山西省西伯利亚远志种质资源的ISSR分析①
2015-06-18刘超等
刘超等
摘 要 应用ISSR标记对山西省7个野生居群和1个人工栽培居群共60份西伯利亚远志种质进行遗传多样性分析。结果表明:从100对ISSR引物中共筛选出20条多态性好、条带稳定的引物,60份材料DNA共获得338个扩增位点,其中多态性位点334个,多态性比率为98.82%;有效等位基因数(Ne)、Nei's基因多样性指数(H)及Shannon多样性信息指数(I)分别为1.433 7、0.265 6和0.414 2,居群的遗传相似度在0.655 2~0.977 9,表明8个居群具有丰富的遗传多样性,这与其自身繁殖特点有关;遗传分化系数(Gst)和基因流(Nm)分别为0.526 0和0.450 5,表明居群内和居群间均有一定程度的分化,基因流水平较低;利用UPGMA法进行聚类分析,将8个居群划分为3类。研究结果为合理引种、驯化、保护和利用西伯利亚远志野生资源提供了重要的参考依据和数据支持。
关键词 西伯利亚远志 ;ISSR ;种质资源 ;遗传多样性 ;聚类分析
分类号 Q949.753.3
西伯利亚远志(Polygala sibirica L.)为远志科(Polygalaceae)远志属(Polygala L.)植物,是多年生草本,高10~30 cm,叶片下部小的为卵形,上部大的为披针形或椭圆状披针形;总状花序假顶生或腋外生;花为蓝紫色,龙骨瓣较长,具流苏状鸡冠状附属物;蒴果呈倒心形。全国各地均有该物种的分布。
中药远志为中国大宗常用药材,中华人民共和国卫生部药典委员会于1977年首次将西伯利亚远志P. sibirica L.列入《中国药典》[1],规定其和远志P. tenuifolia Willd.同为药用远志的2种基源植物,主治心肾不交所引起的失眠多梦、健忘惊悸、神志恍惚、咳痰不爽、疮疡肿毒及乳房肿痛等。关于药用远志来源的争议较多,王光志等[2]于2008年对药物远志进行了比较全面的本草考证,通过用药历史与名称、品种的考证,发现虽然药材品种较多,但远志古今用药品种基本一致,均为远志P. tenuifolia Willd.与西伯利亚远志P. sibirica L.。远志最初来源于山东半岛一带,古代本草以山西、河南为道地,今以山西、陕西的质量最好,产量最大,为道地产地。由于市场近年来对远志药材的需求量剧增,导致野生资源遭到了极大破坏,远志现已被列入国家重点保护的三级野生品种中[3]。中药远志研究主要集中在化学成分的分离鉴定和药理作用方面,而关于遗传多样性的研究报道较少。
李佳等[4]曾对主产区远志P. tenuifolia Willd.的种质资源遗传多样性进行了ISSR分析,包括野生种和栽培种,探索野生和栽培环境对远志遗传多样性的影响。ISSR分子标记为第二代分子标记技术,克服了RAPD标记稳定性和重复性差等缺点[5],且ISSR标记多态性丰富,已被广泛应用于品种鉴定、多样性分析、指纹图谱的构建等研究中[6]。
目前对药用远志的研究主要集中在远志P.tenuifolia Willd.这个种,而对西伯利亚远志的研究较少。本研究利用ISSR技术对西伯利亚远志道地产地之一的山西省居群进行分析,从分子水平上探讨其种质资源的遗传多样性和亲缘关系,旨在为其种质资源的开发利用及遗传育种研究提供一定的依据。
