夹竹桃与珊瑚树光合光响应及PSⅡ叶绿素荧光特性比较
2015-04-18刘圣恩林开敏石丽娜陈梦瑶周丽春张秀娟
刘圣恩, 林开敏, 石丽娜, 陈梦瑶, 周丽春, 张秀娟
(1.福建农林大学林学院,福建 福州 350002;2.长江大学园艺园林学院,湖北 荆州 434025)
夹竹桃与珊瑚树光合光响应及PSⅡ叶绿素荧光特性比较
刘圣恩1, 林开敏1, 石丽娜1, 陈梦瑶1, 周丽春1, 张秀娟2
(1.福建农林大学林学院,福建 福州 350002;2.长江大学园艺园林学院,湖北 荆州 434025)
采用LI-6400型便携式光合测定仪,对夹竹桃和珊瑚树进行光合光响应拟合和叶绿素荧光光响应测定,分析Pn与荧光参数之间的联系.结果表明:夹竹桃和珊瑚树的光合光响应曲线变化趋势基本一致,采用非直角双曲线模型拟合良好.在同等光合有效辐射下珊瑚树的净光合速率(Pn)大于夹竹桃,夹竹桃的光补偿点(LCP)和光饱和点(LSP)分别为82.67 μmol·m-2·s-1和824.35 μmol·m-2·s-1,珊瑚树的LCP和LSP分别为33.93 μmol·m-2·s-1和757.19 μmol·m-2·s-1,珊瑚树的耐阴能力大于夹竹桃,夹竹桃的光强适应范围稍大于珊瑚树.在相同光照条件下夹竹桃的实际光化学效率(ΦPSⅡ)大于珊瑚树,夹竹桃通过光化学猝灭(qP)转换光能的作用高于珊瑚树,而以非光化学猝灭(NPQ)耗散光能的作用小于珊瑚树.两种灌木ETR和PCR光响应曲线变化的“拐点”可能与光饱和点有关.在园林应用中夹竹桃适合全光环境,珊瑚树适合适度遮阴环境.
夹竹桃; 珊瑚树; 光响应曲线; 叶绿素荧光
光合作用是作物生长发育和产量形成的基础,植物光响应曲线反映了光合速率随光照强度的变化特性[1].净光合速率(Pn)和叶绿素荧光参数是评价植物耐荫性的重要生理指标,对揭示植物耐荫性及其机理有重要作用[2-3].植物光补偿点(light compensation point, LCP)和光饱和点(light saturation point, LSP)的高低直接反映了植物对弱光的利用能力,是植物耐荫性评价的重要指标[4].一般光补偿点低的植物能在弱光环境中生存,光补偿点低且光饱和点高的植物能适应多种光环境[5]. 叶绿素荧光动力学参数反映光合作用过程的重要信息,与一些“表观性”的气体交换指标相比,叶绿素荧光参数在探测逆境对光合作用影响方面具有独特的作用,能够反映光合系统“内在性”特点[6-8],被视为研究植物光合作用与环境关系的内在探针.
夹竹桃(Neriumindicum),夹竹桃科,常绿灌木或小乔木,见于公园、厂矿、行道绿化,或是作庭园观赏植物.目前国内关于夹竹桃的研究主要集中在其活性成分皂甙对钉螺等生物的化感效应上[9],鲜见其在园林应用环境下的生理生化特性.珊瑚树(Viburnumodoratissimum),忍冬科,常绿灌木或小乔木,尤其在南方园林配置中应用广泛.有关珊瑚树光合特征的研究较多,包括其与银杏、女贞、海桐、大叶黄杨等园林优质树种的光合特性探讨[10-12],珊瑚树与夹竹桃的光合特性比较还未见报道.在园林设计中,植物的景观效果通常被放在第一位,而忽视了植物与环境之间的关系,长此以往,部分群落生长不良,甚至消失[13].因此只有同时兼顾植物的景观效果和生理生态特性才能充分发挥其园林和生态功能.本研究通过对夹竹桃和珊瑚树光合生理特性的比较,探讨其对环境重要因子之一,光因子的响应,并结合2个树种的生态特性深入分析,探讨两者对光能利用及其对光照环境变化的适应能力,比较两者的耐阴性,并以此为契机为今后城市园林绿化建设中植物的选择和应用提供依据.
1 材料与方法
1.1 试验地概况与材料
研究区位于荆州市沙市区太岳路段,位于江汉平原腹地,111°150′-114°050′ E,29°260′-31°370′ N,属亚热带季风气候区,年日照时数1800-2000 h,年平均气温15.9-16.6 ℃,年降雨量1100-1300 mm.土壤以水稻土、潮土、黄棕壤为主体,土层深厚肥沃.本研究选择的是荆州太岳路东西走向,相同生长环境下,健康、长势良好的10年生夹竹桃和珊瑚树为研究材料,夹竹桃平均株高3 m,平均胸径15 cm;珊瑚树平均株高4 m,平均胸径20 cm.
