多杀性巴氏杆菌HN06基因组分泌蛋白的预测与分析
2015-06-11刘文静徐卓菲陈焕春
彭 忠,梁 婉,2,刘文静,徐卓菲,谭 臣,吴 斌*,周 锐,陈焕春
(1.华中农业大学动物医学院农业微生物学国家重点实验室,生猪健康养殖协同创新中心,湖北武汉430070;2.华中农业大学动物科学技术学院农业动物遗传育种与繁殖教育部重点实验室,生猪健康养殖协同创新中心,湖北武汉430070)
多杀性巴氏杆菌是一种严重危害畜牧业的重要的革兰阴性病原菌,对多种畜禽及野生动物甚至人都能造成不同程度的感染和致病[1-2]。依据荚膜抗原的不同,多杀性巴氏杆菌常被分为A、B、D、E、F 5种血清型[3]。临床调查发现,不同血清型的多杀性巴氏杆菌表现出一定的宿主及致病偏好性[2]。多杀性巴氏杆菌的毒力因子众多,目前已知的毒力因子主要包括荚膜、脂多糖(LPS)、黏附因子、巴氏杆菌毒素(PMT)、铁摄取相关蛋白、唾液酸代谢、透明质酸酶以及外膜蛋白(OMP)[4]。
有研究发现[5-6],革兰阴性菌中1/3的蛋白合成后需要被转运或分泌到细胞外发挥作用,包括某些水解酶类、细胞毒素、胶质类、生长因子、激素类或抗体等,这类蛋白被称为分泌蛋白。分泌蛋白一般占细菌基因组编码蛋白总量的10%~20%,它们不仅参与细胞信号转导,而且对于细胞增值、分化及凋亡都具有十分重要的调控作用。因此,研究细菌的分泌蛋白对于了解细菌的生命周期及其致病机理以及病原菌与宿主间的相互作用具有十分重要的意义。然而,由于分泌蛋白不易被高浓度提取,这对于开展分泌蛋白研究产生了较大的阻碍。
随着全基因组测序技术及生物信息学的发展,特别是细菌全基因组序列的测定和公布及配套生物信息学工具的开发为细菌分泌蛋白的预测和功能研究提供了巨大的帮助。自2001年分离于美国的禽源多杀性巴氏杆菌Pm70株的全基因组测序被报道后[7],最近几年又有几株完整的多杀性巴氏杆菌全基因组序列被公布,其中的多杀性巴氏杆菌HN06株是一株来源于中国海南地区的D 型产毒素巴氏杆菌,分离自一头患有萎缩性鼻炎病猪的鼻腔,其全基因组序列的测序于2012年完成并公布[8],是世界上第1株完成全基因组序列测定的产毒素多杀性巴氏杆菌。本研究将利用Signal P、Tmhmm、Phobius、Target P、Lipop、GPI-SOM、Tat P、SecretomeP 等生物信息学工具预测出其分泌蛋白[9],并将这些蛋白与其他3株多杀性巴氏杆菌(Pm70、3480、36950)的分泌蛋白进行比较分析,筛选出4株菌株共有的经典途径Sec分泌蛋白,并对其进行了初步的功能分析,为进一步筛选多杀性巴氏杆菌亚单位疫苗候选蛋白奠定基础。
1 材料与方法
1.1 材料
多杀性巴氏杆菌Pm70、3480、36950 及HN06的全基因组信息均下载于GenBank(http://www.ncbi.nlm.nih.gov/genome/genomes/912);分泌蛋白预测及分析工具包括Signal P4.1(http://www.cbs.dtu.dk/services/SignalP/);Tmhmm v 2.0(http://www.cbs.dtu.dk/services/Tmhmm/);Phobius(http://phobius.sbc.su.se/);Target P1.1(http://www.cbs.dtu.dk/services/TargetP/);Lipo P1.0(http://www.cbs.dtu.dk/services/LipoP/);GPI-SOM(http://gpi.unibe.ch/)以及SecretomeP 2.0(http://www.cbs.dtu.dk/services/SecretomeP/)。
1.2 方法
1.2.1 Sec途径分泌蛋白的筛选 分泌蛋白的氨基酸序列的N 端通常有一段10个~50个氨基酸的疏水性片段即信号肽[6],用以引导随后的蛋白质多肽链穿过内质网膜到达腔内,完成后,一些信号肽序列被信号肽酶切除,释放出成熟的分泌蛋白。因此蛋白质序列N 端是否具有信号肽可以作为一个重要的预测依据。对Pm70、3480、36950及HN06等4个基因组的蛋白序列用Signal P4.1[10],进行信号肽的预测,保留具有信号肽的蛋白。使用Tmhmm v2.0[11]筛选出具有0或1个跨膜螺旋的蛋白,由于该软件在判断具有1个跨膜螺旋的蛋白时,不能准确区分信号肽和跨膜区,因此需要使用Phobius软件重新预测具有1个跨膜螺旋的蛋白,剔除确实具有跨膜区的蛋白[12]。再利用GPI-SOM 判断上述筛选的蛋白中是否具有锚定位点,剔除锚定蛋白。最后,采用Target P1.1软件预测蛋白的亚细胞定位,进一步确定该蛋白的分布类型,保留分泌型蛋白。上述筛选的蛋白在LipoP 软件的预测下被分为4类,即被Ⅰ型信号肽酶识别的蛋白、被Ⅱ型信号肽酶识别的蛋白、细胞质蛋白和跨膜蛋白。
