不同海拔川滇高山栎叶片的解剖结构特征
2015-04-17姜永雷邓莉兰黄晓霞
姜永雷 +邓莉兰 +黄晓霞

摘要:采用生态解剖学方法,分析滇西北玉龙雪山自然保护区不同海拔(2 750、2 900、3 050、3 200、3 350、3 500 m)梯度内川滇高山栎叶片细胞解剖结构特征,探讨其对高山环境的生态适应机制。结果表明:随着海拔的升高,叶片总厚度、表皮厚度、栅栏组织厚度、海绵组织厚度和栅栏组织厚度/海绵组织厚度的比值(P/S值)等均呈增大趋势;而栅栏细胞的长径、短径等的变化呈降低趋势;叶片解剖结构在不同海拔高度的这种差异表明,不同海拔梯度上复杂的环境条件限制了植物的生长和分布;川滇高山栎通过叶片细胞结构组织发生的变化是在一定程度上适应环境的表现。
关键词:川滇高山栎;叶片解剖;海拔;形态结构;适应环境
中图分类号:Q944.56;S792.180.1 文献标志码: A文章编号:1002-1302(2015)01-0195-04
收稿日期:2014-06-06
基金简介:国家自然科学青年基金(编号:31100292);云南省省部级重点学科、省高校重点实验室及校实验室共享平台资助;西南林业大学博士启动项目(编号:11032)。
作者简介:姜永雷(1988—),男,河南周口人,硕士研究生,研究方向为园林植物。E-mail:10-29jyl@163.com。
通信作者:黄晓霞,博士,副教授,主要从事植物生理生态研究。E-mail:huangxx@swfu.edu.cn。环境因子对植物的分布有重要的影响,其中海拔对植物的生长发育、组织功能和叶片的解剖结构等影响较显著,是影响植物分布的重要生态因素之一[1-2]。海拔对植物分布的限制归结于高海拔的自然环境因子,随着海拔的上升,平均气温下降,大气压及CO2分压降低,光强增强,UV-B辐射增加等,这些因素对植物叶片的形态结构和生理特性都有重要的影响。此外,随着海拔的升高,土壤养分、含水量也都能极大地影响高山植物的生长,导致植物叶片的组织结构发生差异。有研究认为[3-4],植物叶片组织结构的差异是对环境变化的一种响应,叶片是植物进化过程中对环境变化比较敏感且可塑性较大的器官,与植物的光合作用和呼吸作用有着密切的关系,环境因子如水分、温度、光照等的变化常会导致叶的外部形态及厚度、表皮细胞及附属物,叶肉栅栏组织、海绵组织、胞间隙、厚角组织和叶脉等形态解剖结构差异[5],这种差异与植物体的生态生理功能以及生态系统密切相关[6-8]。不同海拔上植物叶片的组织结构差异都是植物对海拔梯度变化的一个长期的适应过程。因此,通过研究叶片解剖结构特征,分析其组织结构在海拔梯度上的差异性,这对揭示植物的适生环境有重要意义,而且本土植物对不同海拔梯度下环境变化的长期适应形成了稳定的结构[9],探讨这些结构特征与海拔之间的相互关系对认识植物适应生态环境变化有重要意义。
川滇高山栎(Quercus aquifolioides)为壳斗科(Fagaceae)栎属(Quercus)多年生常绿木本植物,属亚高山硬叶栎林,是我国特有的木本植物,广泛分布于川滇两省,垂直分布幅度极宽,生态适应幅度很大[10]。目前,对川滇高山栎的研究主要集中在光合特性[11]、遗传多样性[12]和生理特性[13]方面,有关海拔对川滇高山栎叶片解剖结构特征影响的研究少见报道。本研究以生长于滇西北玉龙雪山自然保护区内的川滇高山栎天然林为试验材料,通过在不同海拔生境下对川滇高山栎叶片作解剖结构特征比较分析,了解其形态结构特征与环境因子之间的关系,比较不同海拔梯度下川滇高山栎叶片形态结构对环境的响应,其研究结果为揭示亚高山植物对生态环境的适应机制及预测植物对未来气候变化的响应提供一定的理论依据。
1材料与方法
1.1研究地概况