1.2 方法
1.2.1 光合光响应测定 于2013年7月晴朗无云的天气情况下,于上午08:00-11:30,日照充足,空气相对湿度40%-50%,空气温度32 ℃,风速1 m·s-1.取每株灌木上部发育良好的叶片,使用LI-6400便携式光合仪(Licor, USA),采用LI-6400的红蓝光源,设定CO2浓度为360 μmol·s-1,叶温30 ℃,相对湿度45%-65%,光合有效辐射强度(photosynthetically active radiation, PAR)从高到低设定为2000、1800、1600、1400、1200、1000、800、600、400、200、100、50、0 μmol·m-2·s-1,每一树种选取10株,待系统稳定后,每株取4片叶,每片叶对应即时光强记录5个Pn(net photosynthetic rate)值,并拟合光补偿点(LCP)和光饱和点(LSP).
1.2.2 叶绿素荧光光响应测定 使用Licor-6400叶绿素荧光叶室测定,植物叶片暗适应一昼夜,测定叶片初始荧光(Fo)后,以脉冲0.8 s、6000 μmol·m-2·s-1的强闪光.光强设置与光合光响应测定相同,最小等待时间2 min,最大4 min,CO2<20 mg·L-1时自动匹配,待系统稳定后.每一树种选取10株,每株取4片叶,每片叶记录5个相应值.记录初始荧光(the minimal fluorescence,Fo) 、最大荧光(the maximal fluorescence,Fm)、最大光化学效率(the maximal photochemical activity of photosystemⅡ,Fv/Fm),需要测定叶片光化学猝灭(the photochemical quenching coefficient,qP)、非光化学猝灭(non-photochemical quenching coefficient, NPQ)、最大光化学效率(the maximal photochemical activity of photosystemⅡ,Fv/Fm)、潜在光化学效率(the potential photochemical activity of photosystemⅡ,Fv/Fo)等指标.
1.3 数据处理
研究表明非直角双曲线模型在相关树种的光合光响应上拟合良好[14,15],且关于夹竹桃和珊瑚拟合的决定系数均达到0.96以上[5].模型如下:
(1)
Pn=φ×PAR-Rday
(2)
模型中Pn为净光合速率(μmol·m-2·s-1),PAR为光合有效辐射(μmol·m-2·s-1),Pmax为一定CO2浓度下的最大净光合能力(μmol·m-2·s-1),φ为表观量子效率(Apparentquantumefficiency,AQE),k为光响应曲线曲角,Rday为暗呼吸速率(μmol·m-2·s-1).用公式(2)对PAR小于200μmol·m-2·s-1进行线性回归得出LCP.用SPSS进行光响应模型的拟合以及参数估计,用Excel处理图表.
2 结果与分析
2.1 夹竹桃和珊瑚树光合光响应曲线
由图1可知夹竹桃和珊瑚树Pn随PAR的变化均包括两个阶段,PAR较小时,随PAR增大,Pn显著提高,此阶段为“快速响应”期.当Pn增加到一定值后,随PAR增大,Pn增幅渐趋平缓,此阶段为“平稳期”.在“快速响应”期珊瑚树Pn随PAR增加趋势明显大于夹竹桃,且相同光合有效辐射下珊瑚树的净光合速率明显大于夹竹桃,表明相同光照条件下珊瑚树的光能利用率高于夹竹桃.且从图1可以看出,珊瑚树最大净光合速率(Pmax)明显大于夹竹桃,且拟合的结果与真实值相符合.
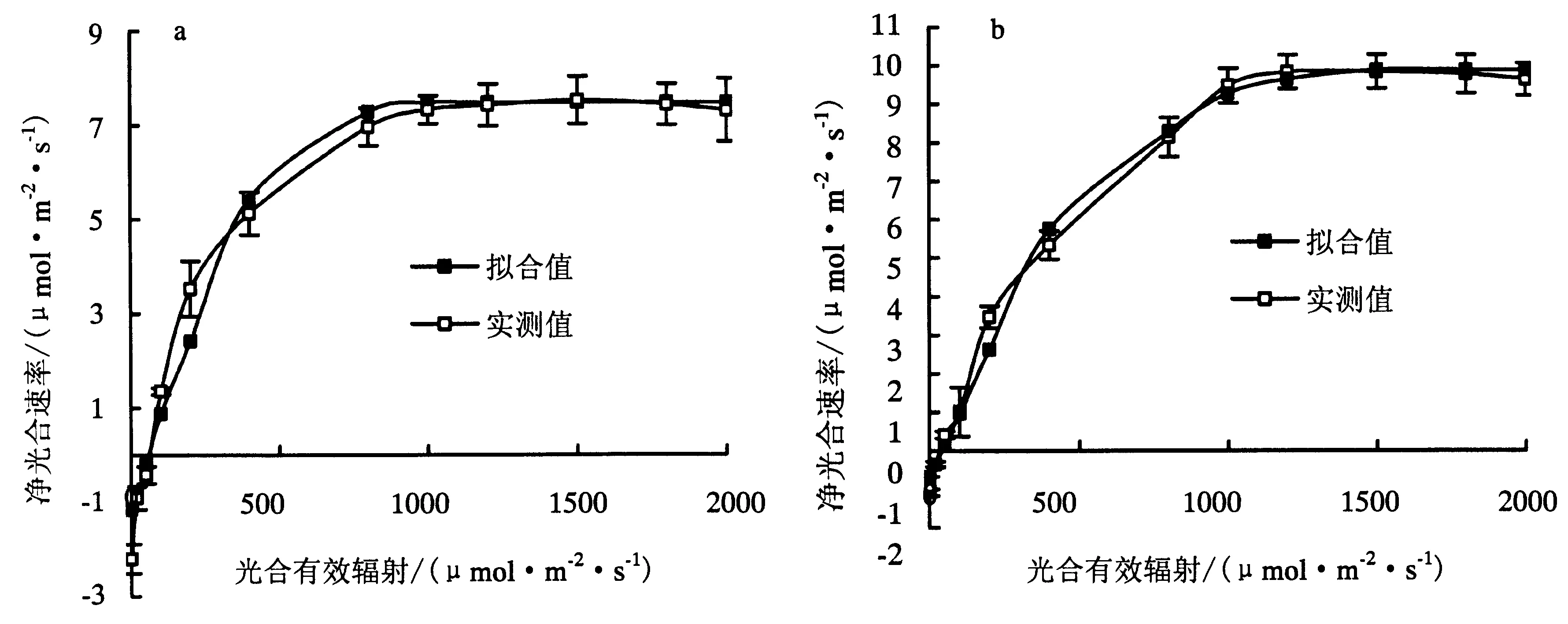
a:夹竹桃;b:珊瑚树.
