黄绿木霉菌发酵液对水稻防御酶的影响
2015-04-17林志伟孙冬梅迟丽
林志伟+孙冬梅+迟丽

摘要:以黄绿木霉菌在马铃薯几丁质营养液中培养96 h获得的发酵滤液为试验药剂,进行室内稻瘟病菌孢悬液人工接种与防治处理,测定水稻植株各防御性关键酶的变化规律,研究黄绿木霉菌发酵液对水稻植株抗病性的诱导能力。结果表明,喷施发酵液的水稻植株,稻瘟病病斑比对照减小;不同处理间水稻植株酶活性变化不同,喷施发酵液并接种病菌的水稻植株各种防御酶活性峰值出现较早,接种12 h,多酚氧化酶(PPO) 含量是对照的4倍,苯丙氨酸解氨酶(PAL)含量比对照高25%,绿原酸含量比对照高60%;接种48 h,过氧化物酶(POD)含量是对照的8倍。黄绿木霉菌几丁质发酵液具有抗御稻瘟病菌侵染和提高防御酶活性的作用。
关键词:黄绿木霉菌;几丁质;稻瘟病;防御酶;多酚氧化酶;苯丙氨酸解氨酶;过氧化物酶
中图分类号: S435.11文献标志码: A文章编号:1002-1302(2015)01-0121-03
收稿日期:2014-03-31
基金项目:黑龙江省齐齐哈尔市科技计划(编号:NYGG2010-09)。
作者简介:林志伟(1970—),男,黑龙江勃利人,硕士,副教授,主要从事植物病虫害综合防治技术研究。Tel:(0459)6819182;E-mail:lzwsdm@sohu.com。几丁寡糖为功能型低聚糖,水溶性好且易被分散吸收,具有多种生理和生物学功能,尤其是可提高植物对病原菌的抵抗能力,启动和激发植物的防御系统[1],目前已被广泛应用到农业生产领域[2]。几丁寡糖可由几丁质通过酸解法、氧化法和酶解法降解产生[3]。传统化学方法降解几丁质的反应条件难以控制,反应稳定性和重复性较差,易造成环境污染。利用生物发酵技术降解几丁质成为当前研究热点,至今,已发现包括细菌、真菌和植物[4]在内的许多生物能产生几丁质降解酶。黄绿木霉菌(Trichodema aureoviride)对几丁质有较好的降解能力[5],对水稻纹枯病菌[Thanatephorus cucumeris (Frank) Donk]、稻瘟病菌(Pyricularia grisea)、核盘菌[Sclerotinia sclerotiorum (Lib.) de Bary]等有良好的应用潜能[6-8]。有研究表明,微生物代谢产生的几丁寡糖可以诱导植物提高抗病能力[9-12],但有关黄绿木霉菌代谢产物诱导抗病能力的产生还未见报道。研究黄绿木霉菌在含几丁质营养液中发酵所得滤液对水稻植株防御酶系的影响,探索黄绿木霉发酵几丁质产生几丁寡糖的利用途径,明确黄绿木霉菌防病作用机理,对开拓植物病害的生物防治资源具有重要意义。
1材料与方法
1.1供试材料
供试菌种黄绿木霉菌T4、水稻稻瘟病菌ZD1,由黑龙江八一农垦大学植物保护系提供;试验水稻品种为垦鉴稻5号,由黑龙江八一农垦大学农学院水稻中心提供。改良马铃薯几丁质发酵培养液为:马铃薯200 g、葡萄糖5 g、K2HPO4 0.7 g、KH2PO4 0.3 g、KCl 0.01 g、FeSO4 0.01 g、MgSO4 0.5 g、胶态几丁质8 g,用蒸馏水定容至1 000 mL,分装于500 mL三角瓶中。
