嵊泗列岛海域三种贻贝贝体框架性状对壳重的影响效应*
2015-04-10郑晓静邹李昶任夙艺刘祖毅王志铮
郑晓静 杨 阳 邹李昶, 任夙艺 刘祖毅 王志铮①
(1. 浙江海洋学院 舟山 316022; 2. 余姚市水产技术推广中心 余姚 315400; 3. 嵊泗县海洋与渔业局 嵊泗 202450)
“渔业碳汇”是指通过渔业生产活动促进水生生物吸收水体中的CO2, 并通过收获将所固定的碳移出水体的过程和机制, 是 CO2减排的重要组成部分(唐启升, 2010)。据联合国《蓝碳》报告, 地球上55%的生物碳捕获由海洋生物完成(刘慧等, 2011)。贝类作为近海海洋生物泵的重要环节, 一方面借助碳酸钙(CaCO3)泵直接吸收海水中的碳酸氢根(HCO3–)形成碳酸钙来固碳(张朝晖等, 2007), 另一方面通过高效滤取水体中的悬浮颗粒有机碳以促进软体部的增长(张继红等, 2005), 是浅海区固碳增汇的重要生物类群之一。以紫贻贝为例, 其贝壳和软体部中的碳含量就分别达12.68%和45.98% (周毅等, 2002)。研究表明, 大规模的贝类养殖活动对水体中悬浮颗粒有机物质的数量以及组成有一定的控制作用(Kasparet al, 1985;Young, 1993; Prinset al, 1995; 董双林等, 1999;Nakamuraet al, 2000), 养殖贝类对黄海海洋生态系统的固碳贡献率达 0.46%, 而野生贝类则仅为 0.02%(刘慧等, 2011)。无疑, 大力发展浅海贝类养殖产业对于提升养殖海域海洋生态系统的服务功能具有十分重要的现实意义。
嵊泗列岛海域既是我国厚壳贻贝(Mytilus coruscus)的重要原产地, 也是浙江省贻贝养殖规模最大的区域(养殖面积达1733hm2), 养殖对象为紫贻贝(Mytilus edulis)、厚壳贻贝以及因杂交或基因渐渗(introgressive hybridization)(Andersonet al, 1938)而出现的少量“杂交贻贝” (张义浩等, 2003; 沈玉帮等, 2006; 白晓倩等, 2014)。据报道, 滤食性贝类贝壳的碳含量为贝壳干重的 12%, 且不同海区和种类之间的差异不显著(张继红等, 2005), 贻贝壳长与壳重的增长基本是一致的或略有前后(王如才等, 1993), “杂交贻贝”通过显著提高贝体框架特征中滤食功能区占比值来强化其杂种生长优势, 贝体滤食水平剖面功能区占比较消化功能区占比在表征紫贻贝、厚壳贻贝、“杂交贻贝”间贝体框架特征相似性程度上更具影响力(白晓倩等, 2014), 表明贝体框架特征作为贝类外部形态信息的综合反映和种质遗传规定性的外在体现, 既左右着贝类滤食功能区和消化功能区的空间配置, 也深刻影响着贝类的壳重增长及其固碳增汇趋势。鉴于此, 本文作者于2011年10月22日以厚壳贻贝同生群养殖个体、同域生长的紫贻贝同生群养殖个体以及混于厚壳贻贝和紫贻贝养殖筏架中的“杂交贻贝”为实验对象, 采用多元统计方法开展了三者贝体框架性状对壳重的影响效应研究, 并由此探析了三者间壳重增长对策的差异, 旨为贻贝科物种固碳生物学研究和嵊泗海域“渔业碳汇”产业开发提供基础资料。
1 材料与方法
1.1 样品来源