2.2 夹竹桃和珊瑚树的光合光响应参数
在一定环境条件下,叶片的最大净光合速率(Pmax)反映了植物叶片的最大光合能力.研究表明(表1),相同光照条件下,珊瑚树的Pmax是夹竹桃的1.7倍,珊瑚树的AQE是夹竹桃的1.2倍,两者之间差异显著(P<0.05).Rday反映的是植物在没有光照条件下的呼吸速率,与叶片的生理活性有关[16].表1中夹竹桃的暗呼吸速率大于珊瑚树表明夹竹桃的生理活性高于珊瑚树,也是在相同PAR下珊瑚树的Pn大于夹竹桃的原因之一.夹竹桃的LCP是珊瑚树的1.6倍,表明珊瑚树对弱光适应和利用能力强于夹竹桃,而夹竹桃的LSP是珊瑚树的1.06倍,表明在一定的光强范围内,夹竹桃的强光适应和利用能力高于珊瑚树.夹竹桃的拟合参数为0.921,珊瑚树的拟合参数为0.988,相比较而言,珊瑚树的拟合效果更好.
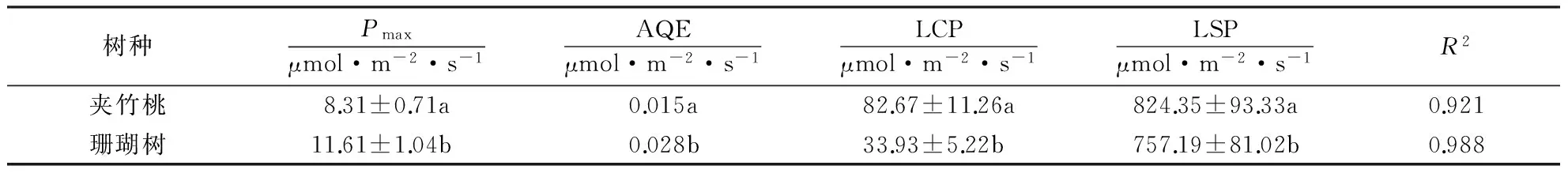
表1 夹竹桃和珊瑚树光合光响应拟合参数1)
1)同列带有不同的字母表示在0.05水平上差异性显著.
2.3 夹竹桃和珊瑚树叶绿素荧光光响应特征
2.3.1 夹竹桃和珊瑚树叶绿素荧光特性比较 如图2夹竹桃和珊瑚树的实际光化学效率(ΦPSⅡ)(图a)对PAR的响应曲线所示,两者的的变化趋势基本一致,均表现为PAR较低时时实际光化学效率较高,且随着PAR的增加ΦPSⅡ值逐渐减少,表明随着光照强度的增加PSⅡ功能反应中心开放程度降低.夹竹桃和珊瑚树有效光化学效率(Fv′/Fm′)(图b)光响应曲线变化趋势与ΦPSⅡ光响应曲线相似,都是随着PAR的增加Fv′/Fm′逐渐下降,珊瑚树的Fv′/Fm′光响应曲线变化斜率,即下降趋势在任一点都显著大于夹竹桃,表明夹竹桃反应中心活性较珊瑚树稳定且大于珊瑚树.夹竹桃和珊瑚树Fv′/Fm′光响应曲线拐点所对应的PAR值不同可能与它们的光饱和点不同有关.ΦPSⅡ是PSⅡ反应中心部分关闭情况下的实际PSⅡ光能捕获的效率,反映叶片用于光合电子传递的能量占所吸收光能的比例,即实际光能利用率,夹竹桃的光能利用率高于珊瑚树.
