,使青蒿二烯产量高达40 g·L−1。本课题组之前在该领域的工作主要集中在酿酒酵母底盘细胞与不同功能模块之间适配性研究[15-16]。
HMGR催化MVA途径中第一个可逆反应,是MVA途径上第一个限速酶[17]。表达截短的HMGR基因——tHMGR(去除HMGR的调控区域)可以增加 HMGR的表达量[18],从而提高后续产物的产量[10]。增加ERG20基因的表达量[19]或减少FPP流向其他支路的流量[20],可以增加FPP的积累量,从而增加青蒿二烯的产量。Pitera等[21]研究发现增加HMGR和ERG20基因的拷贝数,可以提高青蒿二烯的产量。Delta位点在基因组中拷贝数大于100,利用 Delta位点进行外源基因整合,在没有选择压力的存在下传代50次整合基因型仍稳定存在,引入的外源基因经过传代后仍能多拷贝存在[22-23]。
本研究通过过表达酿酒酵母tHMGR和ERG20基因、表达外源青蒿二烯合酶基因ADS实现酿酒酵母生产青蒿二烯的目的,如图1所示。之前的大量研究都集中在上调或基因组单位点整合过表达内源基因,以基因组整合或导入外源质粒表达ADS基因,实现酿酒酵母中合成青蒿二烯的目的。内源基因多拷贝过表达,往往要进行多次整合,加大了实验的工作量;若采取多拷贝游离质粒方式表达外源ADS基因,则不能保证人工菌株基因型的稳定性。本研究利用本实验室构建的酵母启动子、终止子载体模块库[24],以酵母基因组中多拷贝位点Delta作为整合位点,不仅可以单次整合多个基因,并且整合后的基因都以多拷贝形式存在于基因组,从而实现酵母内源基因tHMGR和ERG20的多拷贝过表达以及ADS基因的异源多拷贝表达,构建可以生产青蒿二烯且基因型较稳定的人工酿酒酵母菌株。除了考虑葡萄糖浓度以及氧气通量等常用参数外,本研究还将某些必需氨基酸作为提高青蒿二烯产量的关键调控参数。
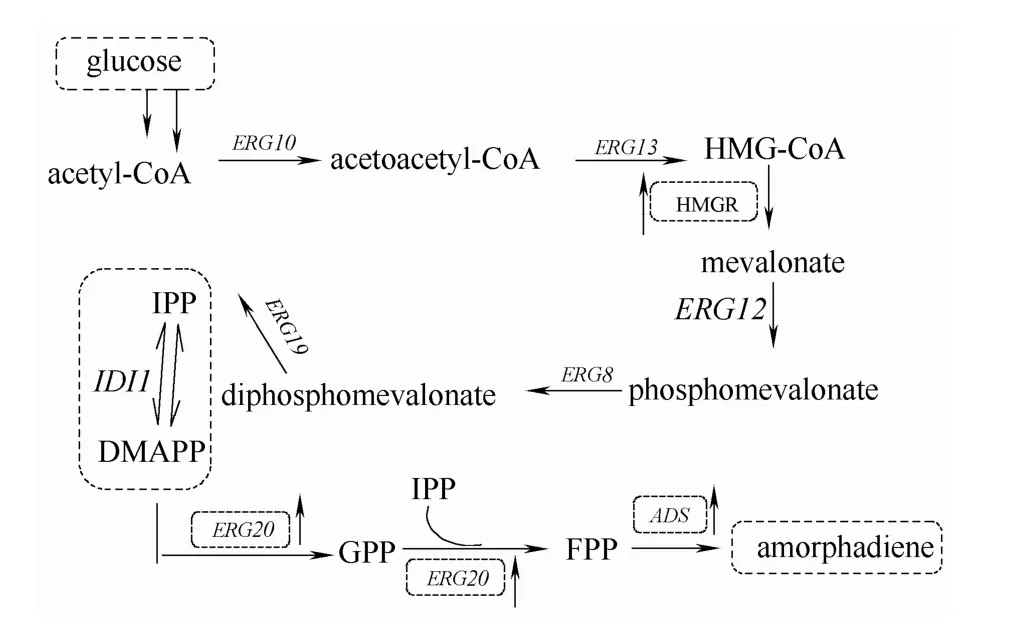
图1 酿酒酵母体内青蒿二烯合成路径的构建Fig.1 Biosynthesis of amorphadiene in engineered S.cerevisiae
1 实验材料和方法
1.1 工具酶及试剂
大肠杆菌感受态细胞DH5α购自Biomed公司,TransStart FastPfu高保真DNA聚合酶购自TransGen公司;T4 DNA连接酶和限制性内切酶 PstI和BamHI购自Fermentas公司;限制性内切酶NotI-HF和BsaI-HF购自NEB公司;质粒小提试剂盒、酵母质粒小提试剂盒、普通 DNA产物纯化试剂盒、琼脂糖凝胶 DNA回收试剂盒购自天根公司;朱栾倍半萜标准品(Valencene)购自TCI公司;正十二烷购自天津市康科德试剂化工厂;PEG3350、鲑鱼精DNA、各类氨基酸粉末购自鼎国昌盛公司;醋酸锂购自北方天医化学试剂厂;去氨基酸酵母氮源(YNB)购自GENVIEW公司。
1.2 菌株、质粒和培养基
宿主菌E. coliDH5α {endA1; hsdR17; gyrA96;thi-1; relA1; supE44; recA1; ΔlacU169(ɸ80lac ΔZM15)} 和 酿 酒 酵 母 菌 株 W303-1a(MATa{leu2-3,112; trp1-1; can1-100; ura3-1; ade2-1;his3-11,15}),酿酒酵母菌株 BY4741(MATa{his3 Δ1;leu2 Δ0; met15 Δ0; ura3 Δ0} ), 酿 酒 酵 母 菌 株CEN.PK2-1C(MATa {ura3-52; trp1-289; leu2-3_112;his3 Δ1; MAL2-8C; SUC2}), 酿 酒 酵 母 菌 株CEN.PK2-1D (MATα{ura3-52; trp1-289; leu2-3_112;his3 Δ1; MAL2-8C; SUC2})购自 Euroscarf公司,酵母表达载体 pRS425K为本实验室构建,详见表1、表2。
LB 培养基(10 g·L−1氯化钠;10 g·L−1胰蛋白胨;5 g·L−1酵母提取物;固体培养基添加1.5%琼脂粉)用于大肠杆菌的培养;YPD培养基(20 g·L−1葡萄糖;20 g·L−1胰蛋白胨;10 g·L−1酵母提取物;固体培养基添加 2%琼脂粉)用于酿酒酵母的培养;SC-ura培养基(20 g·L−1葡萄糖;6.7 g·L−1YNB;2 g·L−1drop-out 氨基酸混合物;固体培养基添加2% 琼脂粉)用于酵母转化子筛选和培养。