综上所述,具有信号肽、不含跨膜螺旋、无GPI锚定位点、通过分泌途径分泌且能被Ⅰ型和Ⅱ型信号肽酶识别的蛋白可初步预测为Sec分泌蛋白[13]。
1.2.2 Tat途径分泌蛋白的筛选 使用Tat P 软件筛选出具有RR 信号肽且D 值大于默认阀值0.36的蛋白,然后使用Tmhmmv 2.0和Phobius筛选出其中无跨膜螺旋结构的蛋白。
1.2.3 无信号肽的非经典分泌蛋白的筛选 用Signal P4.1筛选出没有信号肽的蛋白,并用SecretomeP 2.0 软 件 选 取secp 值 大 于0.5 的 序 列。经Tmhmmv 2.0和Phobius筛选,保留无信号肽、secp值大于0.5的无跨膜螺旋蛋白[14]。
2 结果
2.1 基因组和蛋白组信息搜集
多杀性巴氏杆菌Pm70、3480、36950 及HN06这4株多杀性巴氏杆菌的全基因组序列及蛋白质序列均来自于美国国家生物技术信息中心(NCBI),GenBank登 录 号 分 别 为:NC-002663、NC-017764、NC-016808、NC-017027;其中HN06株的全基因组序列和蛋白质序列由华中农业大学农业微生物学国家重点实验室(湖北,武汉)提交。
2.2 多杀性巴氏杆菌HN06株分泌蛋白预测
2.2.1 Sec途径分泌蛋白预测结果 使用Signal P4.1预测到多杀性巴氏杆菌HN06株含有179个具有信号肽的蛋白,进一步利用Tmhmmv 2.0软件预测到这179个蛋白包括150个无跨膜螺旋的蛋白和22个只有1个跨膜螺旋的蛋白;经Phobius软件检查发现,具有1个跨膜螺旋的22个蛋白中有2个蛋白具有跨膜区。此外,使用GPI-SOM 软件预测出10个锚定蛋白,排除后采用Target P 软件确定其亚细胞定位,筛选出分泌型蛋白157个。最后,用LipoP 软件筛选出能被Ⅰ型信号肽酶识别的蛋白120个,能被Ⅱ型信号肽酶识别的蛋白37个。
2.2.2 Tat途径分泌蛋白预测结果 经Tat P 分析,共预测到HN06株中Tat途径分泌蛋白8个。
2.2.3 非经典途径分泌蛋白预测结果 HN06 株非经典途径分泌蛋白共预测到208个。
2.3 多杀性巴氏杆菌HN06株分泌蛋白组成分析
多杀性巴氏杆菌全基因组共编码2 265个开放阅读框(open reading frame,ORF),经预测其中含有373个分泌蛋白,所占比例为16.5%;这些分泌蛋白中spⅠ、spⅡ、Tat P、非经典途径分泌蛋白比例见图1;就氨基酸就成个数而言,373个分泌蛋白中氨基酸数量最小为36 个,最大为2 987个,绝大多数蛋白质的大小皆位于0~350 个氨基酸残基之间,占分泌蛋白总数的71.1%(图2)。
信号肽是Ⅰ型和Ⅱ型分泌蛋白转运过程中必不可少的一个元件,我们对其长度进行分析发现Ⅰ型分泌蛋白的长度分布在18 个~33 个氨基酸之间,其中19个~26个氨基酸的信号肽最多,占91.7%,Ⅱ型分泌蛋白的长度分布在15个~25个氨基酸之间,其中18个~19 个氨基酸的信号肽最多,占59.5%(图3)。
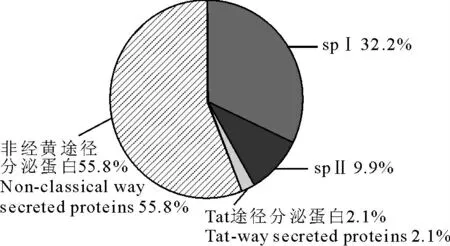
图1 HN06株分泌蛋白组成分布图Fig.1 Pie chart of the distribution of HN06Sec proteins

图2 HN06株分泌蛋白的ORF长度分布Fig.2 Distribution of the Sec protein ORF length of HN06
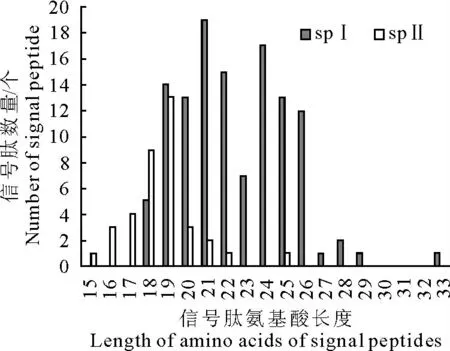
图3 Ⅰ型和Ⅱ型分泌蛋白的信号肽长度分布Fig.3 Distribution of signal peptide length of the Sec proteins recognized by signal peptidaseⅠandⅡ