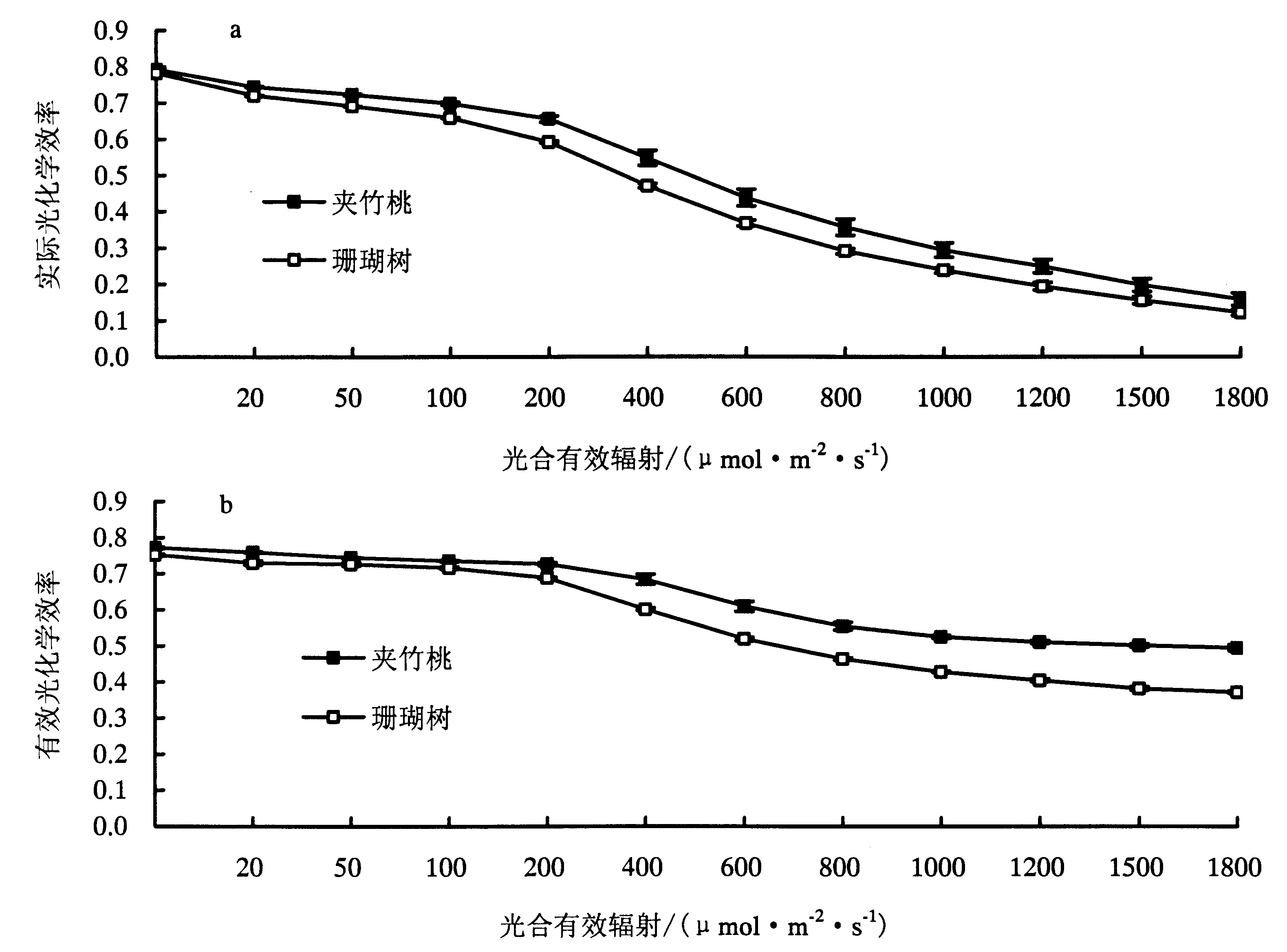
a:ΦPSⅡ光响应;b:Fv′/Fm′光响应.
2.3.2 夹竹桃和珊瑚树qP和NPQ光响应比较 叶绿素吸收的光能主要通过光化学反应、热、荧光3 种形式进行耗散,三者对能量竞争反映在不同形式荧光猝灭对能量的竞争[17].qP(图a)反映PSⅡ所捕获的光量子转化成化学能的效率,反映了植物的光合效率和对光能的利用情况,NPQ(图b)反映PSⅡ天线色素吸收的光能以热的形式耗散的那部分光能[18].如图3所示,夹竹桃和珊瑚树的光化学猝灭光响应曲线变化趋势基本一致,均变现为PAR较低时时qP较高,且随着PAR的增加qP几乎呈直线下降,且夹竹桃和珊瑚树的qP光响应曲线几乎重合.PAR在0-200 μmol·m-2·s-1之间夹竹桃的NPQ>珊瑚树,PAR>200 μmol·m-2·s-1,珊瑚树的NPQ大于夹竹桃,且随PAR增大,两者之间的差额在逐渐扩大.NPQ与qP光响应曲线随PAR的增加,变化趋势正好相反,说明NPQ和qP呈负相关,也表明了qP和NPQ对叶绿素吸收光量子激发能的能量转换存在着竞争关系[19].
2.3.3 夹竹桃和珊瑚树ETR和PCR光响应比较 光照条件下植物叶片叶绿素荧光参数对光照强度有强烈的依赖性[20].如图4所示,两种灌木的ETR(electron transport rate)(图a)和PCR(Photochemistry rate)(图b)光响应变化具有协同性,均表现“快速光响应”和“慢速光响应”两个阶段,且两种灌木所对应的同一荧光参数差异显著.在快速光响应阶段,随PAR的增加夹竹桃和珊瑚树的ETR和PCR几乎呈直线增加,且两种灌木在同一光强下的ETR或PCR的差额逐渐变大.在“慢速光响应”阶段,随PAR的增加夹竹桃和珊瑚树的ETR和PCR的变化开始变缓,且慢慢趋于稳定水平.从图4可以看出两种灌木ETR或是PCR光响应曲线的两个变化阶段明显存在一个“拐点”且两者的拐点均在400-800 μmol·m-2·s-1之间,而由表1可知夹竹桃的光饱和点为824.35 μmol·m-2·s-1,珊瑚树光饱点为757.19 μmol·m-2·s-1,在误差允许的范围内推测ETR和PCR光响应曲线的拐点与光饱和点有关.