本研究所用样品均由嵊泗县金盟海水养殖专业合作社提供, 采自该社筏式养殖的枸杞岛干斜村邻近海域(32°42′16″—32°42′38″N, 122°45′29″—122°45′53″E)。实验对象为以人工培育的 2+龄厚壳贻贝同生群养殖个体(野生亲贝源自非养殖海区, 稚贝出池时间为2009年6月、海区中间培育时间为2009年6月至2010年 4月, 大规格苗种筏式养殖起始时间为 2010年 4月)、同域生长的1+龄紫贻贝同生群养殖个体(苗种源自大连海区)以及混于厚壳贻贝和紫贻贝养殖筏架中的“杂交贻贝”。
1.2 样品贝壳表型性状参数值的测定
样品运回实验室后, 3种实验贝均随机选取其中壳形完整的112枚活体作为测定群体。测定样品经清除壳表附着物、蒸煮并去除软体部及闭壳肌、用定性滤纸吸干壳表水分后, 用 BS223S型电子天平(精度1mg) 逐枚称量壳重(G), 并在壳内面逐一编号保存备用; 用数显游标卡尺(精度 0.02mm)依次测量壳宽SW (左右两壳紧密时的最大距离)、壳长SL (壳前、后端间的最大水平距离)后, 采用白晓倩等(2014)的方法依次测量壳高 SH (BD, 壳背面最高点至腹缘的最短距离)、OA (壳顶至韧带末端的直线距离)、OB (壳顶至壳背面最高点的直线距离)、OC (壳顶至壳后端最远点的直线距离)、OD (壳顶至壳高性状在腹缘的落点的直线距离)、AB (韧带末端至壳背缘最高点的直线距离)、BC (壳背缘最高点至壳后端最远点的直线距离)、CD (壳后端最远点至壳高性状在腹缘的落点的直线距离)等 8项贝体框架性状。具体测量部位如图1所示。
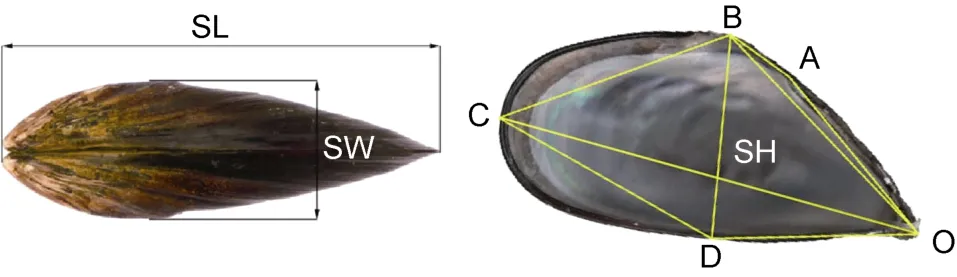
图1 实验贝贝体框架性状的测量部位Fig.1 Schematic of the shell frame characteristics measured in Mytilus
1.3 分析方法
整理所测结果, 借助SPSS 17.0软件计算实验贝壳重(G)及SW、SL、SH、OA、OB、OC、AB、BC、CD等9项贝体框架性状的均值、标准差以及变异系数; 以贝体框架性状为自变量, 在开展它们与壳重间相关分析和对壳重通径分析的基础上, 剖析那些通径系数达到显著水平(P<0.05)的贝体框架性状对壳重的直接作用和间接作用, 进而计算决定系数和复相关指数, 确定影响壳重的关键变量组合; 通过偏回归分析, 保留偏回归系数达到显著水平(P<0.05)的贝体框架性状, 建立这些贝体框架性状对壳重的多元回归方程, 并进行拟合度检验。
2 结果与分析
2.1 实验贝贝壳表型性状的参数估计值
由表1可见, 紫贻贝、厚壳贻贝间的各项贝壳表型性状变异系数均较接近, 且均远低于对应的“杂交贻贝”, 3种实验贝壳重性状变异系数均远大于所涉各项贝体框架性状, 表明本研究所涉3种实验贝在壳重这一贝壳固碳性状上均具较强的选择潜力, “杂交贻贝”贝体框架性状的强可塑性致使其壳重性状的变异系数明显被放大。
2.2 实验贝表型性状间的相关分析