1.3 酿酒酵母表达载体构建

表1 实验中涉及的酵母菌株Table 1 Yeast strains involved in this study
以BY4741基因组DNA为模板,通过特异性引物PCR扩增tHMGR和ERG20、终止子片段TTPI1和TENO2,用限制性内切酶BsaI对以上片段和本实验室构建 pRS425K-启动子-终止子模块[24]LD55和LD19进行酶切;纯化回收产物经T4连接酶连接,得到表达载体 pRS425K:TCYC1-PTEF1-tHMGR-TTEF1和 pRS425K: TTEF1-PGPM1-ERG20-TTPI1。用 overlap PCR方法得到TTPI1-TENO2片段,以BamHI和PstI对扩增片段和载体pRS425K分别进行酶切,纯化回收片段以 T4连接酶连接,得到质粒 pRS425K:TTPI1-TENO2。
外源基因ADS参考GeneBank ID.AF-138959报道的序列经密码子优化,利用Genewiz公司合成的60bp Oligos,用overlap PCR方法合成。利用BsaI限制性内切酶对ADS基因和相应的启动子终止子元件LD01D进行酶切,纯化回收产物,经T4连接酶连接得到 pRS425K:TENO2-PPDC1-ADS-TGPM1-Delta2表达载体。

表2 实验中涉及的质粒Table 2 Plasmids involved in this study
1.4 表达模块整合基因组和转化子筛选
以上4个表达载体测序正确后,用 BamHI和PstI酶切,即可得到片段TCYC1-PTEF1-tHMGR-TTEF1、TTEF1-PGPM1-ERG20-TTPI1、TTPI1-TENO2和 TENO2-PPDC1-ADS-TGPM1-Delta2四个片段,用NotI限制性内切酶对 LDL06[24](pRS425K: Delta1-URA3-TCYC1)进行酶切,得到Delta-URA3-CYC1t片段。
采用醋酸锂转化法用上述 5个片段分别以W303-1a、BY4741、CEN.PK2-1C、CEN.PK2-1D为宿主菌株进行酵母转化,用SC-ura培养基筛选,30℃恒温倒置培养2~3 d,至长出重组克隆,分别挑取8个单菌落在SC-ura固体培养基上划线分纯,30℃恒温倒置培养至长出单菌落,挑取单菌落到SC-ura液体培养基,30℃、200 r·min−1培养至OD600约为 4,取菌液 12000 r·min−1离心 2 min,弃上清收集菌体提取基因组,基因组PCR验证过表达模块和异源表达模块,出现正确大小的目标条带,则证明筛选到了正确的阳性克隆,得到人工酵母菌株 SyBE_Sc00011103、SyBE_Sc00011106、SyBE_Sc00011101、SyBE_Sc00011107。
人工酵母菌株功能模块构建流程如图2所示。分别构建tHMGR, ERG20, ADS基因的表达盒质粒,酶切得到 DNA片段,醋酸锂法酵母转化,将各个片段整合到酵母染色体上,经过PCR验证筛选正确克隆。
1.5 人工酵母菌株发酵
1.5.1 摇瓶发酵 将正确阳性克隆接种到 5 ml SC-ura液体培养基中,30℃、200 r·min−1培养至OD600值约为5.0,以初始OD600值0.1转接至SC-ura液体培养基,30℃、200 r·min−1培养12 h,按初始OD600值0.05转接到50 ml液体培养基中,30℃、200 r·min−1培养,10 h后在发酵液中加入2.5 ml正十二烷进行两相培养,以此条件继续培养至72 h。1.5.2 发酵罐发酵 将正确阳性克隆接种到 5 ml SC-ura液体培养基中,30℃、200 r·min−1培养至OD600值约为5.0,以初始OD600值0.1转接至SC-ura液体培养基中,30℃、200 r·min−1培养12 h,初始OD600值为0.05,初始培养基体积2 L,温度30℃,pH为5.8,发酵培养10 h,加入400 ml正十二烷两相培养。当发酵液中葡萄糖浓度接近2 g·L−1时,批式补料方式补给葡萄糖母液,使培养基中葡萄糖浓度到初始值。在OD600值进入平台期后,按初始浓度补加氨基酸和YNB母液1~2次,至菌体密度保持24 h不再增加为止。