1.2试验方法
1.2.1黄绿木霉菌的发酵培养及发酵滤液获得将孢子浓度为2.5×107个/mL的黄绿木霉菌孢悬液,接种于改良马铃薯几丁质培养液中,28 ℃、120 r/min恒温振荡培养4 d;过滤除去菌体,即得木霉菌几丁质发酵滤液。
1.2.2水稻植株栽培与处理方法将垦鉴稻5号秧苗插栽于直径25 cm、高20 cm的花盆中,按三角位置每盆插栽3穴,1株/穴;每日观察花盆内水量,适当补水,保证水稻正常生长至孕穗期;在水稻孕穗期进行喷雾接种。试验设喷施发酵滤液不接种稻瘟病菌孢子悬液、喷施清水并接种稻瘟病菌孢子悬液、喷施发酵滤液2 h后接种稻瘟病菌孢子悬液、喷施清水不接种稻瘟病菌孢子悬液为空白对照共4个处理,接种稻瘟病菌孢子悬液浓度为1.6×108个/mL,于16:00—17:00,使用手执式喷雾器在水稻植株上方均匀喷雾;分别将每盆水稻用有机玻璃罩保湿36 h,除去有机罩,每日观察水稻叶片病斑出现情况;处理结束后12、24、48、72、96、122、144 h,剪取水稻最上部2张叶片,立即用天平称质量,并按文献[13]方法测定水稻叶片过氧化物酶(POD)、多酚氧化酶(PPO)、苯丙氨酸解氨酶(PAL)及绿原酸等指标。
2结果与分析
2.1黄绿木霉菌几丁质发酵滤液对水稻稻瘟病的控制作用
试验结果表明,在水稻植株接种稻瘟病菌并保湿,各水稻植株叶片上均有稻瘟病病斑出现,病斑出现的时间和病斑数量没有差别,病斑均为典型的稻瘟病急性病斑,病斑较小;喷施发酵滤液的植株,稻瘟病病斑周围的黄色晕圈不明显,而只接种病原菌的水稻叶片,病斑周围有明显的黄色晕圈,病斑向两端延伸明显(表1)。经发酵滤液处理的水稻叶片发病程度减轻,一定程度上抑制了病原菌生长,限制了病原菌对水稻叶片的危害。表1喷施黄绿木霉菌几丁质发酵液对稻瘟病斑的影响
处理病斑长直径
(mm)病斑短直径
(mm)黄晕宽度
(mm)喷施清水并接种稻瘟病孢子悬液3.62a0.84a0.45a喷施发酵液2 h后接种稻瘟病孢子悬液1.12b0.65a0.12b
2.2黄绿木霉菌几丁质发酵滤液对水稻抗病关键酶活性的影响
2.2.1过氧化物酶的活性变化植物体内过氧化物酶活性与植物抗病性呈正相关[14]。试验结果表明,未喷施发酵滤液而接种稻瘟病菌的水稻新叶,POD酶在接种12~72 h内与对照在同一水平范围波动,72 h后发生变化,96 h时POD酶活性大幅跃升,并在96~144 h维持一个较高水平;喷施发酵滤液后接种病菌的水稻植株,叶片POD酶活性在12~48 h内提高幅度很大,24 h时POD酶活性是对照的近7倍,48 h达到近8倍水平,48 h后开始下降,但叶片酶活水平仍略高于对照(图1)。这是由于病原菌侵染水稻植株叶片后,水稻植株对病原菌产生了抗逆反应[15],施用发酵滤液,在一定程度上使叶片POD酶活性提高且抗逆反应提前,有利于保护植物免受病原菌进一步侵染[16]。