由表2、表3、表4可见, 3种实验贝所测各项贝体框架性状均与其壳重呈极显著正相关(P<0.01); 3种实验贝中, 除厚壳贻贝AB-OA、AB-BC、AB-CD组合和紫贻贝AB-OA、AB-CD、OD-BC、OD-CD组合的相关性均未达到显著水平(P>0.05)外, 其余两贝体框架性状组合的相关性均呈显著水平(P<0.05); 厚壳贻贝、紫贻贝和“杂交贻贝”中, 与其壳重呈高度相关的贝体框架性状(相关系数大于或等于0.75)对壳重性状的影响力排序依次为 OC > SL > OB > SH(BD) > SW、OC > SL> SW 和 SL > OC > SH(BD) > OB > BC > CD > OD >SW > OA; 3种实验贝所测各项表型性状间的相关系数基本上均呈“杂交贻贝”> 厚壳贻贝>紫贻贝。表明本研究所选贝体框架性状包含了壳重决定和种间框架形态甄别双重信息, 对它们进行相关分析具有重要意义。
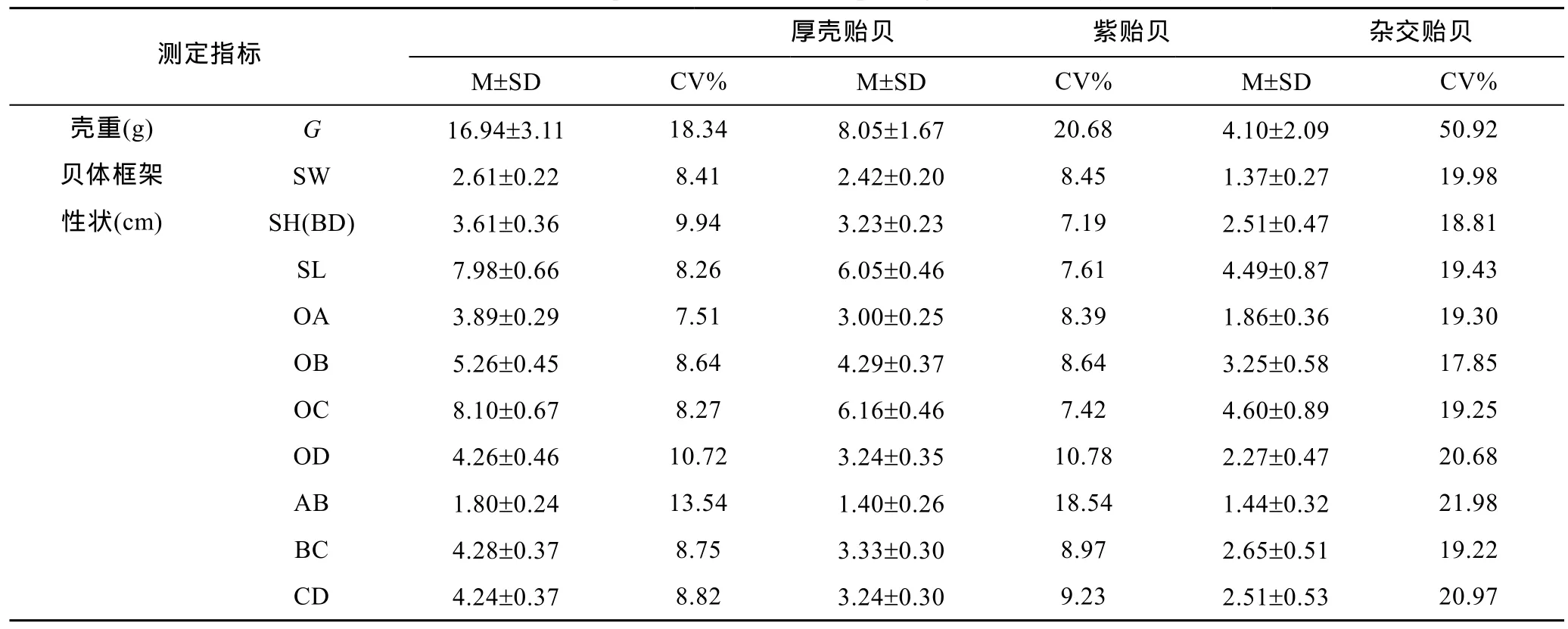
表1 实验贝贝壳表型性状的统计量描述Tab.1 The descriptive statistics of morphological traits of Mytilus
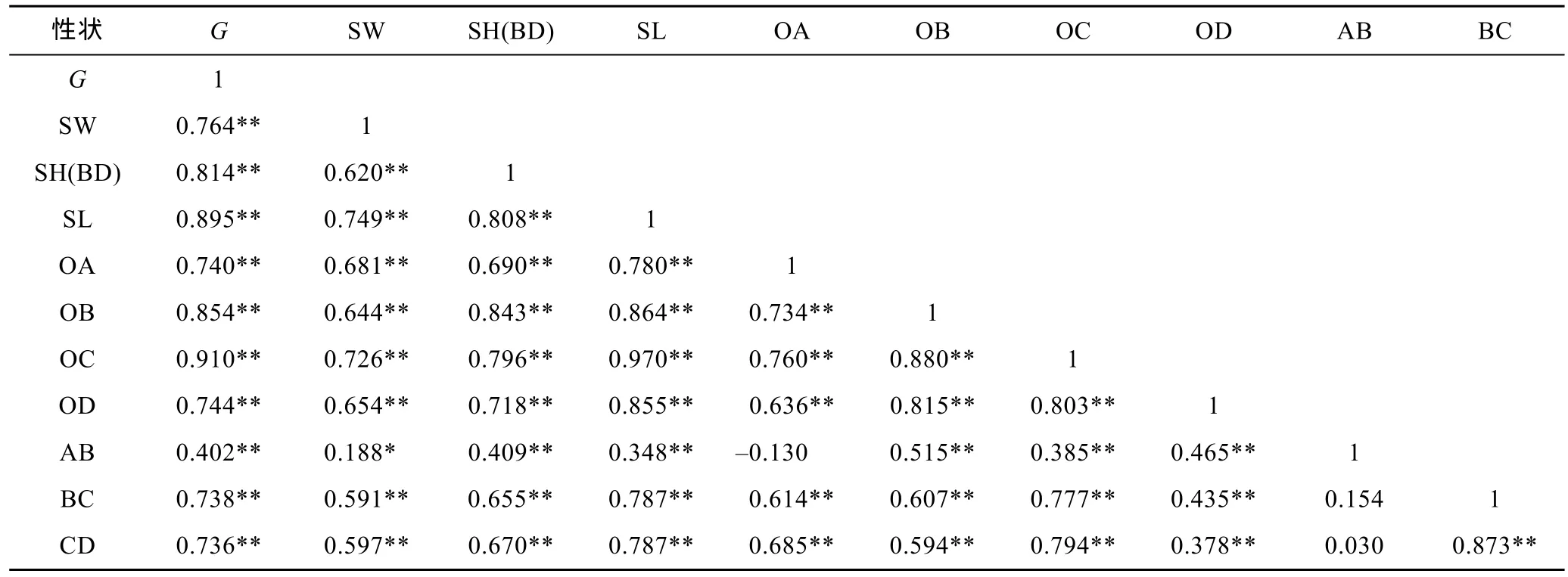
表2 厚壳贻贝贝壳表型性状间的相关系数Tab.2 The correlation coefficients between the phenotype traits of M. coruscus
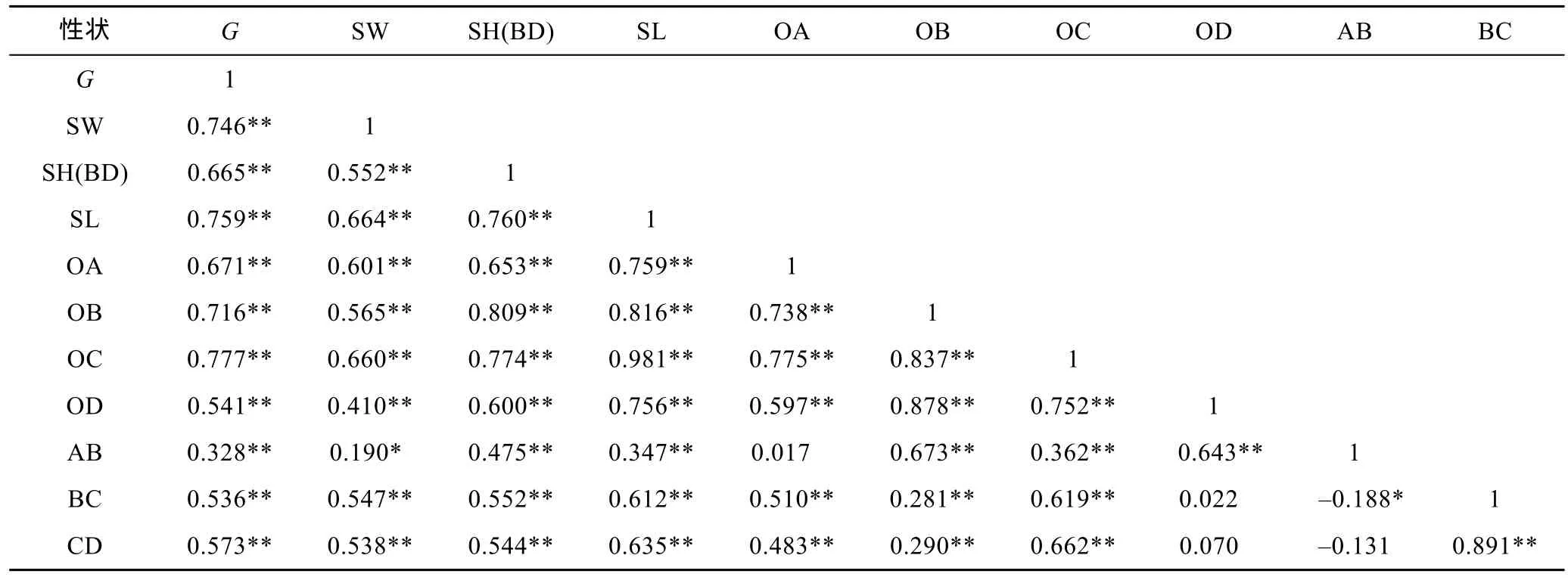
表3 紫贻贝贝壳表型性状间的相关系数Tab.3 The correlation coefficients between the phenotype traits of M. edulis
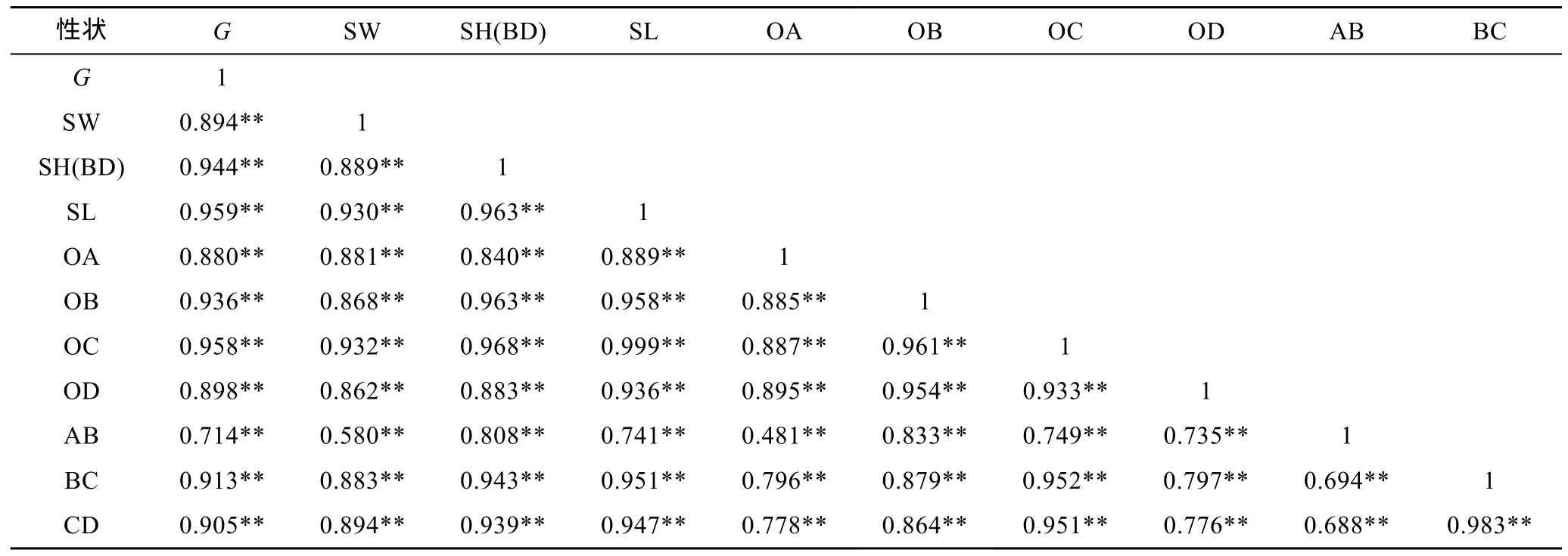
表4 “杂交贻贝”贝壳表型性状间的相关系数Tab.4 The correlation coefficients between the phenotype traits of the hybrid mussel
2.3 实验贝贝体框架性状对壳重的通径分析
根据通径分析原理, 分别获得厚壳贻贝、紫贻贝和“杂交贻贝”测定群体贝体框架性状对壳重的通径系数。经显著性检验, 剔除未达到显著水平的性状(P>0.05), 并按相关系数组成效应, 将保留下来的各贝体框架性状与壳重的相关系数(rij)剖分为直接作用(通径系数P1)和通过其它性状的间接作用(P2)两部分,并列结果于表5。
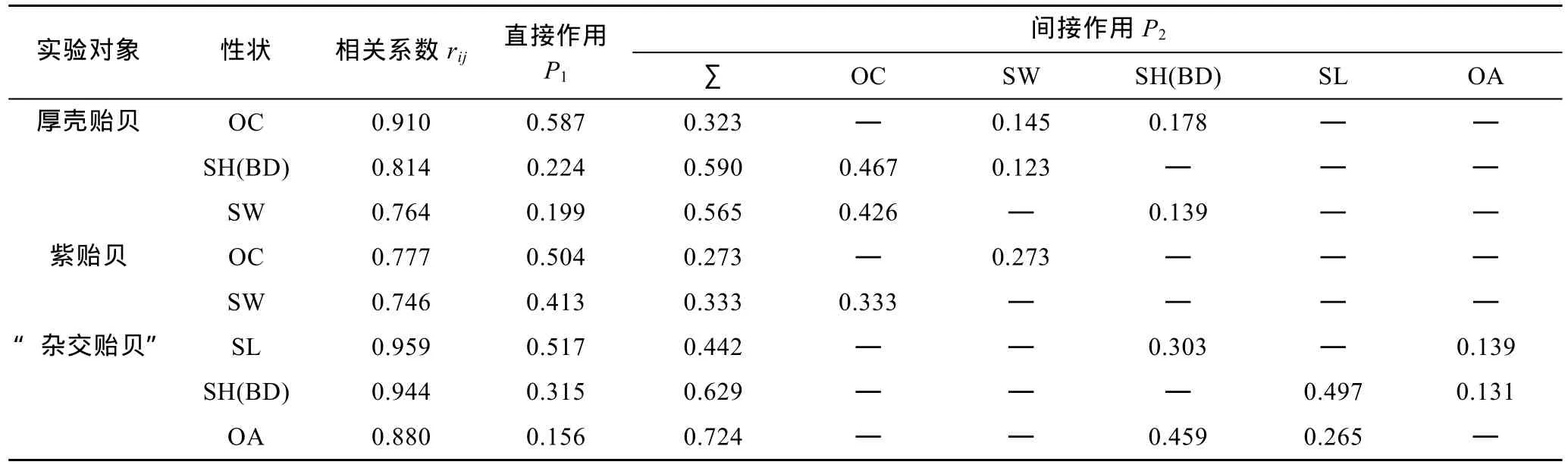
表5 实验贝贝体框架性状对壳重的通径分析Tab.5 The path analysis of the phenotypic traits to the shell weight of Mytilus
由表2和表5可见, 厚壳贻贝测定群体被保留的3个贝体框架性状对壳重的相关系数、直接作用和间接作用分别呈 OC > SH(BD) > SW、OC > SH(BD) >SW和SH(BD) > SW > OC, 除OC对壳重的直接作用明显大于间接作用外, 另外2个性状的直接作用均明显小于间接作用, 且SH(BD)、SW对壳重的间接作用均主要通过OC来实现, 而OC通过SH(BD)、SW对壳重的间接作用则均较小, 表明OC是影响壳重的核心变量, 而 SH(BD)、SW 则均为影响壳重的从属变量。经计算, 三者对壳重的相关指数为0.868。
