青天葵甲羟戊酸激酶基因的编码蛋白预测和表达分析
2015-05-30黄琼林何瑞詹若挺
黄琼林 何瑞 詹若挺

摘 要 甲羟戊酸途径(mevalonic acid, MVA)是植物三萜类化合物生物合成的主要途径,而甲羟戊酸激酶(mevalonate kinase, MVK)是MVA途径中的关键限速酶之一。根据前期获得的青天葵[Nervilia fordii(Hance)Schltr.]转录组数据,采用生物信息学方法对青天葵MVK基因编码蛋白的理化特性、功能结构域、二级结构、信号肽、跨膜结构域等进行预测,并采用RPKM法分析该基因在青天葵球茎和叶片的表达量。结果表明,NfMVK基因编码含有383个氨基酸的稳定型亲水性、酸性蛋白质,与其他植物MVK蛋白同源性可达70%。NfMVK蛋白分子量为41.29 ku,含有甲羟戊酸激酶功能域,归属于GHMP_Kinase超级家族。该蛋白没有任何信号肽、转运肽和跨膜结构域,二级结构为混合型结构的蛋白质。NfMVK基因在青天葵中的表达趋势为:球茎>叶片。该结果可为后续利用MVK基因调控青天葵三萜类活性成分的合成提供理论依据。
关键词 青天葵;甲羟戊酸激酶;生物信息学;表达量
中图分类号 R932 文献标识码 A
Abstract Mevalonic acid(MVA)pathway is the dominant approach to fulfill the triterpenoids synthesis in plants, and mevalonate kinase(MVK)is one of key rate-limiting enzymes in MVA pathway. In this paper, one MVK gene was identified from the transcriptome data of Nervilia fordii(Hance)Schltr. and named as NfMVK using bioinformatics. NfMVK encoded 383 amino acids and the coding protein was a stable hydrophilic and acid protein with a molecular weight at 41.29 ku. NfMVK protein, belonged to GHMP_Kinase superfamily, contained a mevalonate kinase functional domain and none signal peptide, and transmembrane region. Secondary structure prediction revealed NfMVK was a hybrid protein. The expression level of NfMVK gene in corm was higher than in leaf. The study would provide a firm foundation for regulating the synthesis of effective triterpenoids by utilization of MVK gene in N. fordii.
Key words Nervilia fordii(Hance)Schltr.; Mevalonate kinase; Bioinformatics; Expression