1.6 青蒿二烯的提取与检测
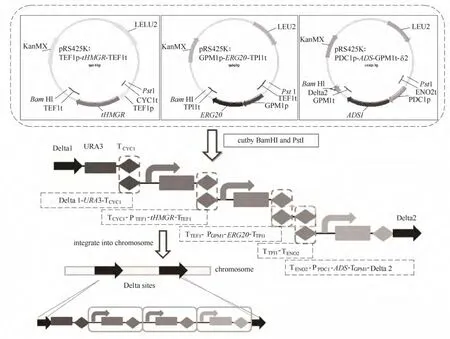
图2 基因整合步骤示意图Fig.2 Schematic of integrating genes into chrosome
发酵结束后,取发酵液 5000 r·min−1离心 10 min,吸取十二烷相,无水硫酸钠干燥,用0.22 μm有机相滤膜过滤,用正己烷稀释,用 GC-TOF/MS检测产物。色谱条件:DB-5MS毛细管色谱柱(30 m×0.25 mm×0.25 μm);升温程序:初始温度100℃,保持 2 min,以8℃·min−1升温至 160℃;以25℃·min−1升温至250℃,保持5 min;进样口温度250℃,分流进样(分流比1:50),进样体积1 μl。质谱条件:EI+源;电子能量70 eV;离子源温度250℃;溶剂延迟时间 5 min;质量扫描方式为选择全离子扫描;定性离子为m/z204(青蒿二烯分子C15H24)和189([M-CH3]+)。
定量方法:使用相同色谱条件测定青蒿二烯的同分异构体朱栾倍半萜的标准溶液(5~150 mg·L−1),绘制浓度-峰面积标准曲线,对青蒿二烯进行定量。
2 实验结果与结论
2.1 酿酒酵母底盘的构建
将 TCYC1-PTEF1-tHMGR-TTEF1、TTEF1-PGPM1-ERG20-TTPI1、TENO2-PPDC1-ADS-TGPM1-Delta2、Delta-URA3-TCYC1以及TTPI1-TENO2等片段同时进行酵母转化,便可以多拷贝位点Delta为整合位点将tHMGR、ERG20、ADS基因依次整合到 CEN.PK2-1C、W303-1a、BY4741和CEN.PK2-1D基因组。经基因组 PCR验证正确的重组转化子,分别命名为:SyBE_Sc00011101、SyBE_Sc00011103、SyBE_Sc00011106、SyBE_Sc00011107。
酿酒酵母MVA途径以乙酰辅酶A为底物,提供倍半萜类化合物合成前体FPP,因此增加FPP的积累量可以增加酿酒酵母生产倍半萜类化合物如青蒿二烯的能力。过表达tHMGR基因和ERG20基因可以增加FPP的积累量。关键酶基因的拷贝数影响青蒿二烯产量[21],但是表达多拷贝质粒的人工菌株往往基因型不够稳定[25],而单拷贝整合过表达目标基因对产物产量的提升不显著。本研究选用多拷贝序列 Delta作为整合位点[22-23],实现了关键酶基因的多拷贝表达,并且以基因组形式稳定存在。
2.2 人工酿酒酵母细胞底盘的优选
进行人工酵母菌株 50 ml摇瓶发酵。用GC-TOF/MS检测产物,在保留时间为10.34 min时检测到青蒿二烯的特征质谱峰m/z=204和m/z=189。根据相同测定条件下得到的朱栾倍半萜浓度-峰面积标准曲线,对人工酵母细胞发酵液中青蒿二烯浓度进行定量。
实验结果表明,表达功能模块的底盘细胞的生长状况以及底盘细胞与功能模块之间适配性的差异,导致人工细胞的生长情况和目标产物积累状况的差异[16]。结果表明,底盘细胞 CEN.PK2由于其较好的生理学特征和生孢能力[25-26],从菌体生长状况和青蒿二烯浓度(表 3)上都显示出明显优势。因此,选择人工酵母菌株SyBE_Sc00011101进行后续研究。
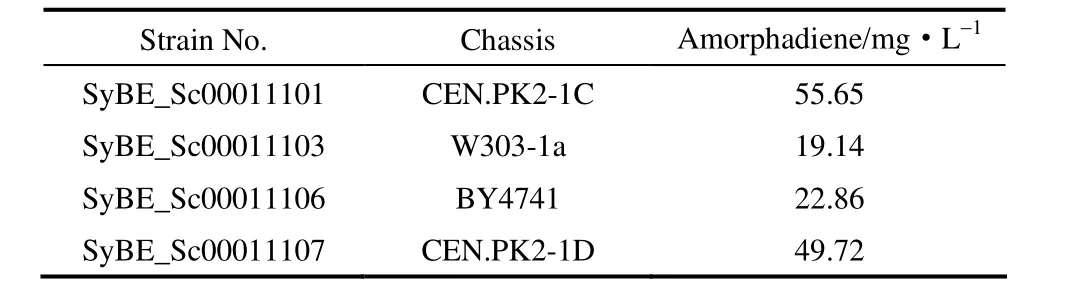
表3 不同底盘的人工菌株青蒿二烯产量Table 3 Production of amorphadiene in artificial strains with different chassis
2.3 发酵优化提高产量
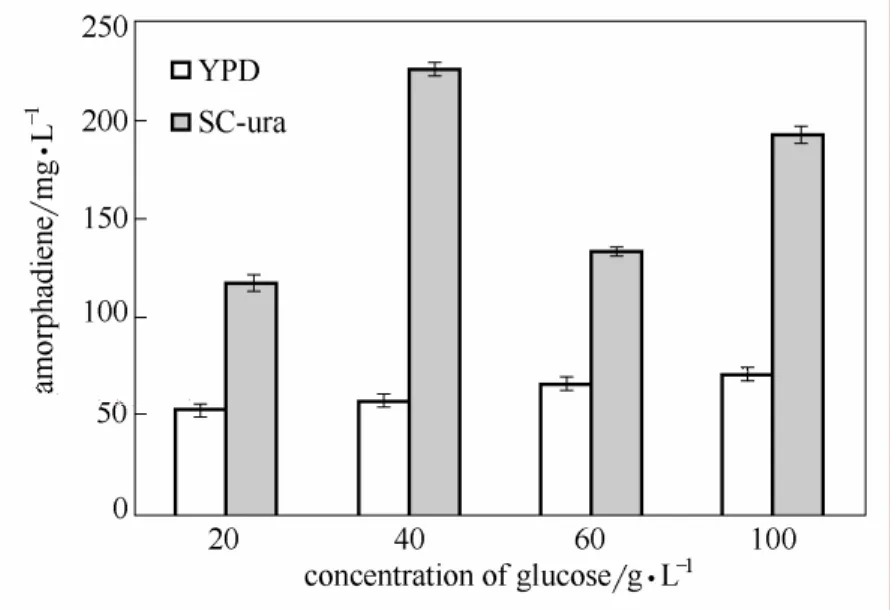
图3 不同葡萄糖浓度下YPD和SC-ura培养基中青蒿二烯产量Fig.3 Production of amorphadiene in YPD and SC-ura with different glucose concentration
选择产量较高的菌株 SyBE_Sc00011101进行摇瓶发酵优化,以期获得更高的产量。