Tat途径分泌蛋白的信号肽N 端含有一段RRmotif保守结构区,因此这种信号肽又称RR-motif型信号肽,通常组成为RR-X-(FGAVML)-(LITMVF)[15],X 为任意氨基酸。本研究对Tat途径分泌蛋白的RR-motif信号肽进行分析发现,HN06中预测到的8个RR-motif均符合一般组成规律(表1)。
2.4 HN06分泌蛋白功能分析
2.4.1 Sec途径分泌蛋白功能分析 根据HN06株蛋白注释信息及功能不明确蛋白的Blastp发现预测到的120 个spⅠ型分泌蛋白中假设蛋白26个,所占比例21.7%。其余蛋白的功能主要是细胞壁合成酶、代谢相关酶类、水解酶及代谢相关蛋白。代谢相关蛋白中除ABC 转运蛋白、磷酸结合蛋白外,还发现有7个铁代谢相关蛋白、4个血红蛋白结合蛋白及4个TonB 依赖性蛋白,这些都是病原菌铁摄取途径必须的蛋白。

表1 HN06株具有RR-motif的8个信号肽Table 1 8RR-motif signal peptides in Pasteurella multocida HN06
spⅡ型分泌蛋白又称脂蛋白,在对其功能分析发现,假设蛋白11 个,所占比例30%,其余蛋白主要是代谢酶类和代谢相关蛋白。
2.4.2 Tat途径分泌蛋白功能分析 预测到的8个Tat途径分泌蛋白中有5个还原酶,1个代谢相关的脱氢酶,1个磷酸二酯酶及1个细胞分裂蛋白。
2.4.3 非经典途径分泌蛋白 HN06株中共预测到非经典途径分泌蛋白208 个,其中假设蛋白84个,所占比例40.4%。其余蛋白中DNA 复制相关蛋白和酶占16.3%,代谢酶类占9.6%,氧化还原酶类占6.3%,另外还有7个噬菌体相关蛋白、2个血红素结合受体及热击蛋白、冷击蛋白及黏附素等。
2.5 四株多杀性巴氏杆菌共有sec途径分泌蛋白功能分析
对其他3株巴氏杆菌使用同样的方法预测出其sec途径分泌蛋白,并对这些分泌蛋白序列使用Blastp进行比对,共得到203个sec途径分泌蛋白。各株菌所包含的sec途径分泌蛋白的分布情况见图4,四株菌共有的sec途径分泌蛋白共114个,占总数的56.2%,至少在两株菌中存在的sec途径分泌蛋白共69个,占总数的34.0%,剩余的9.80%为各个菌株所特有。
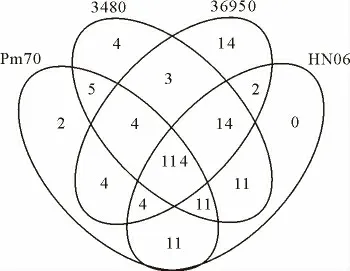
图4 Pm70、3480、36950及HN06株Sec分泌蛋白分布韦恩示意图Fig.4 Venn diagram of the distribution of Sec proteins of Pm70,3480,36950and HN06strains
对这4株巴氏杆菌预测到的共有Sec分泌蛋白的功能分析发现,有13个功能未知的假设蛋白,所占比例为11.4%,其余的蛋白中代谢相关蛋白占39.5%,代谢酶类占9.65%,其中代谢相关蛋白中有31.1%是铁代谢、血红蛋白结合蛋白和TonB 依赖性蛋白。
3 讨论
生物信息学的发展使得人们能够将其广泛的应用于科学研究中,更快速有效的获取信息。但是不同的软件有着不同的计算方式,预测的结果也不尽相同,甚至会有预测错误的现象出现,进一步验证实验十分必要。
在本次研究中,预测到巴氏杆菌HN06株分泌蛋白共373个,其中Sec分泌蛋白157个,Tat途径分泌蛋白8 个,无信号肽的非经典途径分泌蛋白208个。Sec分泌蛋白被Ⅰ型信号肽酶识别的有120个,被Ⅱ型信号肽酶识别的有37个。在对其功能分析后发现,Sec分泌蛋白中代谢相关蛋白和酶类及细胞壁水解和合成相关酶类较多,Tat途径分泌蛋白中还原酶较多,而非经典途径分泌蛋白中DNA 复制相关蛋白和酶类较多。Pm70、3480、36950及HN06这4株巴氏杆菌共有的Sec分泌蛋白中代谢相关蛋白较多,并且铁代谢、血红蛋白结合蛋白和TonB依赖性蛋白占有较大比例。
研究发现,铁是病原菌生长所必须的微量营养元素,多杀性巴氏杆菌中铁代谢相关基因占全基因组的2.5%。细菌通过铁载体、转铁蛋白和血红蛋白等多种螯合系统调控铁的代谢,每种螯合系统都包括1个外膜受体,1个胞周质结合蛋白及1个胞内ABC转运蛋白[16]。通过预测得知,巴氏杆菌的分泌蛋白中有较大比例参与了铁的代谢,而其他代谢相关酶类等对于细菌的增殖、分化及毒力都有十分重要的意义。分泌蛋白是在细胞内合成后运送到细胞外发生作用的蛋白,它是细胞与外界环境发生相互作用的物质基础。但是分泌蛋白在细胞代谢中具有的重要意义,需要更进一步的试验证实,以便于更清晰的了解其代谢机制和致病原理,筛选出免疫原性更好的蛋白。