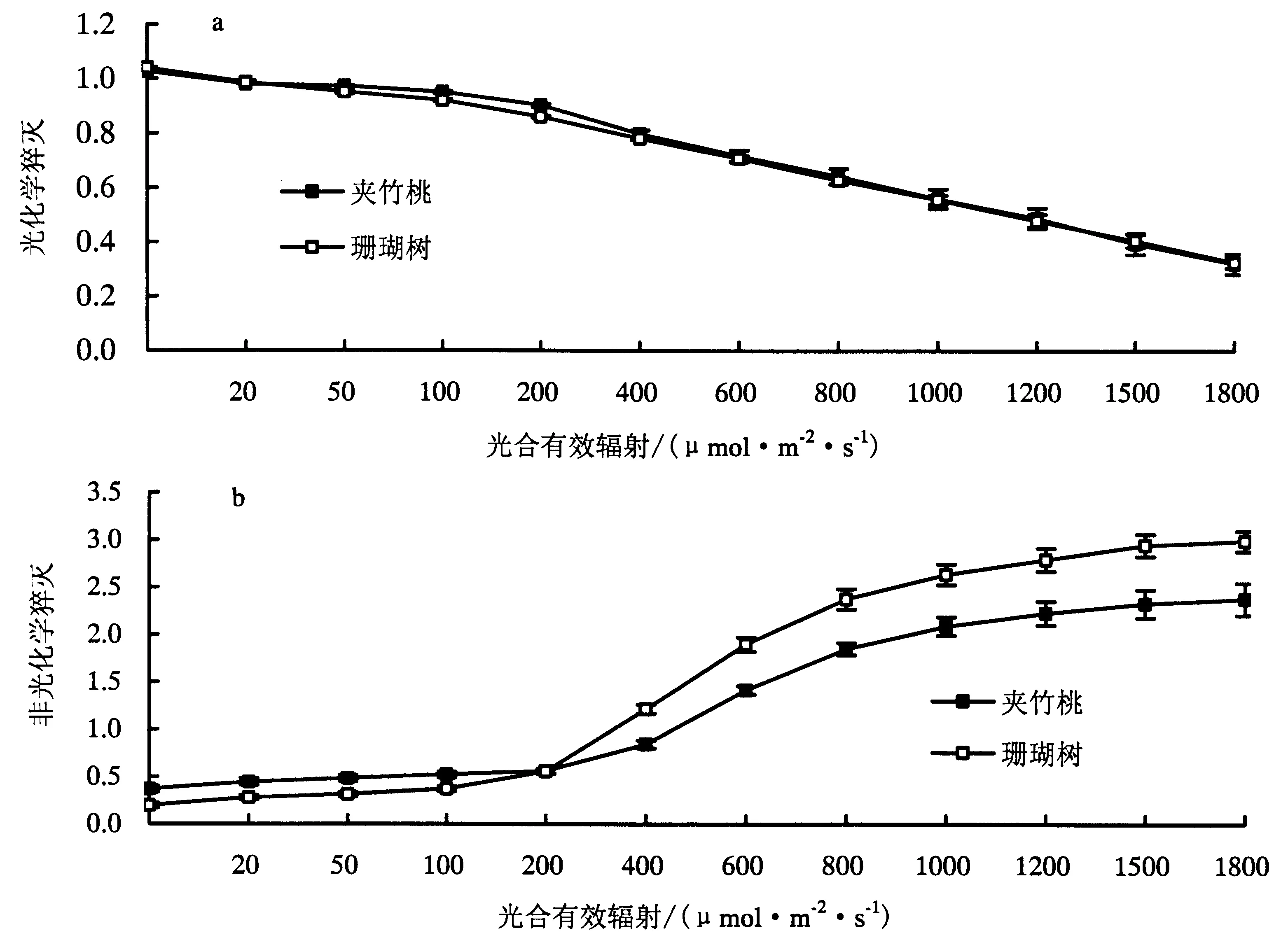
a:qP光响应;b:NPQ光响应.