2.2.2多酚氧化酶的活性变化PPO是一类广泛分布于植物体内能催化多酚类氧化成醌类的质体金属酶,其活性提高有助于提高植物的抗病性[17-18],对病原菌侵染起到一定限制作用。试验结果表明,仅接种稻瘟病病菌的水稻叶片,12 h时PPO酶活性几乎为0,48 h达到最高值,后逐渐下降;清水对照组,水稻叶片PPO酶活性在12~144 h较低,而仅喷施滤液的叶片,PPO酶活性在12 h有显著提高,并持续至72 h,72~144 h 后稍有下降,但仍高于清水对照组(图2),这说明喷施滤液可提高植物的抗病性。
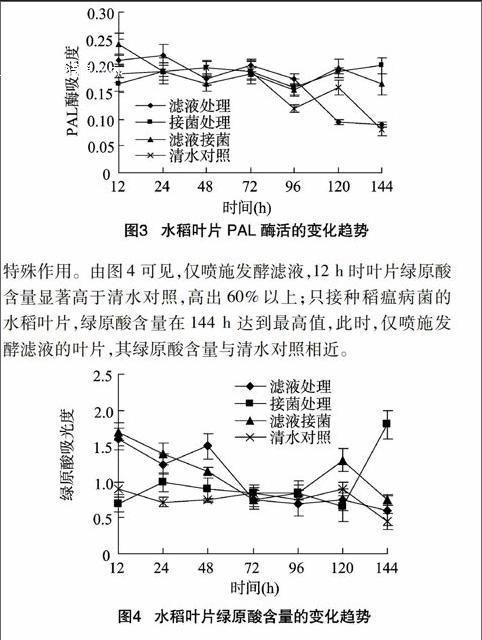
2.2.3苯丙氨酸解氨酶的活性变化PAL是植保素、木质素和酚类化合物合成的关键酶,与系统获得抗性表现存在相关性。试验结果表明,不同处理水稻叶片,在PAL酶活达到高峰后均呈逐渐下降趋势;PAL初期变化趋势与PPO相似,12 h时,各处理水稻叶片PAL表现出明显差异,仅喷施滤液和喷施滤液2 h后接种的水稻叶片,其PAL酶活性比清水对照和只接种病菌分别高25%和10%;仅接种病菌的水稻叶片,其PAL活性在接种120 h后有所升高(图3)。这进一步表明,喷施滤液对植株抗病性的提高有促进作用。
2.2.4绿原酸的活性变化绿原酸能够清除自由基,具有抑制IAA氧化酶对植物生长素的破坏[19],在植物抗病方面起着
特殊作用。由图4可见,仅喷施发酵滤液,12 h时叶片绿原酸含量显著高于清水对照,高出60%以上;只接种稻瘟病菌的水稻叶片,绿原酸含量在144 h达到最高值,此时,仅喷施发酵滤液的叶片,其绿原酸含量与清水对照相近。
3结论
过氧化物酶是植物体内普遍存在且活性较高的一种酶,是植物体内的保护酶之一,可以清除细胞内的H2O2,与植物呼吸作用、光合作用、生长素氧化及木质素形成等有关,其活性可以反映某一时期植物体内的代谢及抗逆性变化[14]。苯丙氨酸解氨酶是植物体内苯丙烷代谢途径的关键酶,其活性高低控制着植物体内多种次生酚类化合物、类黄酮植保素、木质素等抗菌物质的合成,在植物次生代谢和抗病代谢中具有重要的作用[20-21]。多酚氧化酶是一类广泛存在于植物体内的含铜金属酶类,与作物的褐化有关,可以提高植物对病原菌的抗性[22-23]。本研究结果表明,当水稻叶片受稻瘟病菌协迫后,黄绿木霉菌几丁质发酵滤液能够诱导激活水稻叶片内PAL、PPO、POD等几种抗病关键酶的活性,使酶活力有所提高,且叶片内绿原酸含量也得到显著提高。黄绿木霉菌几丁质发酵滤液对稻瘟病具有一定的防治作用。
参考文献:
[1]van der Holst P P G,Schlaman H R M,Spaink H P. Proteins involved in the production and perception of oligosaccharides in relation to plant and animal development[J]. Current Opinion in Structural Biology,2001,11(5):608-616.
[2]王扬,娄永江,杨文鸽. 酶法制备几丁寡糖和壳寡糖研究现状与进展[J]. 东海海洋,2001,19(4):40-45.