[1] Wilson B A,Ho M.Pasteurella multocida:from zoonosis to cellular microbiology[J].Clin Microbiol Rev,2013,26(3):631-655.
[2] Wilkie I W,Harper M,Boyce J D,et al.Pasteurella multocida:diseases and pathogenesis//Pasteurella multocida[M].Springer Berlin Heidelberg,2012:1-22.
[3] Carter G R.Studies on Pasteurella multocida.I.A hemagglutination test for the identification of serological types[J].Am J Vet Res,1955,16(60):481-484.
[4] Harper M,Boyce J D,Adler B.Pasteurella multocida pathogenesis:125years after Pasteur[J].FEMS Microbiol Lett,2006,265(1):1-10.
[5] 赵莉莉,余利岩.以SecA 蛋白为“动力泵”的细菌Sec蛋白转运途径[J].微生物学通报,2008,35(7):1119-1123.
[6] 何冰芳,米 兰,陈文华.大肠杆菌蛋白质分泌机理及其重组蛋白分泌表达新进展[J].食品与生物技术学报,2012,31(6):561-569.
[7] May B J,Zhang Q,Li L L,et al.Complete genomic sequence of Pasteurella multocida,Pm70[J].Proc Nat Acad Sci,2001,98(6):3460-3465.
[8] Liu W,Yang M,Xu Z,et al.Complete genome sequence of Pasteurella multocida HN06,a toxigenic strain of serogroup D[J].J Bacteriol,2012,194(12):3292-3293.
[9] Bagos P G,Nikolaou E P,Liakopoulos T D,et al.Combined prediction of Tat and Sec signal peptides with hidden Markov models[J].Bioinformatics,2010,26(22):2811-2817.
[10] Petersen T N,Brunak S,von Heijne G,et al.SignalP 4.0:discriminating signal peptides from transmembrane regions[J].Nature Meth,2011,8(10):785-786.
[11] 于海瑛.膜蛋白跨膜螺旋结构预测研究[D].江苏南京:南京理工大学,2012.
[12] Ivankov D N,Payne S H,Galperin M Y,et al.How many signal peptides are there in bacteria[J].Environ Microbiol,2013,15(4):983-990.
[13] 杨 羽,吴清民.羊布鲁氏菌16M 基因组分泌蛋白的生物信息学分析[J].畜牧兽医学报,2009,40(7):1059-1062.
[14] 张秀红,王 颖,段江燕.长双歧杆菌亚种JCM1217基因组分泌蛋白的分析[J].中国食品学报,2012,11:160-169.
[15] Chan C S,Chang L,Winstone T M L,et al.Comparing system-specific chaperone interactions with their Tat dependent redox enzyme substrates[J].FEBS lett,2010,584(22):4553-4558.
[16] Krewulak K D,Vogel H J.Structural biology of bacterial iron uptake[J].Biochim Biophys Acta(BBA)-Biomembr,2008,1778(9):1781-1804.