青天葵是岭南地区名贵中药,为兰科植物毛唇芋兰[Nervilia fordii(Hance) Schltr.]的干燥全草或叶片,具有清热、润肺、散瘀、解毒等显著功效[1-2]。由于自身萌发的苛刻条件和人类的过度开采,青天葵资源已经面临濒危。植物化学研究结果表明,三萜类化合物是青天葵发挥抗肿瘤、抗炎等功效的物质基础之一[3-6]。因此,在青天葵药效成分比较清楚的前提下,利用现代植物生物工程提高三萜类化合物的含量,是提高青天葵药效,缓解青天葵资源紧缺的新思路。
甲羟戊酸(mevalonic acid, MVA)途径是三萜类成分生物合成的主要途径[7],以乙酰辅酶A为原料,经过MVA这一关键前体和一系列酶促反应,生成重要中间体异戊烯基焦磷酸酯(isopentenyl diphosphate,IPP)及其双键异构体二甲丙烯二磷酸(dimethyallyl diphosphate, DMAPP)。IPP先与DMAPP结合生成牻牛儿基焦磷酸(geranyl diphosphate, GPP),再与GPP以头尾方式连接合成法尼基焦磷酸(farnesyl diphosphate, FPP),最后FPP以尾尾方式连接形成三萜类化合物[8]。甲羟戊酸激酶(mevalonate kinase, MVK)是MVA途径酶促反应中第一个ATP依赖酶,能将三磷腺苷上的磷酸基团转移到MVA的5位羟基上而产生甲羟戊酸-5-磷酸(mevalonate-5-phosphate, MVAP),这一过程还伴随着ADP的释放并最终生成IPP[9]。因此,MVK被认为是萜类生物合成MVA途径的关键限速酶之一。MVK基因参与MVA途径合成多种萜类及其衍生物,对植物萜类成分积累、生长发育具有重要的调控作用,目前已从杜仲[10]、三七[11]等药用植物的转录组数据中鉴定获得。本研究在前期青天葵转录组测序结果的基础上,对青天葵MVK基因进行生物信息学和表达量分析,为探讨MVK基因调控青天葵萜类成分的生物合成机制提供理论依据。
1 材料与方法
1.1 材料
前期研究[12]中,本课题组以生长旺盛期的成熟青天葵叶片和球茎为材料,利用Illumina RNA-seq高通量测序平台进行全转录组测序,通过序列组装、比对和注释共获得142 220条unigene。本研究以“mevalonate kinase”为搜索主题词,以E<10为标准,在获得注释的青天葵转录组基因数据库中检索编码青天葵甲羟戊酸激酶的Unigene序列。并在NCBI的Genbank数据库下载海枣、丹参等10种植物MVK基因的编码区序列。
1.2 方法
1.2.1 生物信息学分析 采用OMIGA软件将MVK基因编码区翻译成氨基酸序列,利用在线软件Protparam(http://web.expasy.org/protparam/)分析氨基酸序列,计算基因编码蛋白的基本理化参数,并运用Protscale在线工具(http://web.expasy.org/protscale/)分析蛋白的亲水性/疏水性特征;分别使用ClustalX1.86和MEGA 4.0软件完成氨基酸序列的多重比对和系统进化树的构建;采用NCBI的Conserved Domains Database(http://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)预测青天葵MVK蛋白具有的保守功能域;通过SOPMA在线工具(http://npsa-pbil.ibcp.fr/cgi-bin/npsa_automat.pl? page=npsa_sopma.html)分析青天葵MVK蛋白的二级结构分析;分别采用TargetP1.1(http://www.cbs.dtu.dk/services/TargetP/)和TMHMM (http://www.cbs.dtu.dk/ services/ TMHMM-2.0/)完成青天葵MVK蛋白的转运肽、信号肽和跨膜结构域的预测。