研究区位于滇西北亚高山玉龙雪山自然保护区内,其西北临长江石鼓第一湾和金沙江虎跳峡,东麓是海拔约3 km的干海子盆地,南面是丽江盆地。玉龙山体南北长35 km、东西宽13 km,主山脊上有13座海拔5 km以上的高峰,主峰扇子陡海拔5 596 m,耸立于群峰之上,与南麓的雪嵩村(海拔为 2 700 m)的相对垂直高相差近3 km,而与西北部的金沙江虎跳峡相差约3 900 m[14]。巨大的海拔差异导致气候条件不同,从而形成了不同的森林群落类型,呈现出明显的山地垂直分布带谱。在海拔2 750~2 900 m均有灌木的川滇高山栎,成林集中在2 900~3 350 m。2 900 m为川滇高山栎和云冷杉林带的交错带,交错带以上形成东亚地区保存最完整的川滇高山栎林带,郁闭度在0. 4~0. 6之间,平均树高 7~12 m,平均胸径可达18~30 cm。海拔3 500 m是川滇高山栎林在我国滇西北部分布的最高限。由于风、雪等因素的影响,川滇高山栎以斑块状镶嵌于海拔高于3 500 m的高山苔原带,并逐渐向上扩展。随着海拔的升高,玉龙雪山气候变冷变湿,不适于很多树种生长,使得川滇高山栎林形成优势,乔木除云杉冷杉、云南松和川滇高山栎有分布外,很少有其他种类。
1.2研究材料
供试叶片为采自中国滇西北玉龙雪山自然保护区内不同海拔(2 750、2 900、3 050、3 200、3 350、3 500 m)梯度的川滇高山栎叶片。根据玉龙雪山自然保护区内川滇高山栎分布的特征,于2013年5月上旬在玉龙雪山(100°10′~100°20′E、27°10′~27°40′N)上采集川滇高山栎叶片,设置6个采样点2 750、2 900、3 050、3 200、3 350、3 500 m,每个采样点选取若干株(n≥5)长势相当、无病虫害、成熟的中树(树高5~12 m,胸径7~12 cm),分别采集其叶片,采样时间为09:30—11:30。取样时在树冠下部东、南、西、北4个方向采摘一年生枝上的成熟叶片,各个方位上取等量叶片,混匀。用 0.1 mol/L 磷酸缓冲液(pH值 6. 8)轻洗涤除去表面污物,再用FAA(50%乙醇 ∶38%甲醛 ∶99.5%冰醋酸=90 ∶5 ∶5)野外固定,立即固定并保存。样品统一编号,带回西南林业大学园林学院实验室进行叶片解剖结构参数的测定。
1.3研究方法
1.3.1切片制作和叶片解剖机构参数的测定切片的制作参考李芳兰的方法[15];在光学显微镜(Nikon ECLIPSE Ti-S,LEICA DM 2000)下观测叶片总厚度、叶表皮厚度、栅栏组织厚度、细胞层数、海绵组织厚度、栅栏组织细胞的长短径等7项指标,每项指标重复测5次,取其平均值。
1.3.2统计分析所有的数据分析都利用SPSS 11.5 统计分析软件进行一元方差分析(one-way ANOVA),分析海拔梯度上叶片解剖结构特性参数差异,平均数间的多重比较采用Duncans检验方法,P<0.05时差异显著。
2结果与分析
2.1川滇高山栎叶片的基本解剖结构特征
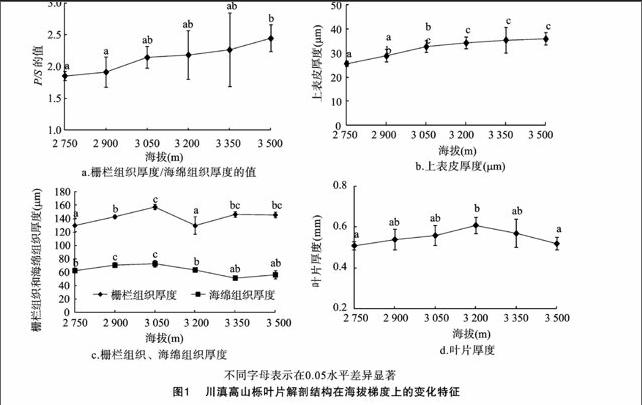
川滇高山栎叶片为异面叶,叶片全缘或有刺状锯齿,由图1 可以看出,川滇高山栎叶片平均厚度为(550.00±0.04)μm,背面有褐色的锈状绒毛。主脉在叶下部凸起,呈不规则半圆形。在光学显微镜下,川滇高山栎叶表面的角质层变化不明显;叶片上表皮由2~3层排列紧密的长方形细胞组成,平均厚度为(32.20±2.76) μm;叶肉细胞中上表面的栅栏组织(P) 和近下表皮面的海绵组织(S)明显分为两部分,且其厚度比值约为2.14。其中,栅栏组织细胞呈柱状,排列紧密,细胞层数变化较大,有1~3层,甚至4层,海绵组织细胞形状和排列不规则,细胞间隙较大,平均厚度为(62.91±3.57) μm;叶片的下表皮也由1层大致呈长方形的细胞组成,但是明显小于上表皮细胞。
