丙二醛引起草鱼肠道、肝胰脏谷胱甘肽/谷胱甘肽转移酶通路抗氧化应激
2015-03-14林秀秀元土蔡春芳雨薇科全徐登辉其刚
林秀秀 叶 元土 蔡春芳 吴 萍 黄 雨薇 陈 科全徐登辉 彭 侃 罗 其刚
(苏州大学基础医学与生物科学学院,苏州 215123)
饲料油脂是水产动物所必需的结构性物质和能量物质[1]。然而,其易发生氧化酸败产生多种有害中间产物,会对鱼体肠道[2]、肝胰脏[3-4]等内脏器官产生重要影响。其中丙二醛(MDA)是脂质过氧化次生产物中含量最为丰富的活性醛[5],研究表明,MDA能与多种氨基酸或多肽作用,从而影响其蛋白质的结构功能[6],且MDA对线粒体呼吸功能及相关脱氢酶具有不同程度的损伤作用[7],进而导致细胞损伤。
还原型谷胱甘肽(GSH)是由谷氨酸、半胱氨酸和甘氨酸组成的小分子三肽,其能将体内有害物质转变为无害物质,并排出体外起到解毒作用,是体内重要的抗氧化剂和自由基清除剂[8]。研究证实,细胞内GSH的含量主要是通过谷胱甘肽还原酶(GSR)、谷胱甘肽合成酶、谷氨酸半胱氨酸连接酶和部分谷胱甘肽硫转移酶(GSTs)来调节的[9],同时这些酶构成了 GSH/GSTs通路。有研究表明,当机体受到有害物质侵害时,会引起GSH/GSTs通路相关酶发挥抗氧化防御作用[10-12],以保护细胞免受氧化损伤。
本试验在饲料添加不同梯度MDA的条件下,采用荧光定量PCR(qRT-PCR)技术对草鱼(Ctenopharyngodon idellus)肠道、肝胰脏的GSH/GSTs通路相关基因表达量进行测定,并结合肠道、肝胰脏和血清中MDA和GSH含量变化进行综合分析,旨在探讨MDA对草鱼肠道、肝胰脏GSH/GSTs通路的影响。
1 材料与方法
1.1 试验饲料
1.1.1 MDA 溶液的制备
制备原料为 1,1,3,3-四乙氧基丙烷(1,1,3,3-tetraethoxypropane)(Sigma-Aldrich公司产品,浓度≥96%)。制备方法:精确量取 1,1,3,3-四乙氧基丙烷 31.500 m L,用 95%乙醇溶解后定容至100 m L,搅拌 15 m in,此时每毫升溶液相当于MDA 100 mg。MDA现配现用,不做保存。
1.1.2 饲料配制
以酪蛋白和秘鲁蒸汽鱼粉为主要蛋白质源,豆油(“福临门”牌1级大豆油)为主要脂肪源,配制基础饲料。以基础饲料为对照组,然后在基础饲料中分别添加 61(B1组)、124(B2组)和185 mg/kg(B3组)的MDA,配制3种试验饲料。基础饲料组成及营养水平见表1。MDA的添加方式:依据每日的投喂量配制相应体积的MDA溶液,均匀的喷洒在饲料中。饲料中MDA含量依据本实验室饲料中氧化鱼油对草鱼生长及肌肉脂肪酸组成的影响[13]试验中氧化鱼油添加量对应的MDA含量设定。
1.2 试验鱼与饲养管理
草鱼来源于浙江一星饲料有限公司养殖基地,为池塘培育的1冬龄鱼种,共300尾,平均体重为(74.8±1.0)g。从中选择 240 尾草鱼,随机分为4组,每组设3个重复,每个重复20尾。养殖试验在浙江一星饲料集团试验基地进行,在面积为3 335 m2(平均水深1.8 m)的池塘中设置网箱,网箱规格为 1.0 m×1.5 m×2.0 m。将各组草鱼以重复为单位随机分配在12个网箱中。