1.2.2 青天葵MVK基因的表达量分析 采用RPKM法(Reads Per Kb per Million reads)计算青天葵MVK基因在叶片和球茎的表达量,其计算公式为:
RPKM(MVK)=
设RPKM(MVK)为编码青天葵MVK基因的Unigene表达量,则C为唯一比对到编码青天葵MVK基因的Unigene读数,N为唯一比对到所有Unigene的总读数,L为编码青天葵MVK基因的Unigene的碱基数。
2 结果与分析
2.1 青天葵MVK基因检索结果
在青天葵转录组基因数据库里检索到注释为MVK基因的Unigene 16258,其长度为1 158 bp,包含MVK基因编码区全长。因此,选择Unigene 16258序列用于本研究分析,并命名为NfMVK。
2.2 MVK蛋白的基本理化特征分析
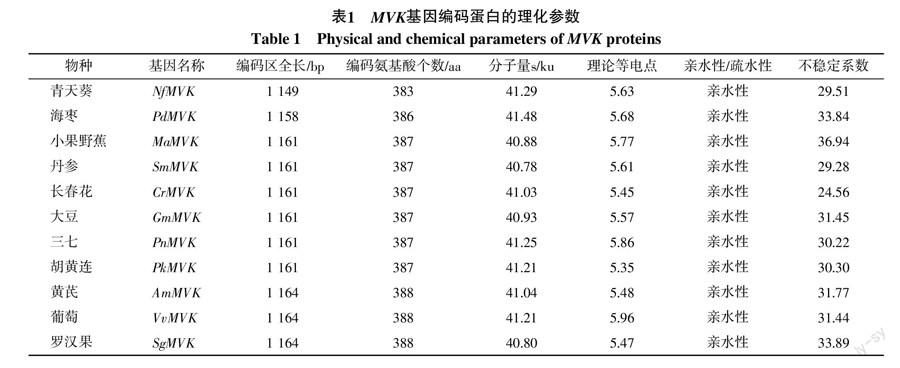
由表1可知,海枣等10种植物的MVK蛋白均含有380多个氨基酸,分子量在40~42 ku之间,说明不同植物来源的MVK蛋白在氨基酸个数和分子量上并无明显的差异;理论等电点均小于7,表明这些MVK蛋白均属于酸性蛋白质;不同植物MVK蛋白多肽链整体均显示为亲水性蛋白;不同来源的MVK蛋白的不稳定系数均小于40,属于稳定类蛋白质。NfMVK编码蛋白包含383个氨基酸,分子量为41.29 ku,也为稳定类亲水性、酸性蛋白质,可见其与其他MVK蛋白具有相似的理化特征。
2.3 MVK蛋白序列多重比对和进化分析
利用ClustalX 1.83软件对NfMVK基因编码蛋白与其余10种MVK蛋白进行多重序列比对,结果见图1。NfMVK编码蛋白与其他MVK蛋白序列存在着多处保守区域,其中与海枣MVK蛋白的相似度最高,为70%,与黄芪MVK蛋白相似较低,也可达63%,说明NfMVK编码蛋白与其他MVK蛋白有着较好的同源性,初步认为NfMVK为编码青天葵MVK蛋白的基因。
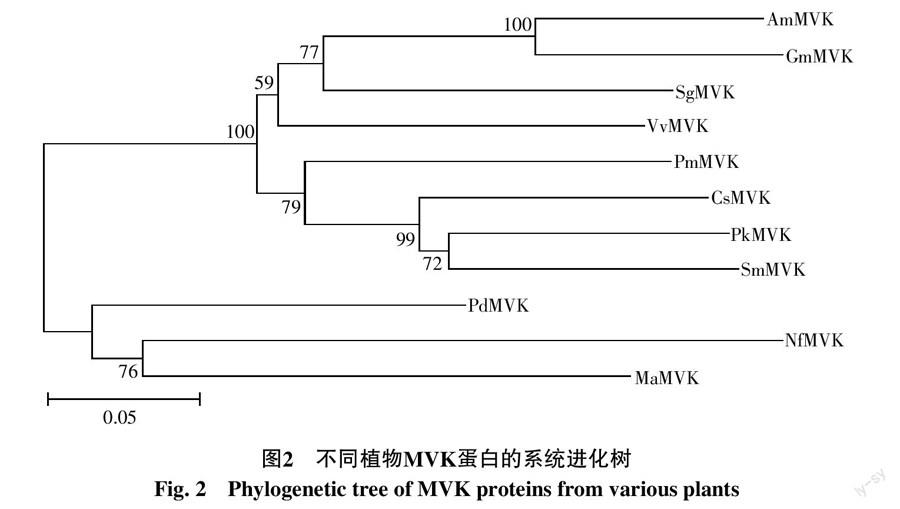
从系统进化树(图2)也可看出,NfMVK编码蛋白与同为单子叶植物来源的海枣、小果野蕉MVK蛋白首先聚成一支,而双子叶植物来源的MVK蛋白则聚焦成另外一个分支,这与传统植物进化结果相一致。
2.4 NfMVK编码蛋白的保守功能域预测
分析蛋白质序列中含有的功能域,能够推测蛋白质的功能。本研究采用CDD在线工具预测NfMVK编码蛋白的保守功能域,结果显示(图3),NfMVK编码蛋白包含GHMP_Kinase N端和C端功能域,分别位于130~211和293~365个氨基酸之间,说明其归属于包括高丝氨酸激酶、半乳糖激酶和甲羟戊酸激酶的GHMP_Kinase超级家族。除此之外,NfMVK编码蛋白还含有位于7~344个氨基酸之间的Mevalonate Kinase多域功能区,表明其具有甲羟戊酸激酶的功能,因此,NfMVK确定为青天葵甲羟戊酸激酶基因,其编码蛋白即青天葵甲羟戊酸激酶。将NfMVK核苷酸序列提交至Genbank,获得的登记号为KP280082。