2.3.1 摇瓶发酵培养基成分对发酵过程中青蒿二烯积累的影响 培养条件对人工酿酒酵母的生长状况和青蒿二烯产量影响较大。以不同培养基用SyBE_Sc00011101进行50 ml摇瓶发酵实验。实验发现,菌体在SC-ura培养基中生长速度不及YPD培养基中,但图3青蒿二烯产量的比较显示,菌体在SC-ura培养基中青蒿二烯产量高于YPD培养基中。Albers等[27]发现无氧发酵条件下氮源对酿酒酵母生长速率和产乙醇能力影响较大,因此氮源种类可能是菌体在两种培养基中的生长能力和青蒿二烯生产能力差别的重要因素。培养基中葡萄糖浓度对青蒿二烯产量影响明显。葡萄糖浓度为40 g·L−1的 SC-ura培养基中青蒿二烯浓度最高为 225.3 mg·L−1。
2.3.2 发酵罐分批次补料发酵 选择 SyBE_Sc00011101以葡萄糖浓度为20 g·L−1的SC-ura培养基进行5 L发酵罐实验,初始培养基体积2 L,温度30℃,pH恒定为5.8,在发酵培养10 h时,加入400 ml正十二烷两相培养。
为研究发酵罐中溶氧对发酵过程和产物积累的影响,设定转速和空气流量分别为250 r·min−1、2 L·min−1和 500 r·min−1、4 L·min−1,以 SC-ura(葡萄糖 20 g·L−1)培养基对 SyBE_Sc00011101菌株进行发酵培养,发酵过程中以分批补料方式补给葡萄糖,在不限制生长潜力的前提下,保证菌体有氧呼吸代谢[28]。生长进入平台期后,补加氨基酸和YNB至初始浓度。结果(图4)表明,在转速较快、空气流量较大时,OD600值明显较高;发酵培养至155 h时,转速为250 r·min−1、空气流量为2 L·min−1的发酵罐中青蒿二烯浓度为 335.51 mg·L−1;而转速为 500 r·min−1、空气流量为 4 L·min−1的发酵罐中青蒿二烯浓度为562.81 mg·L−1,是前者的1.68倍。
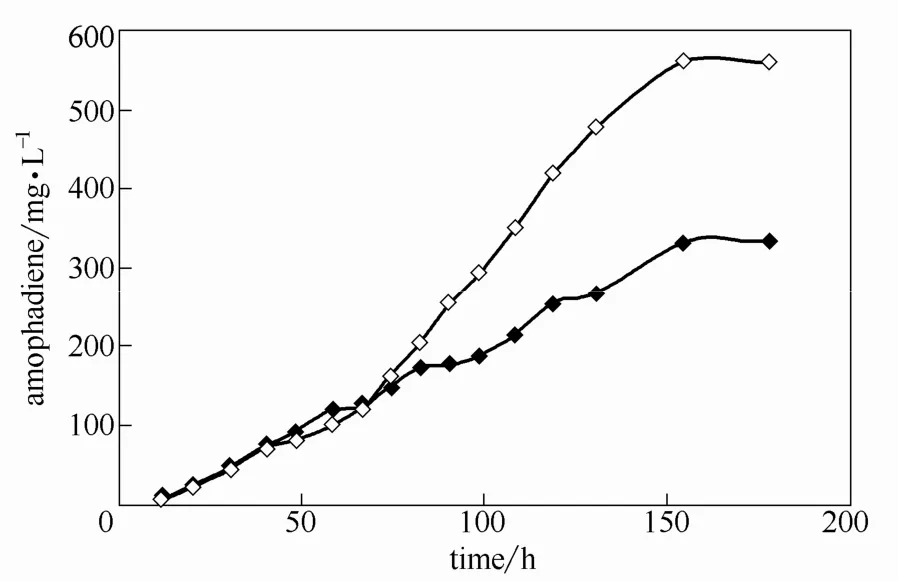
图4 发酵罐转速和空气通量的优化Fig.4 Optimization of rotation speed and air flux
这可能是由于随着溶氧量的增大,人工酵母细胞的呼吸代谢加强,葡萄糖生成丙酮酸的过程加快,同时削弱了无氧条件下丙酮酸生成乙醇和乙酸的过程,而生成更多乙酰辅酶A,经MVA途径为青蒿二烯合成提供更充足的FPP。通过增加底物浓度加快反应速率,从动力学上加快青蒿二烯积累;根据巴斯德效应,酵母在氧气充足时消耗更少的葡萄糖,产生更多的能量,在热力学上促进了青蒿二烯的积累。
由于底盘细胞 CEN.PK2-1C部分氨基酸基因缺陷,经外源基因整合后仍不能合成部分必需氨基酸,如亮氨酸(Leu)、组氨酸(His)和色氨酸(Trp),需要在培养基中添加这些氨基酸。Watson[29]研究发现不同氮源和氨基酸比例影响酵母生长。培养基中这3种氨基酸的浓度是否会影响产量,仍需要进一步探索。选取 Leu、His、Trp初始浓度分别为 0.1 g·L−1、0.02 g·L−1、0.02 g·L−1和 1 g·L−1、0.2 g·L−1、0.2 g·L−1的 SC-ura 培养基(分别记为 SC-ura和 SC-ura+10aa),设定发酵条件为:初始OD600为0.05,培养基体积2 L,pH为5.8,转速 500 r·min−1,空气流量 4 L·min−1,10 h 后加入400 ml正十二烷两相培养。当葡萄糖浓度接近2 g·L−1时,补加葡萄糖母液使其浓度达到约20 g·L−1,菌体进入生长平台期时按初始浓度补加YNB和氨基酸。
实验结果(图5)表明:(1)发酵培养155 h时 SC-ura+10aa培养基中青蒿二烯浓度为 1.05 g·L−1;SC-ura培养基中青蒿二烯浓度为 562.81 mg·L−1,说明该人工酵母菌株生产青蒿二烯的过程受到氨基酸浓度的影响。(2)发酵培养35 h后,SC-ura培养基中人工菌株生长进入平台期,在40 h补加 YNB和氨基酸母液至初始浓度,随后菌体二次生长,说明氮源以及氨基酸浓度会限制该菌株的生长。但若菌株生长速度过快,而空气通量保持不变,则单位OD600菌体溶氧量降低,一定程度上加快了单位菌体消耗葡萄糖的速率,而氮源的不充足使得人工菌株相关代谢失衡,所以SC-ura培养基中虽然有较高的菌体终密度,但青蒿二烯终浓度却远低于SC-ura+10aa培养基中。