表1 基础饲粮组成及营养水平(风干基础)Table 1 Composition and nutrient levels of the basal diet(air-dry basis)
各组草鱼分别用商品料驯化1周后,开始正式投喂。每天08:00和15:00定时投喂,投饲率为体重的2%~4%。每10 d依据投饲量估算鱼体增重并调整投喂率,记录每天的投饲量。正式养殖试验共72 d。
每周用试剂盒测定水质1次,试验期间溶解氧浓度>8.0 mg/L,pH 7.08~ 8.40,氨氮浓度<0.2 mg/L,亚硝酸盐浓度<0.01 mg/L,硫化物浓度<0.05 mg/L。试验期间养殖水温25~33℃。
1.3 样本采集与分析
1.3.1 基因样本采集与分析
1.3.1.1 基因样本采集
养殖72 d、停食24 h后,每网箱随机取3尾(每组共9尾)鱼作为基因分析样本。鱼体表面经75%酒精消毒,常规解剖,快速取出内脏团,在冰浴中剪取中肠的前1/4肠段以及部分肝胰脏,用磷酸盐缓冲溶液(PBS)漂洗2~3次,各样品一式二份,迅速装于EP管中,液氮速冻,于-80℃保存备用。采样所用剪刀镊子均经灭酶灭菌处理。
1.3.1.2 引物设计
基因序列依据叶元土等[14]基因测序结果,qRT-PCR中谷氨酸-半胱氨酸连接酶催化亚基(GCLC)、谷胱甘肽还原酶(GSR)、pi-谷胱甘肽硫转移酶(GSTpi)、微粒体谷胱甘肽硫转移酶1(MGSt1)基因及内参基因β-肌动蛋白(β-actin)所使用的 Taqman引物由 Prime 5.0软件设计,引物序列见表2。

表2 定量PCR引物Table 2 Primers used in quantitative PCR
1.3.1.3 总RNA 的提取和反转录cDNA
利用1.3.1.1所采集的单个网箱3尾草鱼肠道与肝胰脏样本,各取25 mg合并为1个样本用于总RNA的提取,每组3个平行样本(共9尾鱼、3个测定样本)。在样本中加入液氮,研磨成粉末后,加入1 m L Trizol覆盖粉末,待Trizol解冻成液体后全部转移到1.5 m L的EP管中,按照Trizol方法提取总RNA。按照PrimeScriptTMRT Master M ix(TaKaRa公司)反转录试剂盒的方法将RNA反转录成cDNA,将反转录产物稀释后于-20℃保存备用。
1.3.1.4 定量分析
使用CFX96荧光定量PCR仪和SYBR Premix Ex TaqTMⅡ(TaKaRa公司)荧光染料对草鱼GSH/GSTs通路的相关基因(GCLC、GSR、GST-pi、MGST1)及内参基因β-actin进行定量分析。反应体系为20μL:SYBR Prem ix Ex TaqTMⅡ10 μL,上、下游引物各 1 μL,cDNA 2 μL,灭菌水6μL。PCR反应采用2步法,反应条件为:95℃预变性30 s、95℃变性 5 s、60 ℃退火 30 s,共 40个循环,最后进行熔解曲线(melting curve)分析。
1.3.2 肠道、肝胰脏组织匀浆样品制备与分析
每个网箱随机另取3尾鱼,取部分肠段及肝胰脏,组织匀浆后,2 000 r/min离心15 m in,取上清液,用于GSH和MDA含量测定。GSH和MDA含量均采用南京建成生物工程研究所生产的试剂盒测定。