2.5 青天葵MVK蛋白的信号肽、转运肽和跨膜结构分析
信号肽、转运肽位于蛋白的N端,前者指导分泌性蛋白到内质膜上合成,后者引蛋白质在细胞质基质游离核糖体上合成的前体蛋白转运至叶绿体基质中发挥作用。信号肽和转运肽的分析有利于对蛋白质的细胞定位等特性的掌握[13]。采用TargetP1.1在线工具分析结果显示,NfMVK蛋白均不含有信号肽和转运肽。
跨膜结构是膜内蛋白和膜脂层相结合、在细胞膜上发挥“锚定”作用的主要部位,可见预测蛋白质的跨膜结构域有助于对蛋白质的结构和定位等信息的理解。利用TMHMM软件预测NfMVK蛋白序列的跨膜结构域,结果如图4所示,NfMVK蛋白不含跨膜结构域,整条多肽链均处于细胞膜外。因此,NfMVK是一种在细胞质基质中合成并直接发挥功能的蛋白质。
2.6 青天葵MVK蛋白的二级结构预测
如图5所示,NfMVK蛋白二级结构由α-螺旋、不规则盘绕、β-转角和延伸链等元件组成,各元件所占比例分别为43.86%、34.20%、5.48%和16.45%。从蛋白的整体结构来看,α-螺旋和不规则盘绕是NfMVK最大量的结构元件,而延伸链和β-转角则相对分散于整条多肽链中。因此,NfMVK为混合型结构的蛋白质。
2.7 NfMVK基因的表达量分析
在对转录组高通量测序数据的处理中,RPKM法对测序长度和基因长度均作了归一化,使得不同长度的基因在不同测序深度下得到的基因表达水平估计值具有可比性,是目前最常用的基因表达量计算方法[14]。结果见图6所示,NfMVK在球茎中的RPKM表达量均高于在叶片中的RPKM表达量,提示青天葵甲羟戊酸激酶基因的表达趋势为:球茎>叶片。
3 讨论与结论
转录组分析已被公认为发现功能基因的有力手段。转录组以高通量模式挖掘功能基因,有助于整体、快速地解读药用植物的基因组信息,在分子水平为药用植物的功能基因的发现、活性成分的生物合成及其调控机制的阐明提供重要的科学依据[15]。因此比起传统的基因研究模式,经过简并引物PCR、RACE技术等繁琐步骤获取药用植物的功能基因,本研究通过转录组分析挖掘青天葵MVK基因具有明显的优势。
本研究应用生物信息学方法鉴定并分析了青天葵MVK的核苷酸和氨基酸序列,推测其为编码383个氨基酸,以α-螺旋和不规则盘绕为主要结构元件的酸性、亲水性稳定蛋白质,并与单子叶植物来源的MVK蛋白有着较高的同源性。青天葵MVK归属GHMP激酶超级家族中,含有甲羟戊酸激酶等3个保守功能域,表明其具有甲羟戊酸激酶的功能,如激酶活性、参与萜类代谢以及ATP结合细胞质定位等。TMHMM跨膜结构预测发现青天葵MVK蛋白不含跨膜结构域,说明其为细胞质酶,定位于细胞质中发挥催化作用。这些结论可为青天葵甲羟戊酸激酶的功能鉴定、表达载体构建、融合蛋白纯化、基因工程调控等实验研究提供理论指导。
甲羟戊酸途径在药用植物中可生成三萜皂苷合成所需的IPP等前体物质,因此MVK基因的表达情况可能会影响药用植物中三萜类物质的含量。研究表明,MVK基因主要分布在根等地下器官[11,16-17]。本研究也发现,NfMVK在地下组织球茎中的表达高于地上组织叶片,与在其他植物中的表达趋势相符合,推测青天葵三萜类成分主要在球茎中合成。
本研究利用生物信息学全面分析了NfMVK的分子功能,并初步明确其在青天葵中的表达趋势,可为后续利用融合蛋白表达和植物侵染表达等研究验证其生物学功能,为进一步应用NfMVK调控青天葵萜类成分合成奠定基础。
参考文献
[1] 陈蔚文. 岭南本草(二)[M]. 广州: 广东科技出版社, 2010: 326-351.