1 材料与方法
1.1 材料
1.1.1 植物材料
实验材料分别采自山西省不同地区的8个居群,包括7个野生居群和1个人工栽培居群,居群代号见表1。经过云南大学马海英副教授鉴定可知这8个居群均为西伯利亚远志P. sibirica L.。其中太原市TZ居群10份种质;长治市CZT 和CZY居群各5份种质;宁武县NWI和NWII居群各10份种质;太原古交市TG居群5份种质;运城市YC居群5份种质;吕梁市关帝山GDS居群10份种质等。共计60份种质,其地理信息见图1。
1.1.2 仪器设备
紫外分光光度计;Biometra高性能PCR仪(德国Analytik Jena公司生产);紫外凝胶成像仪等。
1.2 方法
1.2.1 基因组DNA的提取及检测
参照Doyle等[7]的 CTAB法,稍作改良(加入CTAB前先加入去多糖Buffer去除多糖)。称取0.03 g干燥的植物叶片进行DNA提取,用50 μL的0.1×TE溶液进行溶解,在1%的琼脂糖凝胶电泳下检测其完整性,用紫外分光光度计测定DNA的浓度和纯度,用0.1×TE溶液稀释至20 ng/μL后于-20℃冰箱中保存。
1.2.2 引物筛选及PCR扩增检测
根据加拿大大不列颠哥伦比亚大学(UBC)公布的100对ISSR引物序列进行引物合成,由上海生工生物工程技术服务有限公司合成。从8个居群中各选取2个样品的DNA模板进行扩增筛选,其中有33条引物能扩增出条带,对每条引物进行退火温度筛选,其中条带清晰、条数较多且重复性较好的引物共计20条(表1)。PCR扩增反应在德国Analytik Jena公司生产的Biometra 高性能PCR仪中进行。最终确定的反应体系为25 μL,包括20 ng/μL的DNA、10×buffer(含MgCl2)、2.5 mmol/L 的dNTPs、5 U/μL的Easy Taq DNA聚合酶,最后以去离子水补足至25 μL。PCR扩增程序为40个循环:95℃变性30 s;最适温度(根据每条引物的理论退火温度再进行梯度PCR确定,部分种质退火温度见图2和图3)退火30 s;72℃延伸90 s;循环结束后以72℃延伸10 min。扩增产物经由0.5×TBE溶液配置的2.0%的琼脂糖凝胶电泳分离,在90 V电压下于冰面上电泳1.5 h,用EB(溴化乙锭)染色后将其置于紫外凝胶成像仪中观察并保存图像。
1.3 数据统计与分析
ISSR标记为一种显性标记,图谱中的每一条带都视为1个分子标记,有清晰条带者记为1,同一位置无带者记为0,采用人工计数方法根据统计数据建立(0,1)矩阵。利用POPGENE 1.32软件分析计算多种群遗传多样性和遗传分化等指数,包括Nei's基因多样性指数(H)、Shannon信息指数(I)、多态位点百分率(PPL)、有效等位基因数(Ne)、居群内基因多样性(Hs)、居群间遗传分化系数(Gst)、基因流(Nm)、Nei's基因遗传距离等。应用GenAlEx6.41软件对种群内和种群间进行AMOVA分析和主成分分析。利用NTSYS2.1聚类分析软件进行UPGMA法聚类分析。
2 结果与分析