由表3和表5可见, 紫贻贝测定群体被保留的2个贝体框架性状对壳重的相关系数、直接作用和间接作用分别呈 OC > SW、OC > SW 和 SW > OC, 它们对壳重的直接作用均大于间接作用, 表明OC是影响壳重的核心变量, SW 为影响壳重的重要变量。经计算, 两者对壳重的相关指数为0.700。
由表4和表5可见, “杂交贻贝”测定群体被保留的3个贝体框架性状对壳重的相关系数、直接作用和间接作用分别呈 SL > SH(BD) > OA、SL > SH(BD) >OA和OA > SL > SH(BD), 除SL对壳重的直接作用明显大于间接作用外, 另外2个性状的直接作用均明显小于间接作用, 其中 SH(BD)对壳重的间接作用主要通过SL来实现, 而SL和OA对壳重的间接作用则均主要通过SH(BD)来实现, 表明SL是影响壳重的核心变量, SH(BD)是影响壳重的重要变量, 而OA则是影响壳重的从属变量。经计算, 三者对壳重的相关指数为0.931。
2.4 实验贝贝体框架性状对壳重的决定程度分析
计算单个性状对壳重的决定系数(di=Pi2,Pi为性状对体重的通径系数)和性状两两交互对壳重的共同决定系数(dij= 2rijPiPj,rij为两性状间的相关系数,Pi、Pj分别为两性状对体重的通径系数)并列结果于表6。由表6可见, 厚壳贻贝测定群体OC、SH(BD)、SW 对壳重的相对决定程度依次为 34.4%、4.0%和5.0%, 性状两两交互对壳重的共同决定系数呈 OCSH(BD) > OC-SW > SH(BD)-SW, 均小于 OC 的决定系数; 紫贻贝测定群体OC、SW对壳重的相对决定程度依次为 25.4%和 17.1%, 两者交互对壳重的共同决定系数略大于OC而明显大于SW;“杂交贻贝”测定群体 SL、SH(BD)、OA对壳重的相对决定程度依次为26.7%、9.9%和 2.4%, 除SL-SH(BD)组合外, 其余性状两两交互对壳重的共同决定系数均小于SL的决定系数。上述结果进一步表明, 贝体长度性状是决定厚壳贻贝、紫贻贝和“杂交贻贝”壳重的核心变量,OC-SW、SL-SH(BD)组合分别对紫贻贝和“杂交贻贝”的壳重影响具较强的协同效应。
由表6可知, 厚壳贻贝测定群体3个性状对壳重决定系数的加和为 0.868, 紫贻贝测定群体 2个性状对壳重决定系数的加和0.700, “杂交贻贝”测定群体3个性状对壳重决定系数的加和为 0.931, 均等于其对应的相关指数R2值, 表明这些性状均为影响壳重的主要性状, 较其它性状对壳重的影响更具重要性。
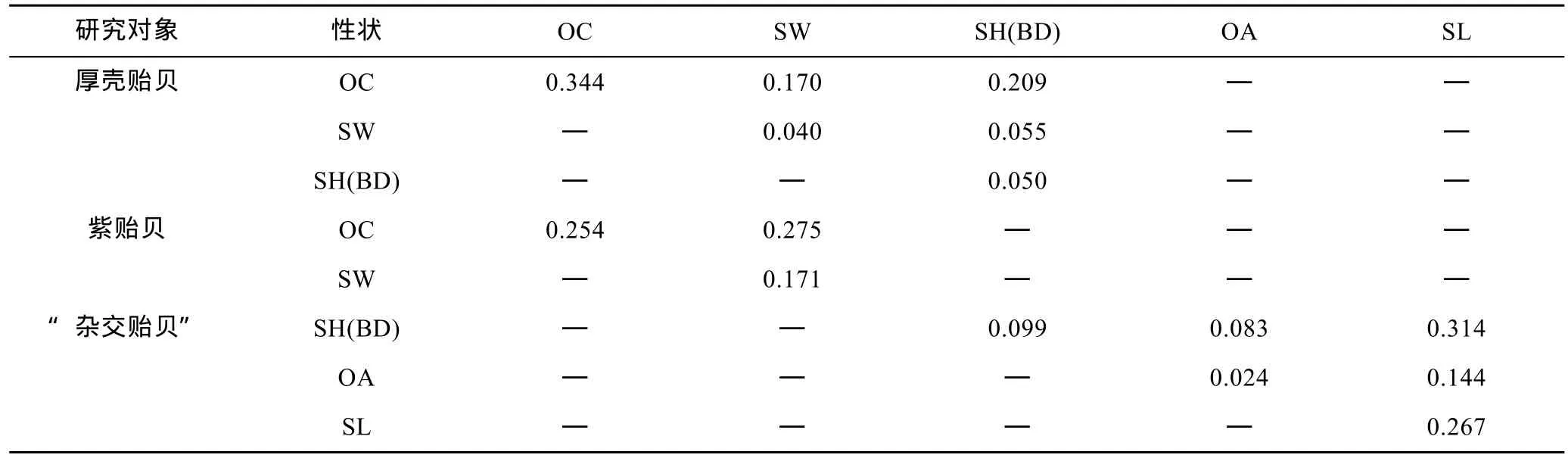
表6 实验贝贝体框架性状对壳重的决定系数Tab.6 The determination coefficients of phenotypic traits on the shell weight of Mytilus
2.5 实验贝贝体框架性状与壳重间多元回归方程的建立
统计实验所测数据并经通径分析和多元回归分析, 在剔除对壳重的偏回归系数不显著的贝体框架性状(P>0.05)后, 再次进行复相关分析和回归分析并将所得结果分别列于表7、表8和表9。