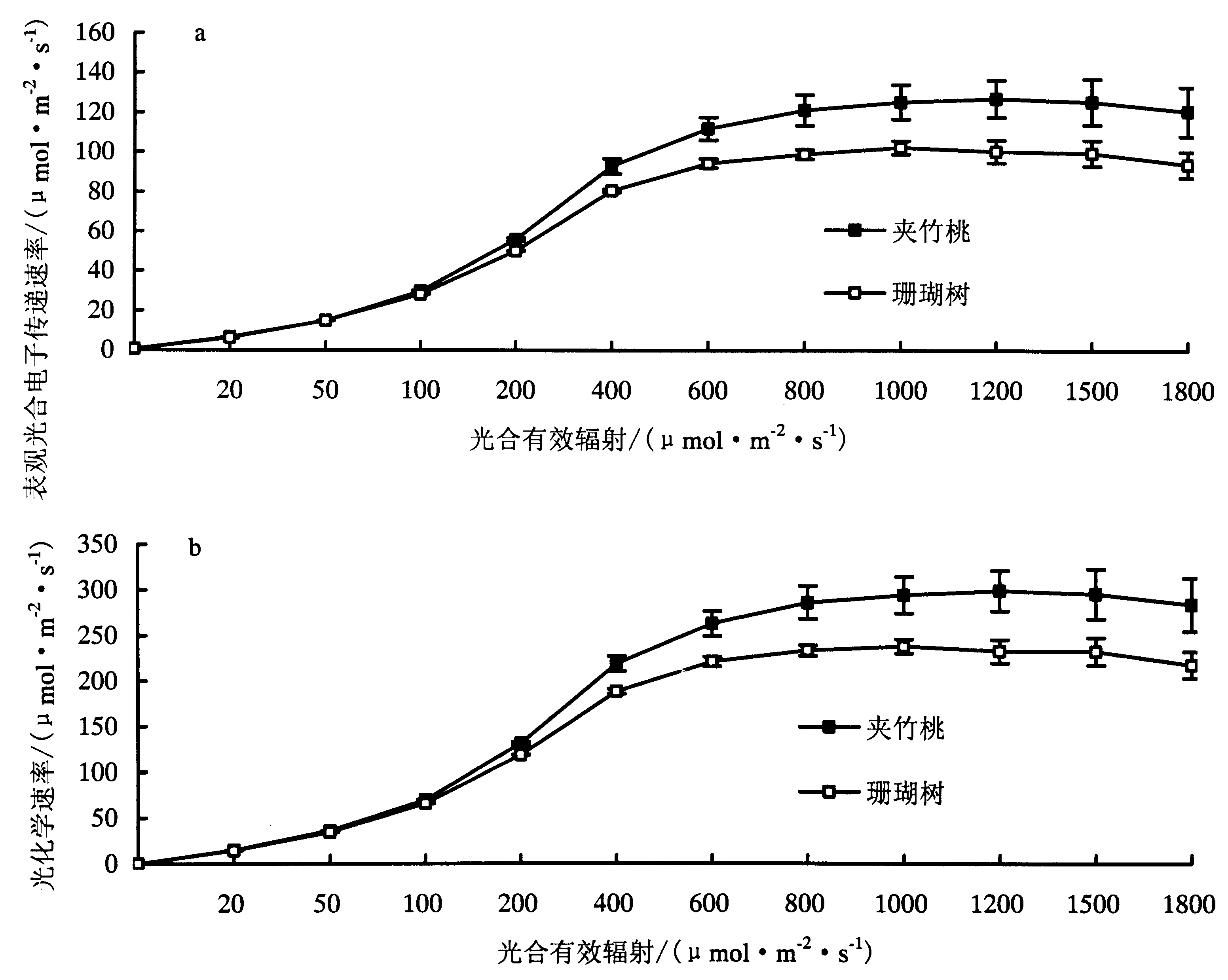
a:ETR光响应;b:PCR光响应.
3 讨论
描述净光合速率对光合有效辐射响应曲线的数学模型较多,包括直角双曲线模型、非直角双曲线模型、Prado-Moraes模型、直角双曲线修正模型等等,不同模型对不同植物拟合的效果不同.闫小红等[14]使用不同模型对4个不同水稻品种的光合光响应进行拟合,发现非直角双曲线模型拟合效果更好.陈卫英等[21]对高粱、苋、大麦、半夏进行研究发现指数修正模型优势更加明显.随着研究的深入,叶子飘等[22]提出了C3植物新模型,并利用新模型对华北平原冬小麦进行拟合,克服了无法拟合PAR达LSP以后Pn随光强增加而降低的特征和LSP拟合值比实测值偏大的缺点.因此针对不同植物光合光响应拟合时应根据植物光响应特征和模型特点选择合适的模型,这样拟合的结果和参数估计才更准确.本研究中采用非直角双曲线模型对夹竹桃和珊瑚树光合光响应曲线进行拟合,各方程的决定系数分别为0.921和0.988,并对0-200 μmol·m-2·s-1光照区间进行线性回归获得了多项光合参数,说明该模型适合夹竹桃和珊瑚树的光合光响应拟合,较好的反映了两种灌木叶片光合光响应过程.