2.1 西伯利亚远志的遗传多样性分析
实验中共筛选出20条引物,扩增出338个位点,其中多态性位点达到334个。从表2可以看出,多态性位点百分率(PPL)为98.82%。绝大多数扩增片段大小在250~2 000 bp,部分电泳片段见图4。8个居群中CZY居群多态性最高,其多态位点为243个,多态率为71.89%;多态性最低的为TG居群,其多态位点为38个,多态率为11.24%;栽培居群TZ的多态位点为163个,多态率为48.22%。用POPGENE1.32软件分析得到的西伯利亚远志的多样性指数见表2,Shanoon's指数(I)为0.4142;Nei's遗传多样性(H)为0.265 6。通过多态性位点百分率(PPL)、遗传多样性(H)和Shanoon's指数(I)分析可知,8个居群中多态性高低依次为CZY>CZT>TZ>NWI>NWII>GDS>YC>TG。
2.2 西伯利亚远志的遗传结构分析
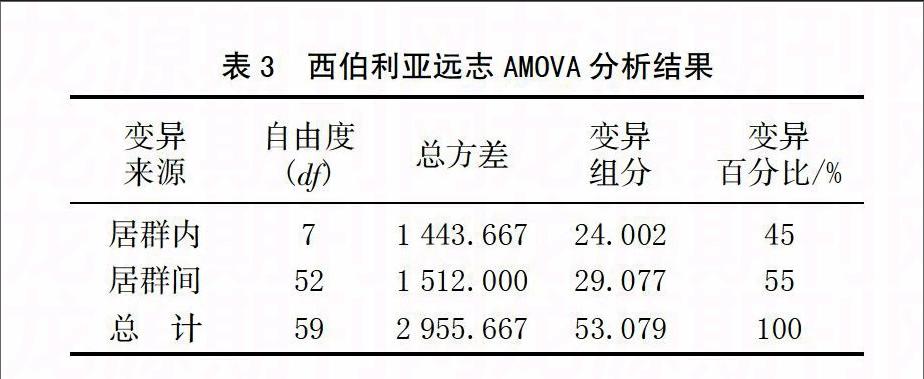
通过POPGENE 1.32软件分析可知,西伯利亚远志居群内基因多样度(Ht)为0.275 9,基因多样性(Hs)为0.1307,遗传分化系数(Gst)为0.526 0,Nm为0.450 5。另外,通过利用GenAlex6.41软件进行AMOVA分析(P=0.010)可知,居群间遗传变异占45%,居群内遗传变异占55%(表3)。
利用POPGENE 1.32软件运算8个居群中NWI和NWII居群的遗传距离最近为0.022 4,CZT和TG居群的遗传距离最远为0.422 8(表4)。不同居群的遗传相似度(即遗传一致度)在0.655 2~0.977 9,平均值为0.806 2。
2.3 西伯利亚远志的聚类分析
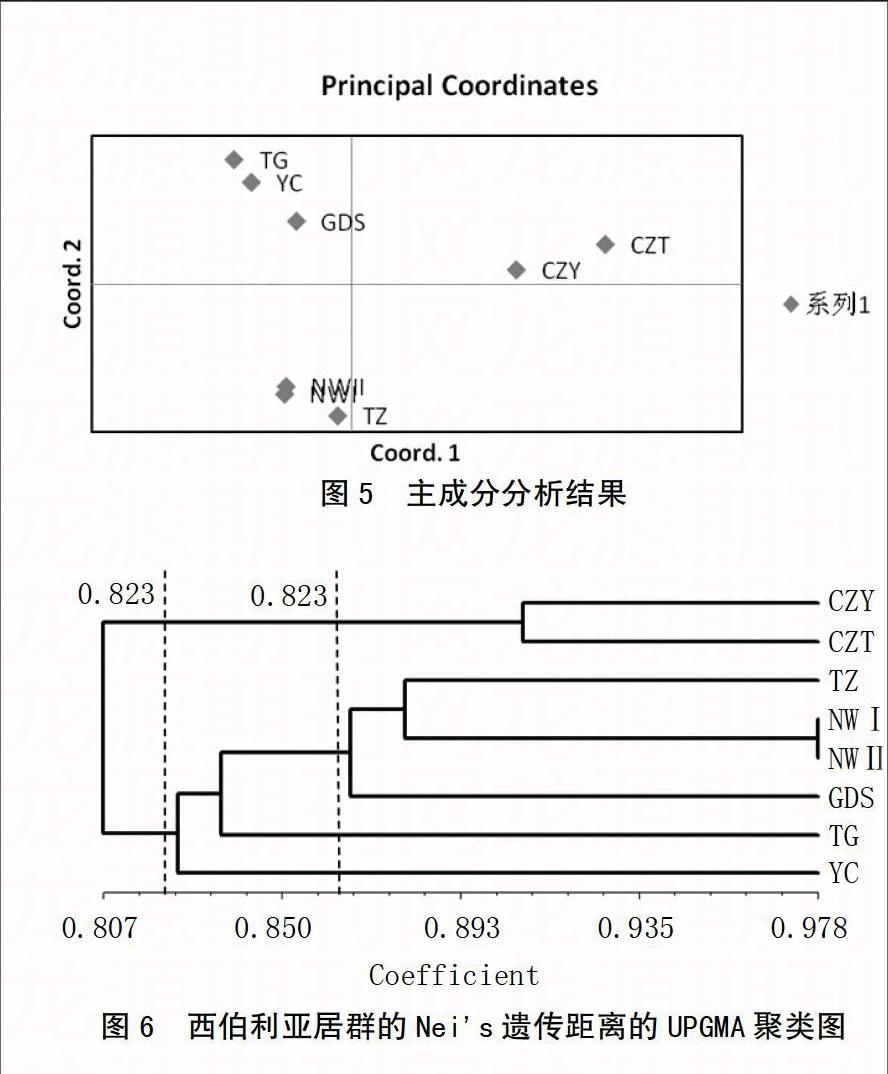
用GenAlex6.41软件构建欧式平方距离,在此基础上对8个居群进行主成分分析,以此检测居群的遗传相似性(图5)。主成分分析可排除重叠和分类意义不大的条带,使重要因素突显出来。主成分分析结果表明,长治的2个野生居群CZY和CZT聚为一类;宁武的2个野生居群NWI和 NWII以及太原的栽培居群TZ聚为一类;太原古交的居群TG、运城YC居群和吕梁GDS居群聚为一类。
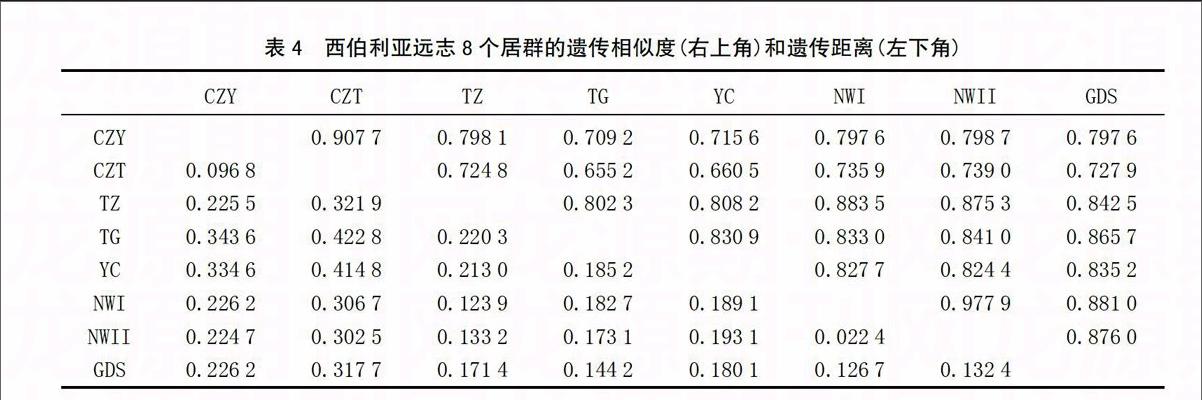
再利用 NTSYS2.1聚类分析软件进行UPGMA法聚类分析,建立聚类分支树状图(图6)。 8个居群种质在0.823和0.876处共分为3类,CZY和CZT居群为一类;NWI、 NWII和TZ居群为一类; GDS、TG和YC居群为一类。其中CZY和CZT居群与其他居群距离最远,聚类分析结果与主成分分析结果基本相同。
3 讨论与结论
3.1 西伯利亚远志的遗传多样性
本研究结果显示,山西省西伯利亚远志居群具有丰富的遗传多样性(H=0.265 6,I=0.414 2,PPL=98.82%)。遗传多样性是生物在长期进化和生长过程中所形成的自然属性,受多方面因素的影响,如生殖方式、生活型、分布范围、花粉、种子传播方式以及演化历史等[8]。一个物种的遗传多样性与其生活史特征和生态特征密切相关,分布范围广泛是物种遗传多样性高的主要因素;其次物种繁殖方式对其影响也较大[9-10],相关学者普遍认为多年生广布种、异交和以种子为传播方式等均会使植物具有较高的遗传多样性。