复相关系数系测量目标变量与其它相关变量组合间线性相关程度的综合性指标, 其数值愈大, 表明目标变量与其它相关变量组合间的关系就愈密切。由表7可知, 厚壳贻贝、紫贻贝和“杂交贻贝”测定群体被保留的贝体框架性状与壳重间的复相关系数均达到极显著水平(R>r0.01), 表明它们与壳重具极为密切的关系; 经检验, 厚壳贻贝测定群体中被保留的 3个贝体框架性状对壳重的复相关指数为 0.868, 紫贻贝测定群体中被保留的 2个贝体框架性状对壳重的复相关指数为 0.700, “杂交贻贝”测定群体中被保留的3个贝体框架性状对壳重的复相关指数为0.931。
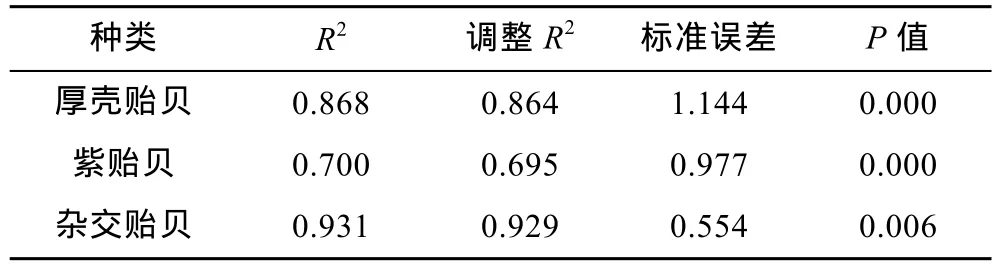
表7 实验贝贝体框架性状与壳重的复相关分析Tab.7 The multiple-correlation coefficients of phenotypic traits on the shell weight of Mytilus
由表8可见, 厚壳贻贝、紫贻贝和“杂交贻贝”测定群体贝体框架性状与壳重间的多元回归方程分别为G= 2.709OC + 2.351SH(BD) + 2.799SW-21.343,G=1.903OC + 3.549SW-12.251和G= 1.235SL + 1.395SH(BD) + 0.908OA-6.634, 回归截距及所有贝体框架性状的偏回归系数均达到极显著水平(P<0.01)。经方差分析表明(表9), 所建立的回归方程的回归关系也均达到极显著水平(P<0.01)。经回归预测, 估计值和实测值间无显著差异(P>0.05), 表明所建方程能够精准反映本研究所涉3种实验贝贝体框架性状与壳重间的相互关系。
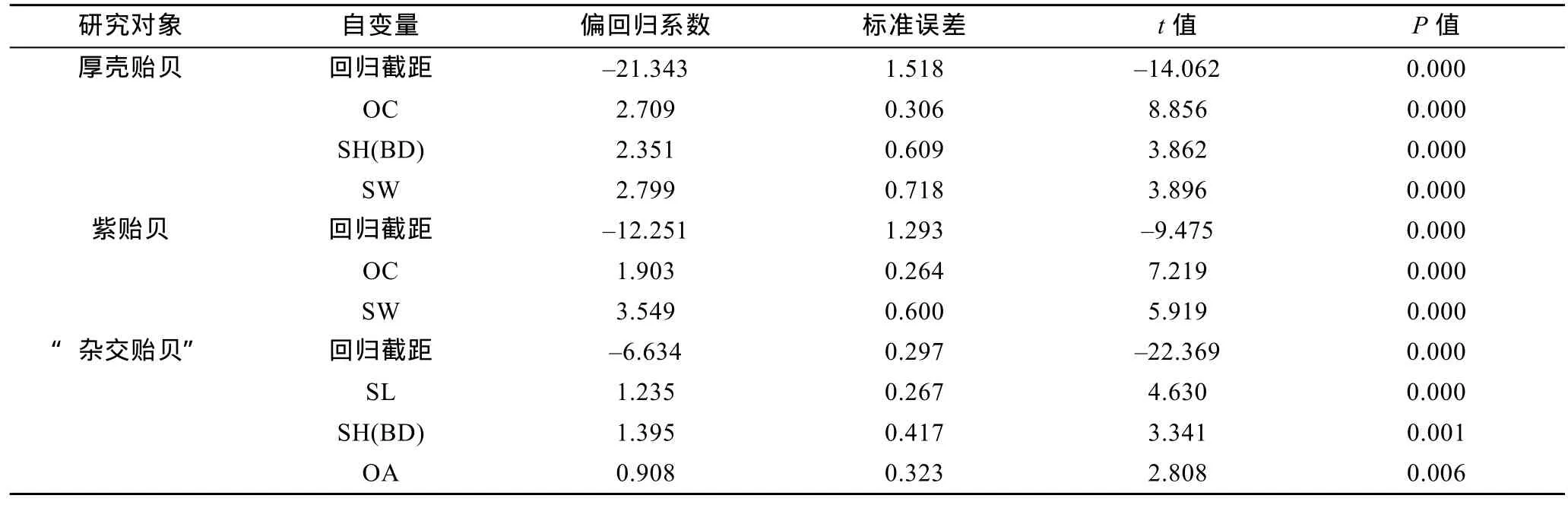
表8 实验贝贝体框架性状与壳重的偏回归系数检验Tab.8 Test on the significance of the partial regressions and constant of Mytilus
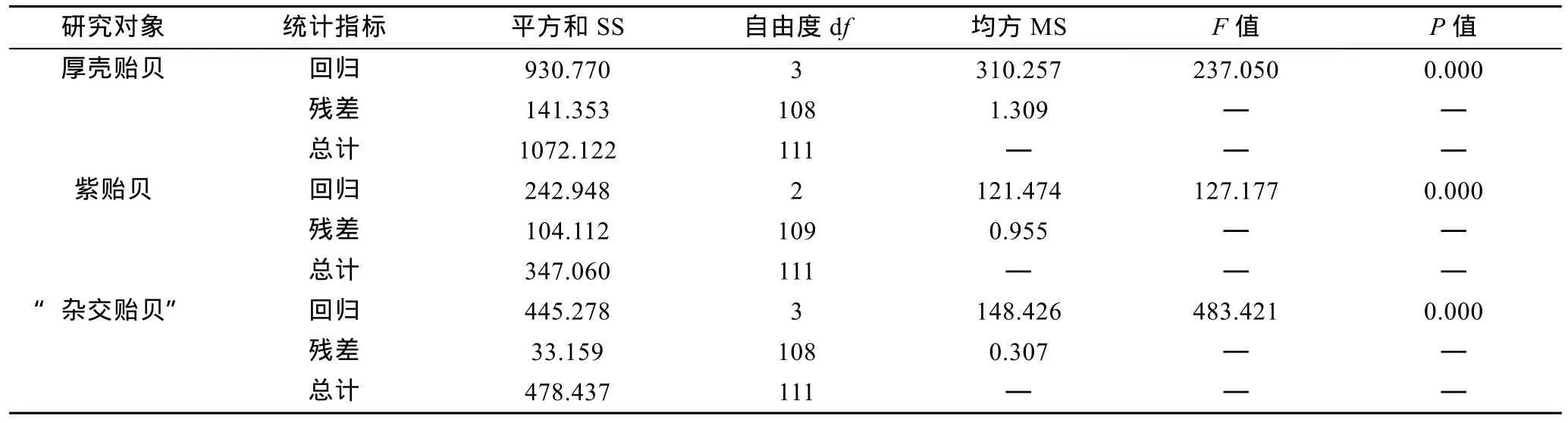
表9 实验贝贝体框架性状与壳重间多元回归方程的方差分析Tab.9 Analysis on the variance of the multiple regression equations of Mytilus
3 讨论
3.1 影响实验贝壳重关键贝体框架性状组合的确定
尽管本研究所涉 3种实验贝的各项贝体框架性状与其壳重的相关系数均达到极显著水平(表2、表3、表4), 但并非均为影响壳重的关键性状。通径分析结果显示, 对厚壳贻贝、紫贻贝和“杂交贻贝”测定群体壳重直接作用达到显著水平的性状组合分别仅为 OC、SH(BD)、SW, OC、SW 和 OC、SH(BD)、OA(表5), 复相关分析也显示上述被保留的贝体框架性状组合与其壳重均达到极显著水平(P<0.01), 而其余性状则均不显著(P>0.05) (表6), 表明通径分析较简单相关分析在度量壳重与贝体框架性状间实质性密切程度上更具可靠性。
刘小林等(2002)认为, 在表型相关分析的基础上,进行通径系数分析和决定系数分析时, 只有当相关指数R2或决定系数加和大于或等于 0.85时, 表明影响因变量的主要自变量已经找到。本研究中, 厚壳贻贝贝体框架性状OC、SH(BD)、SW组合和“杂交贻贝”贝体框架性状 SL、SH(BD)、OA组合对其壳重的决定系数加和分别为0.868和0.931(表6), 说明它们分别是决定厚壳贻贝和“杂交贻贝”壳重性状的关键贝体框架性状组合。