1.3.3 血清样品制备与分析
每个网箱随机另取10尾鱼,以无菌1 m L注射器自尾柄静脉采血,置于Eppendorf离心管中,室温自然凝固 0.5 h后 3 000 r/m in冷冻离心10 min,取上清液分装后,液氮速冻并于-80℃冰箱中保存。血清GSH和MDA含量均采用南京建成生物工程研究所生产的试剂盒测定。
1.4 数据处理
以β-actin为内参基因,计算各目的基因的相对表达量,计算公式为:

采用SPSS 18.0统计分析软件对数据进行分析,组间差异显著性用单因素方差分析(one-way ANOVA)进行统计分析,结果以平均值±标准误(mean±SD)表示。
2 结 果
2.1 肠道、肝胰脏、血清MDA含量
由表3可知,随着饲料中MDA含量的增加,肠道MDA含量有上升的趋势,且二者呈显著的正相关(P<0.05);然而,与对照组相比,各试验组肠道MDA含量均无显著变化(P>0.05)。与对照组相比,除B3组肝胰脏MDA含量显著升高(P<0.05)外,其余试验组均无显著变化(P>0.05),但肝胰脏MDA含量与饲料MDA含量存在显著的正相关(P<0.05)。血清中MDA含量均显著高于对照组(P<0.05)。

表3 肠道、肝胰脏和血清MDA含量及其与饲料MDA含量的相关性分析Table 3 The content of MDA in intestine,hepatopancreas and serum and its correlation analysis w ith dietary MDA content
2.2 肠道、肝胰脏、血清GSH含量
由表4可知,与对照组相比,肠道GSH含量除B2组外其余试验组均显著升高(P<0.05)。肝胰脏GSH含量各组间均无显著差异(P>0.05),但随饲料MDA含量升高有先上升后下降的趋势。血清GSH含量随饲料MDA含量升高呈下降趋势,但各试验组均高于对照组,其中B1和B2组与对照组具有显著差异(P<0.05)。
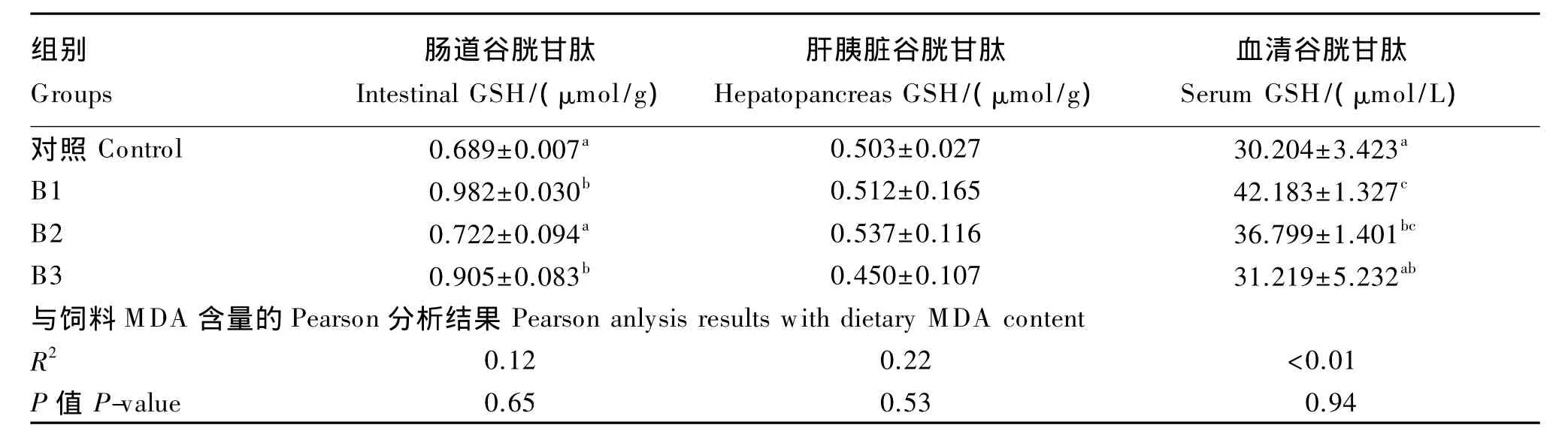
表4 肠道、肝胰脏和血清GSH含量及其与饲料MDA含量的相关性分析Table 4 The content of GSH in intestine,hepatopancreas and serum and its correlation analysis w ith dietary MDA content
2.3 肠道、肝胰脏GSH合成相关基因表达量
采用qRT-PCR技术检测了草鱼肠道、肝胰脏中GSH合成相关基因的表达量,分别包括GSH从头合成途径的关键基因GCLC以及参与还原途径的GSR,结果如表5所示。
随着饲料中MDA含量的增加,肠道GCLC表达量先上调后下调,且二者呈显著正相关(P<0.05);相比对照组,B2和B3组肠道 GCLC表达量均显著上调(P<0.05)。随着饲料中MDA含量的增加,肝胰脏GCLC表达量随之下调,其中B3组显著低于 B1 组(P<0.05)。
除B2组肠道GSR表达量显著上调(P<0.05)外,其余各组肠道和肝胰脏GSR表达量均无显著差异(P>0.05),且GSR的表达量与饲料MDA含量的相关性较低。