[3]乔兴忠,李永娴,汤熙翔. 发酵法产酶生产几丁寡糖的工艺研究[J]. 厦门大学学报:自然科学版,2010,49(2):251-255.
[4]竺国芳,赵鲁杭.几丁寡糖和壳寡糖的研究进展[J]. 中国海洋药物,2000,19(1):43-46.
[5]林志伟,张海燕,孙冬梅,等. 黄绿木霉菌菌株产几丁质酶发酵条件的研究[J]. 安徽农业科学,2011,39(9):5043-5044.
[6]林志伟,靳学慧. 四种木霉菌对稻瘟病菌作用能力的比较研究[J]. 黑龙江八一农垦大学学报,2006,18(4):8-12.
[7]孙冬梅,林志伟. 黄绿木霉发酵液对水稻纹枯病菌作用的研究[J]. 植物保护,2009,35(4):83-86.
[8]孙冬梅,杨谦,宋金柱.黄绿木霉菌代谢产物对大豆菌核病核盘菌的抑菌能力研究[J]. 天然产物研究与开发,2005,17(6):691-695.
[9]戚益平,何逸建,许煜泉.根际细菌诱导的植物系统抗性[J]. 植物生理学通讯,2003,39(3):273-278.
[10]王淑霞,张丽萍,黄亚丽,等. 哈茨木霉Tr-92诱导黄瓜对灰霉病系统抗性的研究[J]. 中国生物防治学报,2013,29(2):242-247.
[11]黄艳青,庄敬华,高增贵,等. 木霉菌诱导甜瓜抗枯萎病相关防御反应酶系的研究[J]. 沈阳农业大学学报,2005,36(5):546-549.
[12]Olson H A,Benson D M. Induced systemic resistance and the role of binucleate Rhizoctonia and Trichoderma hamatum 382 in biocontrol of botrytis blight in geranium[J]. Biological Control,2007,42(2):233-241.
[13]王学奎.植物生理生化实验原理和技术[M].2版. 北京:高等教育出版社,2006.
[14]葛秀春,宋凤鸣,郑重.稻瘟菌侵染后水稻幼苗活性氧的产生与抗病性的关系[J]. 植物生理学报,2000,26(3):227-231.
[15]马国华,王铨茂.水稻品种过氧化物酶活性和木质素含量与抗稻瘟病菌的关系[J]. 植物生理学通讯,1992,28(4):264-267.
[16]孙万春,薛高峰,张杰,等. 硅对水稻防御性关键酶活性的影响及其与抗稻瘟病的关系[J]. 植物营养与肥料学报,2009,15(5):1023-1028.
[17]贺立红,宾金华. 高等植物中的多酚氧化酶[J]. 植物生理学通讯,2001,37(4):340-345.
[18]赵伶俐,范崇辉,葛红,等. 植物多酚氧化酶及其活性特征的研究进展[J]. 西北林学院学报,2005,20(3):156-159.
[19]郭秋娟,金邦荃,陈和平. 绿原酸生物活性与提取方法的研究进展[J]. 食品工业科技,2009,30(8):346-348.
[20]向妙莲,何永明,付永琦,等. 茉莉酸甲酯对水稻白叶枯病的诱导抗性及相关防御酶活性的影响[J]. 植物保护学报,2013,40(2):97-101.
[21]Latha P,Anand T,Ragupathi N,et al. Antimicrobial activity of plant extracts and induction of systemic resistance in tomato plants by mixtures of PGPR strains and zimmu leaf extract against Alternaria solani[J]. Biological Control,2009,50(2):85-93.
[22]张智慧,聂燕芳,何磊,等. 外源茉莉酸甲酯诱导水稻抗瘟性相关防御酶和内源水杨酸的变化[J]. 植物病理学报,2010,40(4):395-403.
[23]黄凤莲,戴良英,罗宽. 药剂诱导水稻幼苗抗寒机制研究[J]. 作物学报,2000,26(1):92-97.