紫贻贝被保留的贝体框架性状组合 OC、SW 虽对其壳重的决定系数加和仅为0.700(表6), 但从两者与壳重间均呈高度相关(表3),对壳重的直接作用均明显大于间接作用, 且两者交互对壳重决定具强协同效应(表5), 以及所建多元回归方程的可靠性(表8、表9)来看, 它们还是基本上能反映贝体框架性状与壳重间的真实关系, 故也均属决定紫贻贝壳重的关键性状。至于厚壳贻贝、紫贻贝以及“杂交贻贝”中, 与壳重呈高度相关的其它贝体框架性状未被选入的原因, 则可能如刘小林等(2004)所认为的其与入选的自变量相关性很强而不能在回归方程共存所致, 这一情形在日本沼虾(王志铮等,2011)、脊尾白虾(杨磊等, 2012)等水产养殖动物的相关研究中亦有出现。
3.2 实验贝贝体框架特征与其壳重增长对策的相关性
据报道, 贝壳的生物矿化(shell biomineralization)发生在位于外套膜和贝壳之间的外套膜外腔中(Weineret al, 1991; Belcheret al, 1996; Faliniet al, 1996), 外套膜外腔液是形成贝壳的物质库, 形成贝壳所需的有机质和Ca2+、CO32-等无机离子都由外套膜分泌, 集合于外套膜外腔中(张文兵等, 2008)。因此, 滤食性贝类的壳重与其外套膜外腔表面积有着极为密切的关系。本研究中决定厚壳贻贝、紫贻贝及“杂交贻贝”壳重性状的核心变量均仅为贝体长度性状 OC或 SL的结果(表5, 表6), 既与贻贝壳长与壳重的增长基本是一致的或略有前后 (王如才等, 1993)的观点相符,也与三角帆蚌(Hyriopsis cumingii)壳长对壳重的直接作用明显大于壳宽、壳高的结果(闻海波等, 2012)相吻合, 表明本研究同域养殖的3种实验贝与闻海波等(2012)报道的 3种不同地理居群三角帆蚌类似, 其外套膜外腔表面积的大小均主要取决于可表征其腔体长度性状的贝体长度性状指标值, 事实上本研究所涉 3种实验贝贝体长度性状均远大于其它贝体框架性状的结果(表1)也充分印证了上述判断的准确性。
外套腔系贝类外套膜与内脏团之间的空腔, 是物质进出的重要场所, 在体内外物质交换中起重要作用(蔡英亚等, 1979)。无疑, 贝类外套膜分泌形成贝壳所需的有机质和Ca2+、CO32–等无机离子势必依赖于流经腔内物质的持续供给。据报道, 嵊泗列岛紫贻贝的生长速度和性成熟速度均明显较厚壳贻贝快(张义浩等, 2003; 常抗美等, 2008), 正交F1代(厚壳贻贝♂×紫贻贝♀)生长性能指标与紫贻贝相当而显著高于厚壳贻贝(P<0.05)(常抗美等, 2008), 本研究所涉3种实验贝的滤食与消化功能区占比分别呈“杂交贻贝”> 紫贻贝 > 厚壳贻贝(P<0.05)和紫贻贝 >“杂交贻贝”≈厚壳贻贝,“杂交贻贝”通过显著提高滤食功能区占比来强化其杂种生长优势(白晓倩等, 2014)。