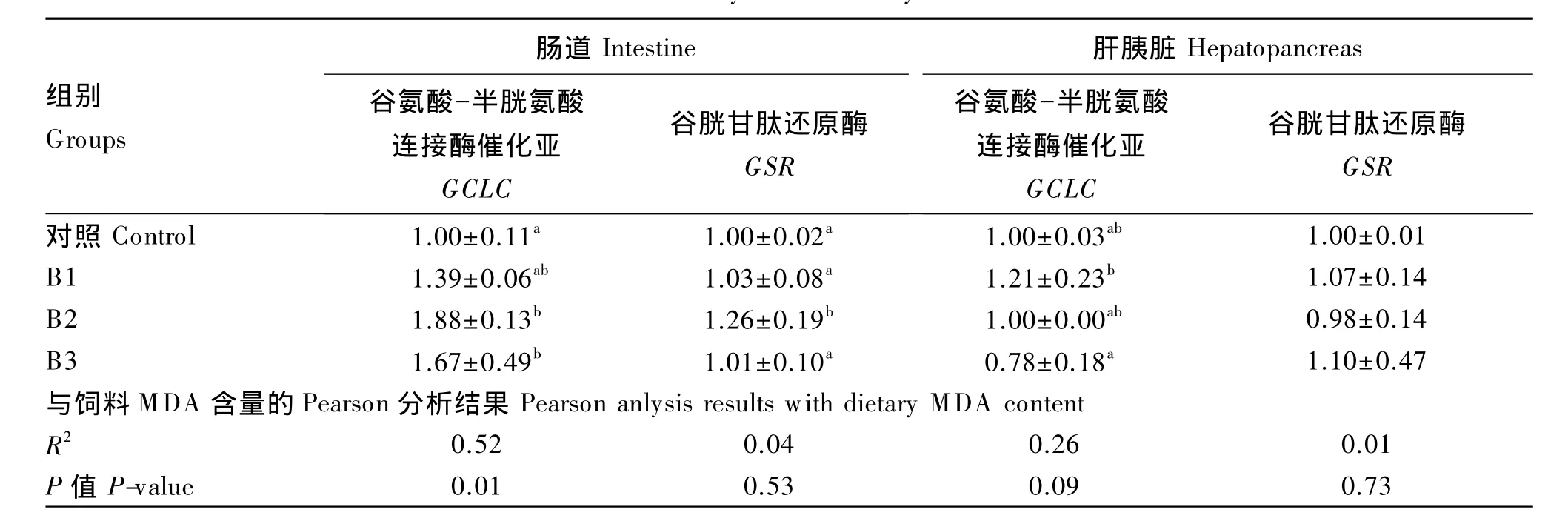
表5 草鱼肠道、肝胰脏中GCLC和GSR的表达量及其与饲料MDA含量的相关性分析Table 5 The expression levels of GCLC and GSR in intestine and hepatopancreas and their correlation analysis w ith dietary MDA content
2.4 肠道、肝胰脏GSTs基因表达量
采用qRT-PCR计算检测了肠道、肝胰脏中GSTs基因表达量,分别包括胞液酶GSTpi以及膜结合酶MGST1,结果如表6所示。
相比对照组,B3和B2组肠道GSTpi和B3组肠道MGST1的表达量显著上调(P<0.05),且肠道GSTpi与MGST 1表达量与饲料MDA含量均呈显著的正相关(P<0.05)。
随着饲料中MDA含量的增加,肝胰脏GSTpi表达量随之上调,且二者呈显著的正相关(P<0.05),其中 B3组相比对照组显著上调(P<0.05)。与对照组相比,肝胰脏MGST1表达量均显著下调(P<0.05)。

表6 草鱼肠道、肝胰脏GSTpi和MGST1表达量及其与饲料MDA含量的相关性分析Table 6 The expression levels of GSTpi and MGST1 in intestine and hepatopancreas and their correlation analysis w ith dietary MDA content
3 讨 论
MDA通过血液系统进入到体内后,可能直接发挥作用,也可能通过促使细胞膜脂质过氧化而导致细胞凋亡[15],因此其对细胞抗氧化系统有重大影响,GSH/GSTs通路是细胞抗氧化系统的重要组成,对维护细胞免受氧化损伤具有重要意义。
3.1 MDA对草鱼肠道、肝胰脏GSH合成相关基因表达的影响
研究表明,肝胰脏与肠道具有相同的胚胎起源,所以在一定程度上保持着解剖和功能性联系,肝胰脏血液供应主要由门静脉和肝动脉提供,其中门静脉的供血量占其全部供血的70%左右,且血液多数来自肠系膜上静脉及肠系膜下静脉,在这些静脉血管中通常含有来自消化道产物[16]。本试验中,在饲料添加不同剂量的MDA,对草鱼进行为期72 d的投喂,通过检测肝胰脏、肠道、血清中MDA含量,发现肠道MDA含量与饲料MDA含量呈正相关性,随饲料MDA含量的增加而增加,但其增加的幅度较小,相比对照组均无显著变化;而各试验组血清MDA含量相比对照组均显著升高。有研究表明,MDA化学稳定性和膜透过性均大于活性氧(ROS)[17],且MDA的代谢途径是由血液流向肝胰脏再由肝胰脏流向机体组织[18],肝胰脏是MDA的主要代谢器官组织,饲料中的MDA大部分经血液流向肝胰脏,因此肠道MDA含量变化较小而血液MDA含量变化较大。