本文通过多拷贝位点整合过表达MVA途径关键基因HMGR以及FPP合成酶基因ERG20,增加MVA路径通量以及FPP的积累量;同时表达青蒿二烯合成酶ADS,虽然经过发酵优化,在5 L发酵罐中青蒿二烯的产量达到1.05 g·L−1,但仍与国内外该领域相关研究存在差距。因此除了HMGR和ERG20之外,FPP向萜类物质的通量应该还受到其他因素的抑制,比如代谢流的平衡以及支路分流的作用。因此,在后续的研究中,将会继续对相关代谢路径上物料的均衡分配进行相关研究以达到提高青蒿二烯产量的目的。
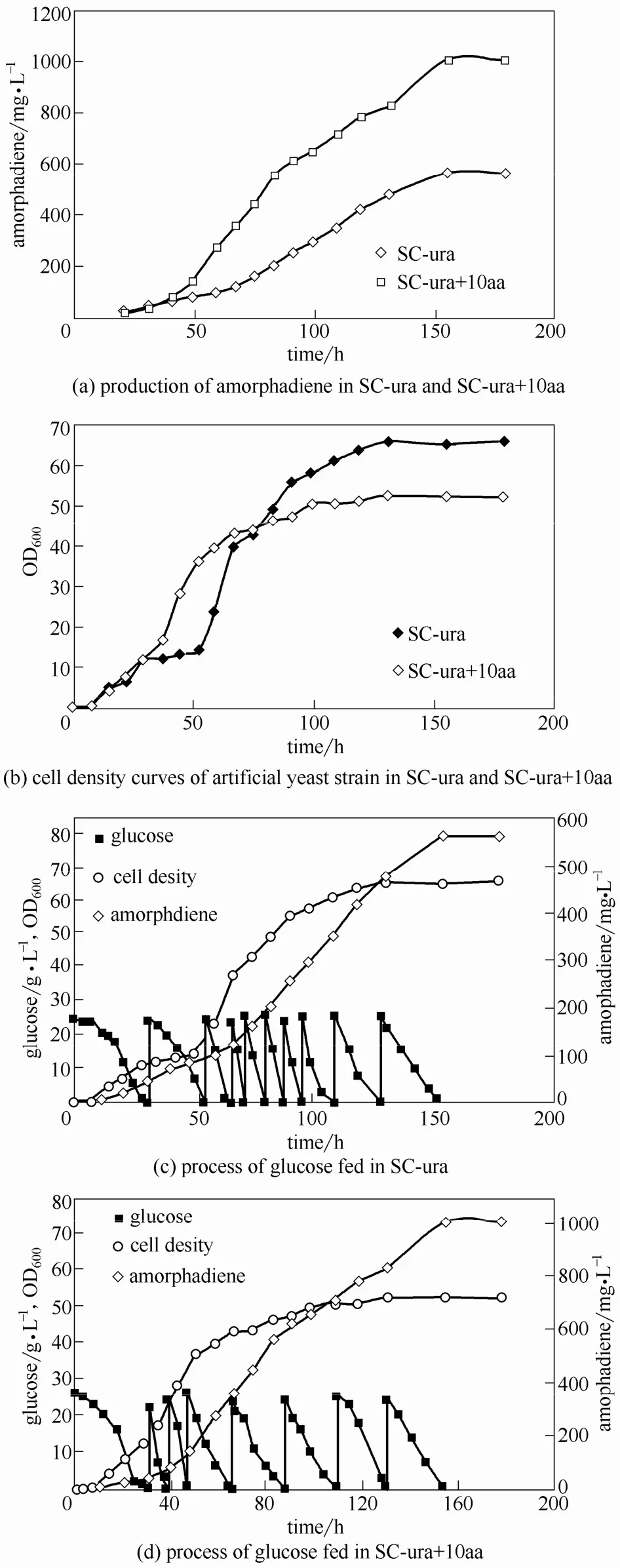
图5 氨基酸浓度和补料发酵过程的优化Fig.5 Optimization of amino acid concentration and fed-batch process
3 结 论
(1)为了增加基因拷贝数,同时克服多拷贝质粒容易丢失的问题,选用基因组上多拷贝位点Delta作为整合位点,可以增加人工酿酒酵母细胞经过MVA途径合成FPP的能力,从而提高青蒿二烯的产量。
(2)发酵过程中,培养基不同会影响菌体浓度和青蒿二烯产量。本研究中SC-ura培养基中青蒿二烯的产量高于 YPD培养基;并且菌体的生长和生产之间存在平衡关系,菌体的生长速率和青蒿二烯的生产速率并不呈正相关。
(3)培养基中葡萄糖浓度影响菌体生长,并且这种影响会反映到青蒿二烯的积累过程上。同时菌体生长依赖于部分必需氨基酸,由于氨基酸是组成蛋白的基本单位,因此对菌体生长以及青蒿二烯的生产影响显著。本研究中发酵罐培养时,选用高浓度氨基酸发酵,可以改善菌体生长状况,从而提高发酵液中青蒿二烯的浓度。
(4)发酵罐培养时,选择较大转速和较高空气通量,能改善菌体生长状况,增强其生产青蒿二烯的能力。
[1]Wallaart T E, Pras N, Qua X W J. Seasonal variations of artemisinin and its biosynthetic precursors intetraploidArtemisia annuaplants compared with the diploid wild-type [J].Plant Medica, 1999, 65(8):723-728
[2]Xu Xingxiang, Zhu Jie, Huang Dazhong, Zhou Weishan.Totalsynthesis of arteannuin and deoxyarteannui [J].Tetrahedron,1986, 42(3): 819 -828
[3]Kong Jianqiang(孔建强), Wang Wei(王伟), Cheng Kedi(程克棣),Zhu Ping(朱平). Research progress in synthetic biology of artemisinin [J].Acta Pharmaceutiaca Sinica(药学学报), 2013, 48(2):193-205
[4]Liu Duo(刘夺), Du Jin(杜瑾), Zhao Guangrong(赵广荣), Yuan Yingjin(元英进). Applications of synthetic biology in medicine and energy [J].CIESC Journal(化工学报), 2011, 62(9): 2391-2397
[5]Schramek N, Wang H, RömischMargl W, Keil B, Radykewicz T,Winzenhörlein B, Beerhues L, Bacher A, Rohdich F, Gershenzon J,Liu B, Eisenreich W. Artemisinin biosynthesis in growing plants ofArtemisia annua. A13CO2study [J].Phytochemistry, 2010, 71(2-3):179-187