因此, 厚壳贻贝SH(BD)、SW对壳重的直接作用均明显小于间接作用, 且两者对壳重的间接作用均主要通过OC来实现, 紫贻贝SW对壳重的直接作用大于间接作用, 以及“杂交贻贝”SH(BD)、OA对壳重的直接作用均明显小于间接作用, SH(BD)对壳重的间接作用主要通过SL来实现, 而SL和OA对壳重的间接作用则均主要通过 SH(BD)来实现等的结果(表5),无疑揭示了本研究所涉 3种实验贝中, 厚壳贻贝因滤食功能区水平剖面占比和消化功能区占比均较小, 生长最为缓慢, 故采取借助 SH(BD)性状扩展消化功能区水平剖面并增加外套膜外腔表面积, 通过 SW 性状扩容外套腔体增进滤食作用, 以此共同辅助 OC来促进其贝壳增重的壳重增长对策, 紫贻贝因滤食功能区水平剖面占比弱配于消化功能区占比, 故采取借助SW性状扩容外套腔体以增强滤食作用, 并与OC一起来促进其贝壳增重的壳重增长对策, “杂交贻贝”则因消化功能区占比严重弱配于滤食功能区占比, 致使其 SH(BD)性状在贝体消化功能区水平剖面的扩展上较厚壳贻贝更为困难, 尚需OA性状的配合。
综上分析可知, 决定本研究所涉3种实验贝壳重的核心变量均为贝体长度性状, 三者间贝体滤食功能区和消化功能区空间配置状况的差异是导致它们选择不同壳重生长对策的重要原因。
王如才, 王昭萍, 张建中, 1993. 海水贝类养殖学. 青岛: 青岛海洋大学出版社, 119—154
王志铮, 吴一挺, 杨磊等, 2011. 日本沼虾(Macrobrachium nipponensis)形态性状对体重的影响效应. 海洋与湖沼,42(4): 612—618
白晓倩, 杨阳, 邹李昶等, 2014. 嵊泗列岛海域三种贻贝贝体框架特征的差异. 海洋与湖沼, 45(5): 1078—1084
刘慧, 唐启升, 2011. 国际海洋生物碳汇研究进展. 中国水产科学, 18(3): 695—702
刘小林, 吴长功, 张志怀等, 2004. 凡纳滨对虾形态性状对体重的影响效果分析. 生态学报, 24(4): 857—862
刘小林, 常亚青, 相建海等, 2002. 栉孔扇贝壳尺寸性状对活体重的影响效果分析. 海洋与湖沼, 33(6): 673—678
杨磊, 赵晶, 杨鹏等, 2012. 池养脊尾白虾形态性状对体重的影响效应. 浙江海洋学院学报(自然科学版), 31(3):191—196
沈玉帮, 李家乐, 牟月军, 2006. 厚壳贻贝与贻贝遗传渗透的分子生物学鉴定. 海洋渔业, 28(3): 195—200
张义浩, 赵盛龙, 2003. 嵊山列岛贻贝养殖种类生长发育调查.浙江海洋学院学报(自然科学版), 22(1): 67—73
张文兵, 姚春凤, 麦康森, 2008. 贝壳生物矿化的研究进展.海洋科学, 32(2): 74—79
张继红, 方建光, 唐启升, 2005. 中国浅海贝藻养殖对海洋碳循环的贡献. 地球科学进展, 20(3): 359—365
张朝晖, 周骏, 吕吉斌等, 2007. 海洋生态系统服务的内涵与特点. 海洋环境科学, 26(3): 259—263
周毅, 杨红生, 刘石林等, 2002. 烟台四十里湾浅海养殖生物及附着生物的化学组成、有机净生产量及其生态效应.水产学报, 26(1): 21—27
闻海波, 顾若波, 曹哲明等, 2012. 3个地理种群三角帆蚌育珠相关性状比较及壳重的通径分析. 上海海洋大学学报,21(2): 161—166
唐启升, 2010. 发展碳汇渔业 抢占蓝色低碳经济的技术高地.科学时报, 2010-07-15
常抗美, 刘慧慧, 李家乐等, 2008. 紫贻贝和厚壳贻贝杂交及F1代杂交优势初探. 水产学报, 32(4): 552—557
董双林, 王芳, 王俊等, 1999. 海湾栉孔扇贝对海水浮游生物和水质的影响. 海洋学报, 21(6): 138—143