西伯利亚远志在中国均有分布,此外还分布于欧洲、俄罗斯西伯利亚、尼泊尔、印度、蒙古和朝鲜等地[11],分布范围广泛。西伯利亚远志为多年生草本,花粉形态和雌雄蕊位置都表现出明显的虫煤传粉特征,且自交不育[12],这些特征都导致其具有较高的遗传多样性。本研究发现,栽培种居群TZ遗传多样性亦较高(H=0.151 4,I=0.230 9,PPL=48.22%),是因为此栽培种是由直接采自野生居群的种子繁育而成的,人工露地种植环境相当于半野生状态。
3.2 西伯利亚远志的遗传分化
西伯利亚远志居群的Gst为0.526 0,表明居群间遗传变异为52.60%;AMOVA分析结果表明,居群间的遗传变异为45%。2种方法分析结果虽然有所差异,但都表明西伯利亚远志种质居群间出现了一定程度的遗传分化。分析认为主要是由于基因流降低而造成的,基因流是影响群体间遗传分化的主要因素[13]。本研究中西伯利亚远志的基因流(Nm)为0.450 5,当居群的基因流小于1时,表示遗传漂变导致了居群间的遗传分化[14]。分析认为,西伯利亚远志基因流较低的原因是野生居群生态环境遭到严重破坏导致的。西伯利亚远志本身生长周期长、产量低、生长分散,近年来的过度采挖造成其生长环境的严重破坏,其密度和再生能力明显减弱,进而使生境片段化,导致野生居群整体基因流降低,造成了居群间遗传分化的加剧。
根据聚类分析和主成分分析结果将西伯利亚远志居群分为3类,这与各居群间的地理距离有一定关系。其中太原市的栽培居群TZ和宁武县的2个野生居群NWI和NWII聚为一类,表现出较大的地域分布差异性,TZ居群是由野生居群引种栽培而来的,这可能是其聚类结果与其地理原产地之间没有相关性的主要原因。国外学者Thimmappaiah等[15]对栽培植物腰果(Anacardium occidentale)的研究也证实了这一点,发现产生这一结果的主要原因是来自不同地方的种质已经进行了基因的自由交流。
3.3 西伯利亚远志的保育策略
山西省为西伯利亚远志的道地产地之一,其与陕西的总产量占全国70%以上[16]。但如今野生西伯利亚远志资源已经难以满足市场需求,资源逐年锐减,有些地方甚至已经濒临灭绝。而人工栽培种子来源复杂,加上其生长周期较长,对栽培技术要求高,难以形成规模化栽培,不能满足市场需求[17]。
野生西伯利亚远志资源具有丰富的遗传变异,是培育优良品种的遗传物质基础。可对野生资源进行就地保护和采集种源,用以建立种质资源圃,通过育种方式形成栽培品种,筛选出适应不同生态环境的优良品种,以此逐步替代野生资源,当条件成熟时也可对野生资源进行恢复和重建。同时提高人工栽培技术水平,建立一套完善优质的远志栽培技术体系,实现西伯利亚远志资源的可持续利用。
参考文献
[1] 中华人民共和国卫生部药典委员会. 中国药典I部[S]. 北京:人民卫生出版社,1977: 265-266.
[2] 王光志,万德光. 远志的本草考证[EB/OL].[2008-02-03]. http://www.paper.edu.cn/releasepaper/content/200802-36.