[6]Bouwmeestera H J, Wallaartb T E, Janssena M H A, Loo B V, Jansen B J M, Posthumus M A, Schmidt C O, Kraker J W D, König W A,Franssen M C R. Amorpha-4,11-diene synthase catalyses the first probable step in artemisinin biosynthesis [J].Phytochemistry, 1999,52(5): 843-854
[7]Kuzuyama T, Seto H. Diversity of biosynthesis of the isoprene units[J].Natural Product Reports, 2003, 20: 171-183
[8]Paddon C J, Keasling J D. Semi-synthetic artemisinin: a model for the use of synthetic biology in pharmaceutical development [J].Nature Reviews, 2014, 12(5): 335-367
[9]Martin V J, Pitera D J, Withers S T, Newman J D, Keasling J D.Engineering a mevalonate pathway inEscherichia colifor production of terpenoids [J].Nature Biotechnology, 2003, 21(7): 796-802
[10]Ro D K, Paradise E M, Ouellet M, Fisher K J, Newman K L,Ndungu J M, Ho K A, Eachus R A, Ham T S, Kirby J, Chang C Y M,Withers S T, Shiba Y, Sarpong R, Keasling J D. Production of the antimalarial drug precursor artemisinic acid in engineered yeast [J].Nature, 2006, 440(7086): 940-943
[11]Tsuruta H, Paddon C J, Eng D, Lenihan J R, Horning T, Anthony L C,Regentin R, Keasling J D, Renninger N S, Newman J D. High-level production of amorpha-4,11-diene, a precursor of the antimalarial agent artemisinin, inEscherichia coli[J].PLoS ONE,2009, 4(2):e4489
[12]Kong Jianqiang(孔建强), Shen Junhao(沈君豪), Huang Yong( 黄勇),Wang Wei(王伟), Cheng Kedi(程克棣), Zhu Ping(朱平). Production of amorpha-4,11-diene in engineered yeasts [J].Acta Pharmaceutiaca Sinica(药学学报), 2009, 44(11): 1297-1303
[13]Kong Jianqiang, Wang Wei, Wang Lina, Zheng Xiaodong, Cheng Kedi, Zhu Ping. The improvement of amorpha-4,11-diene production by a yeast-conform variant of vitreoscillahemoglobin [J].Journal of Applied Microbiology, 2009, 106(3): 941-951