γ-谷氨酰半胱氨酸合成酶(γ-GCL)是合成GSH的限速酶,由GCLC和谷氨酸半胱氨酸连接酶调节亚基(GCLM)组成,GCLC含有γ-GCL底物结合位点和催化功能。试验发现,经MDA刺激,肠道GCLC表达量相比对照组显著上调,且随饲料MDA含量的增加,GCLC表达量先上调后下调,且二者具有显著相关性;然而,肝胰脏GCLC表达量随饲料MDA含量的增加出现先上调后下调的趋势。至少80%的MDA是与蛋白质结合[19],形成的 MDA-蛋白质加合物,导致功能蛋白质功能发生紊乱,质膜流动性降低[20],且该加合物会导致蛋白质进一步氧化应激[21]。肝胰脏和肠道均受到MDA刺激的影响,而肝胰脏是MDA主要的代谢场所,其GCLC受到的刺激相比其他组织更加严重,因此表现出在MDA含量较低的B1组肝胰脏GCLC的表达量显著上调而B2与B3组其表达量下调,而肠道从外界摄入的MDA相比于肝胰脏中的要少,肠道GCLC表达量均表现出上调的趋势,因此少量MDA刺激肠道会使肠道GCLC表达量上调,以应对氧化应激。B3组肝胰脏MDA含量出现显著增加的现象,这可能是肝胰脏GCLC表达量不断下调,对鱼体肝胰脏的保护作用减弱,致使肝胰脏受到了实质性的损伤,从而导致MDA代谢作用减弱并在草鱼肝胰脏中积累。肝胰脏是合成GSH的主要器官组织,且所合成的GSH大部分进入血液后运输到各个器官组织当中。本试验结果发现,随饲料MDA含量的增加,肝胰脏中GSH含量无显著变化,然而血清中GSH含量出现先增加后下降的趋势,这与GCLC在肝胰脏中的表达量变化一致。
GSR是体内一种重要的抗氧化酶类,其主要的生理功能是利用还原型辅酶Ⅱ(NADPH)将氧化型谷胱甘肽(GSSG)还原成GSH,从而为活性氧ROS的清除提供还原力,保护机体免受伤害[22]。本试验发现,除B2组肠道GSR表达量显著升高外,各试验组的肠道和肝胰脏GSR表达量较对照组均无显著差异,且与饲料MDA含量的相关性较低,这可能是由于MDA的作用方式是与蛋白质、DNA等物质反应破坏其结构并生成有害物质而并非直接导致体内自由基的增加,因此GSH与自由基反应形成GSSG的量较少,肠道和肝胰脏GSR的表达量并未受到显著的影响。
3.2 MDA对草鱼肠道、肝胰脏GSTs基因表达的影响
GSTs 是一个具有多种生理功能的基因家族,是由2个分子质量为22~27 ku的多肽亚基组成的可溶性蛋白质,其主要以同源二聚体的形式存在,少数以异源二聚体的形式存在[23]。GSTs催化GSH与亲电子物质发生轭合反应的实质是降低GSH的酸度系数,一旦在活性中心形成硫醇基团,它就能与距硫醇离子很近的H位点的亲电子底物发生反应[24],除此之外,它还具过氧化物酶、脱氯化氢酶等的活性[25]。由于MDA能与多种氨基酸或多肽作用,这些作用可能影响蛋白质的结构功能或者破坏其空间构象,且MDA形成的加合物会对蛋白质等造成二次损伤,在这个过程中会产生多种有害代谢产物,从而刺激GSTs在保护细胞免受细胞毒素和致癌因子的损害发挥重要作用。
本试验涉及GSTs的2种同工酶,分别是胞液酶GSTpi和膜结合酶 M GST1。GSTpi是 G STs家族重要成员,由一个活跃的多态的基因编码,可清除有害物质,避免基因组的损伤[26],也参加及调节细胞的增殖、凋亡、炎症及分化反应[27-28],为此,GSTpi被认为对细胞具有保护作用。研究表明,在正常生长条件下,GSTpi以单体形式与c-Jun氨基末端激酶(JNK)形成复合物,抑制JNK活性,受到有害物质如MDA等刺激后,GSTpi自身形成二聚体,致使复合物解离,JNK磷酸化激活转录因子c-Jun,促进GSTpi转录,增加其含量,其又能反馈抑制JNK,维持 JNK的低活性[29],从而大大降低细胞因受外界刺激而造成的损伤。