2.2川滇高山栎叶片结构特征在海拔梯度上的差异
由图1、图2可以看出,在6个海拔梯度上,叶片厚度随海拔的升高呈现先升后降的趋势,最低海拔2 750 m和最高海拔3 500 m的叶片厚度与海拔3 200 m相比差异显著,叶片厚度在海拔3 200 m附近达到最大,为(0.61±0.04) mm。总体来看,叶片上表皮厚度在(25.73±1.08)~(35.98±2.52) μm之间变化,随海拔的升高呈增加趋势,且叶片上表皮在海拔 3 200 m 处显著增厚。叶肉细胞中栅栏组织和海绵组织厚度的变化范围在(129.61±13.02)~(157.45±3.67)、(5133±3.01)~(72.85±5.08) μm 之间,且随海拔的升高呈现出相似的变化,总体上呈先升高后下降的趋势,这2种组织在海拔3 050 m处显著增厚。栅栏组织与海绵组织的比值随海拔的升高呈增加趋势,且在海拔超过2 900 m时显著增加。
2.3川滇高山栎栅栏组织在海拔梯度上的差异
由表1可以看出,川滇高山栎栅栏组织细胞的长径和短径随海拔的升高呈现出相似的变化趋势,但栅栏组织细胞的长径和短径分别在海拔2 900、3 200 m处达到最大,栅栏组织
细胞短径的变化不显著,而栅栏组织长径在各个海拔之间变化显著。栅栏组织细胞长径与短径的比值随着海拔梯度的变化呈现出上升的趋势,但是各个海拔之间差异不显著。
表1川滇高山栎叶片栅栏细胞组织随海拔梯度的变化
采样点海拔
(m)栅栏组织细胞
长径(μm)栅栏组织细胞
短径(μm)长径/短径的值2 75032.68±2.65a6.35±1.33a1.86±0.07a2 90057.44±2.38d8.01±0.87ab1.92±0.24a3 05044.47±5.55bc7.73±0.41ab2.15±0.17ab3 20041.69±3.35b8.52±2.28b2.19±0.38ab3 35047.68±1.42c8.03±1.31ab2.27±0.58ab3 50049.35±0.69c7.70±0.30ab2.45±0.21bP值0.000***0.357ns0.130ns注:同列数据后不同字母表示差异显著(P<0.05)。
3结论与讨论
海拔是一个重要的地形因子,各种环境因子如气温、气压在海拔梯度上也表现出梯度变化,例如,随着海拔的升高,CO2分压降低,光照强度则增加。由于复杂而多样化的环境和微地形条件导致不同区域植物生活环境的多样化,对植物的生理生态适应产生了不同程度的影响[16]。叶片是植物进化过程中产生的对环境变化比较敏感且可塑性较大的器官,其结构特征最能体现环境因子的影响或植物对环境的适应[17]。川滇高山栎作为一种在玉龙雪山上分布十分广泛的植物,其叶片解剖结构在不同海拔梯度生态环境因子的影响下发生了相应的适应性变化。
在不同海拔的生态环境中,温度和降水量随海拔的变化而成为影响叶肉组织厚度的主要生态因子[18]。在本研究中,随着海拔的升高,川滇高山栎叶片毛、鳞片和叶片的厚度呈升高趋势(图1、图2),这可能是植物对高海拔地区严酷的生态环境适应的结构基础,该结果在王国严等的研究[10]中有所报道。有研究认为,植物的叶片毛、鳞片和叶片厚度对环境因子中的低温、水分散失和紫外辐射有明显的保护作用[7]。此外,在本研究中川滇高山栎叶片上表皮由2层长方形或方形细胞构成,排列紧密,且随着海拔高度的增加而显著升高(图1、图2),下表皮由形状不规则的1层细胞构成,大量的报道认为,这些结构特征可有效地避免水分蒸腾和强辐射对叶肉组织的伤害[19-21],这可能是植物对不同海拔梯度上生态环境因子的长期适应的结构基础。此外,不同海拔区域的温度和干旱引起的胁迫限制了植物的生长。在低海拔地段,干热环境对植物胁迫比较严重,导致植物叶肉细胞的分裂和生长受到抑制;而高海拔地段由于靠近玉龙雪山顶部的雪,其空气湿度和降水量增加,但是温度降低,环境中水热状况有所好转,这种水热条件对植物的生长和发育有利,因此随着海拔梯度的升高,川滇高山栎叶片叶肉和叶片总厚度也相应增加(图1、图2)。相似的结果在李芳兰对四川黄栌(Cotinus szechuanensis)叶片的生态解剖结构特征的研究[18]中也有所报道。