[14]Westfall P J, Pitera D J, Leihan J R, Eng D, Woolard F X, Regentin R,Horning T, Tsuruta H, MelisD J, Owens A, Fickes S, Diola D,Benjamin K R, Keasling J D, Leavell M D, McPhee D J, Renninger N S, Newman J D, Paddon C J. Production of amorphadiene in yeast,and its conversion to dihydroartemisinic acid, precursor to the antimalarial agent artemisinin [J].PNAS, 2012, 109(3): 111-118
[15]Wang Sijia(王思佳), Ding Mingzhu(丁明珠), Yuan Yingjin(元英进).The impact of promoters and vectors on the amorphadiene production in yeast cells [J].China Biotechnology(中国生物工程杂志), 2013,33(8): 15-23
[16]Jia Yunjing(贾云婧), Zhao Juan(赵娟), Ding Mingzhu(丁明珠),Yuan Yingjin(元英进). Fitness of amorphadiene production functional modules and yeast chassis [J].Chemical Journal of Chinese Universities(高等学校化学学报), 2013, 34(12): 2765-2771
[17]Lei Wei(雷桅), Tang Shaohu(汤绍虎), Zhou Qigui(周启贵), Shui Xiaorong(税晓容), Sun Yiming(孙一鸣), Sun Min(孙敏).Bioinformatics analysis of 3-hydroxy-3-methylglutaryl-coA reductase(HMGR)in isoprenoid biosynthesis of mulberry [J].Science of Sericulture(蚕业科学), 2008, 34(3): 4609-4614
[18]Donald K A, Hampton R Y, Fritz I B. Effects of overproduction of the catalytic domain of 3-hydroxy-3-methylglutaryl coenzyme A reductase on a qualene synthesis inSaccharomyces cerevisiae[J].Applied and Environmental Microbiology, 1997, 63(9): 3341-3344
[19]Han Junli, Liu Benye,Ye Hechun, Wang Hong, Li Zhenqiu, Li Guofeng. Effects of overexpression of the endogenous farnesyldiphosphate synthase on the artemisinin content inArtemisia annua[J].Journal of Integrative Plant Biology, 2006, 48(4): 482-487
[20]Paradise E M, Kirby J, Chan R, Keasling J D. Redirection of flux through the FPP branch-point inSaccharomyces cerevisiaeby down-regulating squalene synthase [J].Biotechnology and Bioengineering, 2008, 100(2): 371-378
[21]Pitera D J, Paddon C J, Newman J D, Keasling J D. Balancing a heterologous mevalonate pathway for imprive disoprenoid production inEscherichia coli[J].Metabolic Engineering, 2007, 9: 193-207