LSP和LCP是植物的2个重要的光合生理指标,一般LCP和LSP均较低是典型的耐荫植物,LCP较低而LSP较高的植物对环境适应性很强,LCP较高而LSP较低的植物对光照的适应范围相对较窄,LCP和LSP点都较高的是典型阳性植物[23].通常阴生植物的LCP<20 μmol·m-2·s-1,阳生植物LCP为50-100 μmol·m-2·s-1[24].本研究中夹竹桃的LCP和LSP分别为82.67和824.35 μmol·m-2·s-1,珊瑚树的LCP和LSP分别为33.93和757.19 μmol·m-2·s-1,表明珊瑚树的耐荫性>夹竹桃,且夹竹桃的光强适应范围大于珊瑚树.本研究与陈军等[10]发现珊瑚树耐荫性也大于银杏有相似之处.因此两种灌木在园林配置中应充分考虑两者耐荫能力的差异,进行合理配置,夹竹桃适合全光照环境,珊瑚树适合半阴环境或进行适当遮荫.陈军等在实验中测得的珊瑚树LSP为1 125 μmol·m-2·s-1与本研究的结果差异较大主要与研究对象所处环境、叶片成熟状况、生态因子胁迫程度不同有关.此外苏吉虎等[25]测得的珊瑚树阳生叶LSP为800-900 μmol·m-2·s-1,阴生叶为300-400 μmol·m-2·s-1.可见阳生叶和阴生叶也是影响同一树种光合特性差异的原因.表观量子效率(AQE)反映了叶片在弱光情况下的光合能力,其值越大,植物利用弱光能力越强[26].珊瑚树AQE是夹竹桃的1.2倍,两者之间差异显著(P<0.05),表明珊瑚树对弱光的适应和利用能力强于夹竹桃.通常在适宜生长的条件下,植物的实测AQE在0.03-0.05[27].本研究中夹竹桃和珊瑚树AQE分别为0.015和0.028,夹竹桃可能受外界环境因子轻度胁迫.这与研究材料所处的生态环境有关,实验中的夹竹桃与珊瑚树均处于车流高峰路段,汽车噪音,尾气,粉尘等的污染对植物光合生理特性造成一定的影响.夹竹桃终年常绿,夏季花期繁盛,具有很好的药用价值,由于其毒性较强特点,在园林绿化上,应慎重选择[28].珊瑚树具有春花秋果,四季常青的特性,在园林应用中,夹竹桃对环境,特别是土壤要求不高,具有耐修剪、抗污染能力强的特点,是绿篱和街道绿化的上佳材料[29].再加上其稍喜光,耐阴的特点,可作为南方城市高架桥下园林绿化的配置树种.相关研究表明珊瑚树在高架桥下应用效果良好[5],不过由于高架桥特殊生境条件的限制[30],珊瑚树具体的立地选择及造景形式应深入研究后再做决定.
本研究中,夹竹桃(Fv/Fm=0.838)>珊瑚树(Fv/Fm=0.819),夹竹桃(Fv/Fo=5.16)>珊瑚树(Fv/Fo=4.51),但是在光合——光响应过程中夹竹桃Pn却低于珊瑚树.这是由于光合碳同化能力除了受光化学反应形成的NADPH与ATP的影响,碳同化酶活性也起着关键的作用[31].夹竹桃和珊瑚树叶绿素荧光参数,包括qP、NPQ等的光响应曲线明显存在某一拐点,把叶绿素荧光光响应进程分为“快速光响应”和“缓慢光响应”两个阶段.何炎红等[32]研究表明,qP、NPQ光响应曲线拐点可作为光抑制初始点,利用相关模型进行拟合,该点对应的光强度可作为评价不同树木抵御光胁迫伤害能力和对强光适应能力的指标,本研究有待在此方面进行深入研究.
[1] 蒋高明,何维明.一种在野外自然光照条件下快速测定光合作用——光响应曲线的新方法[J].植物学通报,1999,16(6):712-718.
[2] 王建华,任士福,史宝胜,等.遮荫对连翘光合特性和叶绿素荧光参数的影响[J].生态学报,2011,31(7):1811-1817.
[3] 王瑞,丁爱萍,杜林峰,等.遮阴对12种阴生园林植物光合特性的影响[J]. 华中农业大学学报,2010,29(3):369-374.
[4] 王建华,任士福,史宝胜,等.遮荫对连翘光合特性和叶绿素荧光参数的影响[J].生态学报2011,31(7):1811-1817.
[5] 殷利华.基于光环境的城市高架桥下桥阴绿地景观研究——以武汉城区高架桥为例[D].武汉:华中科技大学,2004.
[6] 钱永强,周晓星,韩蕾,等.Cd2+胁迫对银芽柳PSⅡ叶绿素荧光光响应曲线的影响[J].生态学报,2011,31(20):6134-6142.
[7] 李鹏民,高辉远,RETO J S.快速叶绿素荧光诱导动力学分析在光合作用研究中的应用[J].植物生理与分子生物学学报,2005,31(6):559-566.
[8] 白志英,李存东,赵金锋,等.干旱胁迫对小麦代换系叶绿素荧光参数的影响及染色体效应初步分析[J].中国农业科学,2011,44(1):47-57.
[9] 王万贤,杨毅,王宏,等.夹竹桃强心总甙灭螺活性与机理[J].生态学报,2006,26(3):954-959.
[10] 陈军,张军,雷忻,等.银杏与珊瑚树光合及蒸腾特性研究[J].延安大学学报:自然科学版,2004,23(1):75-78.
[11] 石辉,王会霞,李秧秧,等.女贞和珊瑚树叶片表面特征的AFM观察[J].生态学报,2011,31(5):1471-1477.
[12] 陈洪国.四种常绿植物蒸腾速率,净光合速率的日变化及对环境的影响[J].福建林业科技,2006,33(1):76-79.
[13] 朱丽清.从生态适应性探讨城市园林植物的配置[J].安徽农业科学,2010,38(27):15221-15223.
[14] 闫小红,尹建华,段世华,等.四种水稻品种的光合光响应曲线及其模型拟合[J].生态学杂志,2013,32(3):604-610.
[15] 韩刚,赵忠.不同土壤水分下4种沙生灌木的光合光响应特性[J].生态学报,2010,30(15):4019-4026.
[16] 张淑勇,周泽福,夏江宝,等.不同土壤水分条件下小叶扶芳藤叶片光合作用对光的响应[J].西北植物学报,2007,27(12):2514-2521.