MDA能与细胞中蛋白质、氨基酸、DNA形成相应的加合物,从而破坏细胞结构,且MDA还能与信号蛋白作用,在肝星状细胞(HSCs)中诱导尿激酶型纤溶酶原激活剂分泌增加,从而促进肝纤维化的进展[30]。本试验发现,经MDA刺激后,GSTpi的表达量在肝胰脏和肠道均有所上调,且在B3组均达到最大值。这可能是由于MDA导致细胞损伤,使GSTpi表达量上调,加快清除体内有害物质且在一定程度上抑制了细胞的凋亡。由于MDA主要的代谢部位是肝胰脏,对肝胰脏的损伤作用相对于肠道会更严重,因此本研究发现在受到MDA刺激后,肝胰脏GSTpi的表达量高于肠道。
MGST1是一个分子质量大约为17 ku的膜结合 GSTs[31]。相比胞液 GSTs,MGST1 具有 1 个特殊的Cys49结构,有害物质能够修饰Cys49基团,这些修饰包括烷基化、形成二硫键(或者混合二硫化物与GSH或蛋白质二聚体),或次磺酸修饰,诱导其构象变化和增加活性[32-33]。本研究发现,MDA刺激下,肠道中MGST1表达量上调,而肝胰脏中MGST1表达量则下调。这可能是由于MDA与细胞膜作用,促使其脂质过氧化,产生各种有害物质刺激肠道细胞MGST1表达量上调,MGST1活性增加以加强催化GSH与有害物质结合转变为无害物质并排出体外,起到保护肠道细胞免受进一步损伤的作用。MGST1主要存在于肝胰脏线粒体的内外膜上,而在其他组织中含量较少[34],过量的MGST1会导致肝细胞中线粒体通透性转换孔(MPT)的形成,有害物质进入线粒体当中,使线粒体功能障碍,导致细胞凋亡和坏死[35],因此,在肝细胞中适当减少MGST1的表达也是对细胞的一种保护作用。
将肠道与肝胰脏中2种同工酶与饲料MDA含量进行相关性分析,结果发现除肝胰脏MGST1的表达量与饲料MDA含量相关性较低外,其余均显著相关,这进一步证明了上述的解释,即草鱼肝胰脏MGST1在受到MDA刺激后,表现出两面性:一方面,MGST1催化GSH与有害物质反应起到解毒的作用;另一方面,过多的MGST1会对肝细胞的线粒体造成损伤,其整体表现出的表达量与饲料MDA含量显示较低的相关性,同时,因GSTpi的性质,使得肠道、肝胰脏GSTpi的表达量与饲料MDA含量呈显著正相关;而MGST1在肠道的表达量比肝胰脏少,还未表现出过多的MGST1所造成损伤作用的一面,因此也与饲料MDA含量呈显著正相关。
4 结论
饲喂不同含量MDA的饲料72 d后,MDA诱导肠道GSH/GSTs通路相关基因表达量上调,且肠道GSH含量也有所上升,表明MDA引起了草鱼肠道的抗氧化应激。饲喂低含量MDA(61 mg/kg)饲料后草鱼肝胰脏GCLC表达量表现为上调,而MDA含量超过124 mg/kg时则表现为下调,这表明MDA引起了草鱼肝胰脏的抗氧化应激,并随MDA含量升高其抗氧化应激能力出现下降的趋势。草鱼肠道和肝胰脏中MGST1表达量出现了完全不同的趋势,这表明草鱼肠道和肝胰脏受MDA的影响程度有一定的差异。
[1] 李勇,王雷,蒋克勇,等.水产动物营养的生态适宜与环保饲料[J].海洋科学,2004,28(3):76-78.