[22]Sakai A, Shimizu Y, Hishinuma F. Integration of heterologous genes into the chromosome ofSaccharomyces cerevisiaeusing a delta sequence of yeast retrotansposon Ty [J].Applied Microbiology and Biotechnology, 1990, 33(3): 302-306
[23]Lee Frank W F, SilvaNancy A D. Sequential Delta-integration for the regulated insertion of cloned genes inSaccharomyce scerevisiae[J].Biotechnology Progress, 1997, 13(4): 368-373
[24]Tianjin University. A standardized, high accuracy and general method for build of function module[P]: CN, 201310377238.1. 2013-08-27
[25]van Dijken J P, Bauer J, Brambilla L, Duboc P, Francois J M,Gancedo C, Giuseppin M L, Heijnen J J, Hoare M, Lange H C,Madden E A, Niederberger P, Nielsen J, Parrou J L, Petit T, Porro D,Reuss M, van Riel N, Rizzi M, Steensma H Y, Verrips C T, Vindelov J, Pronk J T. An interlaboratory comparison of physiological and geneticproperties of fourSaccharomyces cerevisiaestrains [J].Enzyme Microbiology Technology, 2000, 26: 706–714
[26]BenAri G, Zenvirth D, Sherman A, David L, Klutstein M, Lavi U,Hillel J. Four linked genes participate in controlling sporulation efficiency in budding yeast [J].PLoS Genetics, 2006, 2(11): e195
[27]Albers E, Larsson C, Lidén G, Niklasson C, Gustafsson L. Influence of the nitrogen source onSaccharomyces cerevisiaeanaerobic growth and product formation [J].Applied and Environmental Microbiology,1996, 62(9): 3187-3195
[28]Pham H T B, Larsson G, Enfors S O. Growth and energy metabolism in aerobic fed-batch cultures ofSaccharomyces cerevisiae: simulation and model verification [J].Biotechnology and Bioengineering, 1998,60(4): 474-482
[29]Watson T G. Amino-acid pool composition ofSaccharomyces cerevisiaeas a function of growth rate and amino-acid nitrogen source [J].Journal of General Microbiology, 1976, 96: 263-268