[3] 李世全. 中药采购技术手册[M]. 西安:陕西科学技术出版社,1998:411-412.
[4] 李 佳,房敏峰,周天华,等. 主产区远志种质资源遗传多样性的ISSR分析[J]. 中草药,2010,41(11):1 881-1 885.
[5] 白 玉. DNA分子标记技术及其应用[J]. 安徽农业科学, 2007, 35(24): 7 422-7 424.
[6] 蒋彩虹,王元英,孙玉合. SSR和ISSR标记技术应用进展[J]. 中国烟草科学,2007,28(2):1-5.
[7] Doyle J J, Doyle J L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue[J]. Phytochemical Bulletin, 1987, 19: 11-15.
[8] Soltis P S, Soltis D E. Genetic variation in endemic and widespread plant species: examples from Saxifragaceae and Polystichum[J]. Aliso, 1991, 13: 215-223.
[9] 钱迎倩,马克平. 生物多样性研究的原理和方法[J]. 北京:中国科学技术出版社,1994:123-137.
[10] Hamrick J L, Loveless M D. Factors influencing levels of genetic diversity in woody plant species[J]. New Forests, 1992, 6(1-4): 95-124.
[11] 中国科学院植物志编委会. 中国植物志.第43卷.第3册[M]. 北京:科学出版社,1997:193-195.
[12] 滕红梅. 药用远志的结构发育与主要药用成分积累关系的研究[D]. 陕西:西北大学,2009.
[13] 邹喻苹,葛 颂,王晓东. 系统与进化植物学中的分子标记[M]. 北京:科学出版社,2001:16-17.
[14] Brown A H D, Clegg M T, Kahler A L, et al. Plant population genetics, breeding, and genetic resources[M]. Sunderland, MA, USA: Sinauer, 1989: 43-263.
[15] Thimmappaiah, Santhosh W G, Shobha D, et al. Assessment of genetic diversity in cashew germplasm using RAPD and ISSR markers[J]. Scientia Horticulturae, 2009, 120(3): 411-417.
[16] 张丽萍. 远志[M]. 北京:中国中医药出版社,2001:1-2.
[17] 赵云生,万德光,严铸云,等. 远志资源生产现状调查[J]. 亚太传统医药,2014,10(24):1-3.