蔡英亚, 张英, 魏若飞, 1979. 贝类学概论(修订版). 上海:上海科学技术出版社, 28—214
Anderson E, Hubricht L, 1938. Hybridizatin in Tradescantia. The evidence for introgressive hybridization. Amer J Botany, 25:396—402
Belcher A M, Wu X H, Christensen R Jet al, 1996. Control of crystal phase switching and orientation by soluble molluscshell proteins. Nature, 381: 56—58
Falini G, Albeck S, Weiner Set al, 1996. Control of aragonite or calcite polymorphism by mollusk shell macromolecules.Science, 271: 67—69
Kaspar H F, Gillespie P A, Boyer I Cet al, 1985. Effects of mussel aquaculture on the nitrogen cycle and benthic communities in Kenepru Sounds, New Zealand. Marine Biology, 85: 127—136
Nakamura Y, Kerciku F, 2000. Effects of filter-feeding bivalves on the distribution of water quality and nutrient cycling in aeutrophic coastal lagoon. Journal of Marine Systems, 26:209—221
Prins T C, Escaravage V, Smaal A Cet al, 1995. Functional and structural changes in the pelagic system induced by bivalve grazing in marine mesocosms. Water Science Technique,32(4): 183—185
Weiner S, Addadi L, 1991. Acidic macromolecules of mineralized tissues: the controllers of crystal formation. Trends in Biochemical Science, 16: 252—256
Young Ahn, 1993. Enhanced particle flux through the biodeposition by the Antarctic suspension-feeding bivalveLater nulaellipticain Marian Cove, King George Island.Journal of Experimental Marine Biology and Ecology,171: 75—90