植物的光合作用主要在叶肉细胞中进行,光合作用效率主要受叶肉细胞中的细胞层数、栅栏组织和海绵组织厚度、栅栏细胞的形态及其组成方式等影响[18]。川滇高山栎叶片叶肉细胞显著分化,栅栏组织发达,常有2~3层,有的甚至有4层长柱形栅栏细胞紧密排列(图1、图2)。这可能是对不同海拔梯度上各种环境因子的响应。Esau等认为,小而厚的叶片、发达的栅栏组织、长柱形的栅栏细胞、排列紧密的海绵组织、厚的角质膜、发达的表皮毛以及下皮层结构等是对干燥(可能主要是空气湿度) 或强光生境的适应[22-23];薄的栅栏组织、疏松的海绵组织(大的胞间隙)、近方形的栅栏细胞和凸透形的表皮细胞是对较弱光照环境或中生、湿生环境的适应[24]。另外,栅栏细胞长径和短径与光照关系密切,栅栏细胞长度随海拔升高总体上先上升后下降,这可能是一种对高海拔的严酷环境因子的积极响应。蔡永立等研究认为,环境中的水分含量变化可以通过植物细胞中叶肉栅栏组织与海绵组织的分化程度反映出来[8,19]。植物在收到干旱等逆境胁迫时,植物叶肉细胞结构上常表现为叶肉栅栏组织发达而海绵组织相对减少,进而导致P/S的值发生相应的变化。本研究发现,川滇高山栎叶肉细胞中的栅栏组织细胞层数在海拔梯度上无明显变化,但其厚度随着海拔升高呈现出先升高后下降的趋势,而海绵组织与栅栏组织厚度的变化相似(图1、图2),但是下降幅度较大,因此P/S的值随着海拔上升也呈增加趋势,这可能是由于川滇高山栎叶肉细胞中栅栏组织对不同海拔梯度上生态环境的变化比较敏感。这与安黎哲等对高海拔地区生长火绒草(Leontopodium leontopodiodes)[25]和李芳兰等对白刺花(Sophora davidii)[18]等干旱河谷耐旱灌木的研究结果类似。此外,Cald认为,植物通过增加叶肉细胞中栅栏组织的厚度来应对高海拔地区短波辐射的增强,进而防止由于叶面温度升高引起的组织灼伤[26]。这与本研究中高海拔的川滇高山栎叶片内栅栏细胞厚度较大的结果一致,但是与Chen等的研究结果[19-20]相反。他们在对胡杨等的研究中发现,P/S的值随着海拔的上升而减小。这一方面可能是因为植物所处的生态环境和选择的海拔梯度不同;另外一方面可能是因为所选择的植物间遗传特性有一定的差异或者植物对不同类型的环境变化的适应方式可能存在较大的差异。
综上所述,川滇高山栎叶片的适应性结构特征是多种生态因子长期作用和积累的结果。随着海拔的升高,川滇高山栎受到的胁迫逐渐加重,但是通过叶片解剖结构发生不同程度的变化能形成一定程度的生态适应,即随着海拔上升,叶片厚度、叶肉栅栏组织厚度和P/S的值呈增加趋势,但是栅栏细胞的长径、短径呈降低趋势。在不同海拔高度,叶片解剖结构的这种差异一方面表明不同海拔梯度上复杂的环境条件限制了植物的生长和分布,另一方面体现了川滇高山栎通过叶片解剖结构组织发生不同程度的变化一定程度适应环境的变化。
参考文献:
[1]康华靖,刘鹏,徐根娣,等. 大盘山自然保护区香果树对不同海拔生境的生理生态响应[J]. 植物生态学报,2008,32(4):865-872.
[2]吴栋栋,周永斌,于大炮,等. 不同海拔长白山岳桦的生理变化[J]. 生态学报,2009,29(5):2279-2285.