[17] HE Y H, GUO L S, TIAN Y L. Photosynthetic rates and chlorophyll fluorescence ofNirariatangutorumat different leaf water potentials[J]. Acta Bot. Boreal Occident.Sin, 2005,25(11):2226-2233.
[18] ZHANG S R. A discussion on chlorophyll fluorescence kinetics parameters and their significance[J]. Chinese Bulletin of Botany, 1999,16(4):444-448.
[19] 王颖,魏国印,张志强,等.7种园林树种光合参数及水分利用效率的研究[J].河北农业大学学报,2006,29(6):44-48.
[20] 尤鑫,龚吉蕊,葛之葳,等.两种杂交杨叶绿素荧光特性及光能利用[J].植物生态学报,2009,33(6):1148-1155.
[21] 陈卫英,陈真勇,罗辅燕,等.光响应曲线的指数改进模型与常用模型比较[J].植物生态学报,2012,36(12):1277-1285.
[22] 叶子飘,于强.光合作用光响应模型的比较[J].植物生态学报,2008,32(6):1356-1361.
[23] 宋宁,郭平毅,原向阳.膜荚黄芪与蒙古黄芪幼苗的光能利用及其耐光性研究[J].西北植物学报,2011,31(11):2283-2289.
[24] 王珊珊,陈曦,王权,等.新疆古尔班通古特沙漠南缘多枝柽柳光合作用及水分利用的生态适应性[J].生态学报,2011,31(11):3082-3089.
[25] 苏吉虎,沈允钢.珊瑚树阳生和阴生叶片光合特性和状态转换的比较[J].植物生理与分子生物学学报,2003,29(5):443-448.
[26] 蒋高明,植物生理生态学[M]. 北京:高等教育出版社,2004:59-65.
[27] 刘建峰,杨文娟,史青胜.崖柏与侧柏光和特性和叶绿素荧光参数的比较研究[J].西北植物学报,2011,31(10):2071-2077.
[28] 郝福玲,方访,凌铁军,等.夹竹桃叶化学成分的研究[J].安徽农业大学学报,2013,40(5):795-801.
[29] 贾德华,王万喜.珊瑚树在园林绿化中的应用[J].安徽农业科学2007,35(5):1351-1353.
[30] 王雪莹,辛雅芬,宋坤,等.城市高架桥荫光照特性与绿化的合理布局[J].生态学杂志,2006(8):938-943.
[31] 伍维模,董合林,危常洲,等.南疆陆地棉与海岛棉光合——光响应及叶绿素荧光特性分析[J].西北农业学报,2006,15(4):141-146.
[32] 何炎红,郭连生,田有亮.七种阔叶树种不同光照强度下叶绿素荧光猝灭特征[J].林业科学,2006,42(2):27-31.
(责任编辑:吴显达)
Comparison on photosynthesis light response and PSⅡ chlorophyll fluorescence characteristics betweenNeriumindicumandViburnumodoratissimum
LIU Sheng-en1, LIN Kai-min1, SHI Li-na1, CHEN Meng-yao1, ZHOU Li-chun1, ZHANG Xiu-juan2
(1.College of Forestry, Fujian Agriculture and Forestry University, Fuzhou, Fujian 350002, China;2.College of Horticulture and Garden, Yangtze University, Jingzhou, Hubei 434025, China)
LI-6400 portable photosynthesis machine was used to fit the photosynthesis light response curves and test the chlorophyll fluorescence light response inNeriumindicumandViburnumodoratissimum, and the relationship betweenPnand fluorescence parameters was analyzed. The results showed that the light-response curves ofN.IndicumandV.odoratissimumtended to be the same. Non-rectangular hyperbola model well fitted the light response curves ofN.IndicumandV.odoratissimum. At the same PAR, the net photosynthetic rate (Pn) ofV.odoratissimumwas higher thanN.indicum, the light compensation point (LCP) and light saturation point (LSP) ofN.Indicumwere 82.67 μmol·m-2·s-1and 824.35 μmol·m-2·s-1, and those ofV.Odoratissimumwere 33.93 μmol·m-2·s-1and 757.19 μmol·m-2·s-1. The shade tolerance capability ofV.odoratissimumwas higher than that ofN.indicum, but the light intensity adaptation range ofN.indicumwas slightly larger than that ofV.odoratissimum. The quantum efficiency of photosystemⅡ (ΦPSⅡ) and photochemical quenching transformation (qP) ofN.indicumwere significantly higher than those ofV.odoratissimum, and the non-photochemical quenching dissipative energy effect (NPQ) was smaller. ETR and PCR light response curves obviously displayed an inflection point, which divided chlorophyll fluorescence response process into “quick response” and “slow response”, and the point might expose potential relationship with LSP.N.indicumwas adaptable for all-optical environment andV.odoratissimumwas adaptable for properly-shaded place in the landscape application.
Neriumindicum;Viburnumodoratissimum; light-response curves; chlorophyll fluorescence
2014-10-08
2014-12-01
福建省自然科学基金项目(2013J01075);国家自然科学基金项目(30872020);福建农林大学部级创新平台资助.
刘圣恩(1989-),男,硕士研究生.研究方向:森林生态学.Email:824048128@qq.com.通讯作者张秀娟(1979-),女,博士,讲师.研究方向:园林植物应用与生态系统恢复.Email:zxj510@yahoo.com.cn.
Q945.79
A
1671-5470(2015)04-0384-07
10.13323/j.cnki.j.fafu(nat.sci.).2015.04.009