[2] 梅全喜. 青天葵的化学成分药理作用与临床应用研究进展[J]. 中华中医药学刊, 2008, 26(10): 2 239-2 241.
[3] 邱 莉, 徐灵源, 缪建华, 等. 青天葵植物化学成分和药理活性研究进展[J]. 时珍国医国药, 2011, 22(9): 2 258-2 260.
[4] 梁英娇, 楚桐丽, 丁 平. 赤芝中三萜类成分及其药理作用研究进展[J]. 2007, 18(2): 168-172.
[5] 金银萍, 焉 石, 刘俊霞, 等. 五味子科植物中环阿屯烷型三萜类成分及其药理作用研究进展[J]. 中草药, 2014, 45(4): 582-589.
[6] 金银萍, 焉 石, 刘俊霞, 等. 五味子科植物中羊毛脂烷型三萜类成分及其药理作用研究进展[J]. 中草药, 2014, 45(1): 137-143.
[7] 杨 鹤, 郜玉钢, 王璠瑛, 等. 人参皂苷等萜类化合物生物合成途径及HMGR的研究进展[J]. 中国生物工程杂志, 2008, 28(10): 130-135.
[8] Mahmoud S S, Croteau R B. Metabolic engineering of essential oil yield and composition in mint by altering expression of deoxyxylulose phosphate reductoisomerase and menthofuran synthase[J]. Proc Natl Acad Sci USA, 2001, 98: 8 915-8 920.
[9] Lange B M,Croteau R. sopentenyl diphosphate biosynthesis via a mevalonate-independent pathway: isopentenyl monophosphate kinase catalyzes the terminal enzymatic step[J]. Proc Natl Acad Sci USA, 1999, 96(24): 13 714-13 719.
[10] 乌云塔娜, 王 淋, 叶生晶. 杜仲甲羟戊酸激酶(EuMK)基因鉴定及生物信息学分析[J]. 经济林研究, 2014, 32(1): 6-12.
[11] 郭 溆, 罗红梅, 陈士林. 三七甲羟戊酸激酶PnMVK1基因的克隆和生物信息学分析[J]. 药学学报, 2012, 47(8): 1 092-1 097.
[12] 黄琼林. 青天葵转录组特征研究[D]. 广州:广州中医药大学, 2013.
[13] 黄琼林, 蔡 春. 不同植物黄酮醇合成酶FLS的生物信息学分析[J]. 广东农业科学, 2014, 41(13): 140-143, 151.
[14] 王 曦, 汪小我, 王立坤, 等. 新一代高通量RNA测序数据的处理与分析[J]. 生物化学与生物物理进展, 2010, 37(8): 834-836.
[15] 吴 琼, 孙 超, 陈士林, 等. 转录组学在药用植物研究中的应用[J]. 世界科学技术-中医药现代化, 2010, 12(3): 457-462.
[16] Lluch M A, Masferrer A, Arró M, et al. Molecular cloning and expression analysis of the mevalonate kinase gene from Arabidopsis thaliana[J]. Plant Mol Biol, 2000, 42: 365-376.
[17] Tang Wei, New ton R J. Mevalonate kinase activity during different stages of plant regeneration from nodular callus cultures in white pine(Pinus strobus)[J]. Tree Physiol, 2006, 26(2): 195-200.