[3]Chartzoulakis K,Patakas A,Bosabalidis A M. Change in water relations,photosynthesis and leaf anatomy induced by intermittent drought in two olive cultivars[J]. Environ Exp Bot,1999,42:113-120.
[4]Klich M G. Leaf variations in Elaeagnus angustifolia related to environmental heterogeneity[J]. Environ Exp Bot,2000,44:171-183.
[5]李芳兰,包维楷. 植物叶片形态解剖结构对环境变化的响应与适应[J]. 植物学通报,2005,22(S1):118-127.
[6]费松林,方精云,樊拥军,等. 贵州梵净山亮叶水青冈叶片和木材的解剖学特征及其与生态因子的关系[J]. 植物学报,1999,41(9):1002-1009.
[7]贺金生,陈伟烈,王勋陵. 高山栎叶的形态结构及其与生态环境的关系[J]. 植物生态学报,1994,18(3):219-227.
[8]蔡永立,宋永昌. 浙江天童常绿阔叶林藤本植物的适应生态学Ⅰ.叶片解剖特征的比较[J]. 植物生态学报,2001,25(1):90-98,130-131.
[9]戴怡龄,安黎哲,陈拓,等. 寒区不同海拔橘黄罂粟叶片结构特征的比较研究[J]. 西北植物学报,2004,24(3):495-503.
[10]王国严,徐阿生. 川滇高山栎研究综述[J]. 四川林业科技,2008,29(2):23-29,34.
[11]郝晓东. 四川卧龙自然保护区川滇高山栎(Quercus aquifolioides)光合特性研究[D]. 北京:北京林业大学,2006.
[12]Zhang X J,Korpelainen H,Li C Y. Microsatellite variation of Quercus aquifolioides populations at varying altitudes in the Wolong natural reserve of China[J]. Silva Fennica,2006,40(3):407-415.
[13]Li C Y,Wu C C,Duan B L,et al. Age-related nutrient content and carbon isotope composition in the leaves and branches of Quercus aquifolioides along an altitudinal gradient[J]. Trees-Structure and Function,2009,23(5):1109-1121.
[14]吴之坤,张长芹. 滇西北玉龙雪山报春花种质资源的调查[J]. 广西植物,2006,26(1):49-55,68.
[15]李芳兰,包维楷,刘俊华,等. 岷江上游干旱河谷海拔梯度上白刺花叶片生态解剖特征研究[J]. 应用生态学报,2006,17(1):5-10.
[16]胡启武,宋明华,欧阳华,等. 祁连山青海云杉叶片氮、磷含量随海拔变化特征[J]. 西北植物学报,2007,27(10):2072-2079.
[17]王勋陵,王静. 植物的形态结构与环境[M]. 北京:兰州大学出版社,1989:205-230.
[18]李芳兰,包维楷,吴宁. 岷江上游干旱河谷海拔梯度上四川黄栌(Cotinus szechuanensis)叶片的生态解剖[J]. 应用与环境生物学报,2007,13(4):486-491.
[19]Chen Y N,Wang Q,Ruan X,et al. Physiological response of Populus eu phraticato artificial water-recharge of the lower reaches of Tarim River[J]. Acta Bot Sin,2004,46:1393-1401.
[20]容丽,王世杰,刘宁,等. 喀斯特山区先锋植物叶片解剖特征及其生态适应性评价——以贵州花江峡谷区为例[J]. 山地学报,2005,23(1):35-42.
[21]王勋陵,王静. 植物形态与环境[M]. 北京:兰州大学出版社,1989:105-138.
[22]Esau K. An atomy of seed plants[M]. 2nd ed. New York:John Wiley And Sons Press,1977:351-372.
[23]Lee D W,Bone R A,Tersis S. Correlates of leaf optical properties in tropical forest sun and extreme-shade plants[J]. American Journal of Botany,1990(77):370-380.
[24]Bone R E,Lee D,Norman J M. Epidermal cells functioning as lenses in leaves of tropical rain-forest shade plants[J]. Applied Optics,1985(24):1408-1412.
[25]安黎哲,戴怡龄,陈拓,等. 乌鲁木齐河源区不同海拔的火绒草叶片结构特征的比较研究[J]. 冰川冻土,2004,26(4):474-481.
[26]Cald M M. Solar UV irradiation and the growth and development of higher plants[C]//Giess A C.Photo-physiology. New York:Academic Press,1993:125-143.唐探,姜永雷,张瑛,等. 铅、镉胁迫下云南樟幼苗的生理特性变化[J]. 江苏农业科学,2015,43(1):199-201.