[2] 汪开毓,叶仕根,耿毅.氧化脂肪对鱼类危害的病理及防治[J].淡水渔业,2002,32(4):60-63.
[3] 韩雨哲,姜志强,任同军,等.氧化鱼油与棕榈油对花鲈肝脏抗氧化酶及组织结构的影响[J].中国水产科学,2010,17(4):798-806.
[4] 任泽林,曾虹,霍启光,等.氧化鱼油对鲤肝胰脏抗氧化机能及其组织结构的影响[J].大连水产学院学报,2000,15(4):235-243.
[5] KROHNE T U,KAEMMERER E,HOLZ F G,et al.Lipid peroxidation products reduce lysosomal protease activities in human retinal pigment epithelial cells via two differentmechanisms of action[J].Experimental Eye Research,2010,90(2):261-266.
[6] ISHIIT,KUMAZAWA S,SAKURAI T,et al.Mass spectroscopic characterization of protein modification by malondialdehyde[J].Chem ical Research in Toxicology,2006,19(1):122-129.
[7] 龙建纲,王学敏,高宏翔,等.丙二醛对大鼠肝线粒体呼吸功能及相关脱氢酶活性影响[J].第二军医大学学报,2006,26(10):1131-1135.
[8] 童海达,王佳茗,宋英.Keap1-Nrf2-ARE在机体氧化应激损伤中的防御作用[J].癌变·畸变·突变,2013,25(1):71-75.
[9] BANKIK,HUTTER E,COLOMBO E,et al.Glutathione levels and sensitivity to apoptosis are regulated by changes in transaldolase expression[J].Journal of Biological Chem istry,1996,271(51):32994-33001.
[10] 聂芳红,孔庆波,刘连平,等.两种二 英类化合物对斑马鱼肝脏MDA、SOD和GST的影响[J].食品与生物技术学报,2009,28(2):210-213.
[11] 郑英,楼宜嘉.对乙酰氨基酚致小鼠肝微粒体谷胱甘肽S-转移酶的激活机制[J].中国药理学与毒理学杂志,2003,17(3):211-215.
[12] 姚芹.亚砷酸钠对小鼠体内谷胱甘肽S-转移酶活力及其基因表达的影响[D].硕士学位论文.乌鲁木齐:新疆医科大学,2009.
[13] 陈科全,叶元土,蔡春芳,等.饲料中氧化鱼油对草鱼(Ctenopharyngodon idellus)生长及肌肉脂肪酸组成的影响[J].动物营养学报,2015,27(6):1698-1708.
[14] 叶元土,蔡春芳,吴萍.氧化油脂对草鱼生长和健康的损伤作用[M].北京:中国农业科技出版社,2015.
[15] 姚仕彬,叶元土,蔡春芳,等.丙二醛对离体草鱼肠道黏膜细胞的损伤作用[J].水生生物学报,2015,39(1):137-146.
[16] 陈蛟,张映林,刘作金.肝肠轴相关机制研究进展[J].现代医药卫生,2014,30(22):3405-3408.
[17] ESTERBAUER H,SCHAUR R J,ZOLLNER H.Chem istry and biochem istry of 4-hydroxynonenal,malonaldehyde and related aldehydes[J].Free Radical Biology and Medicine,1991,11(1):81-128.
[18] 李莉,陈菁菁,李方序,等.氧应激毒性产物丙二醛(MDA)对小鼠体能的影响及其体内代谢[J].湖南师范大学自然科学学报,2006,29(2):97-101.
[19] SLATTER D A,BOLTON C H,BAILEY A J.The importance of lipid-derived malondialdehyde in diabetesmellitus[J].Diabetologia,2000,43(5):550-557.
[20] UCHIDA K,SAKAI K,ITAKURA K,et al.Protein modification by lipid peroxidation products:formation ofmalondialdehyde-derived Ne-(2-propenal)lysine in proteins[J].Archives of Biochem istry and Biophysics,1997,346(1):45-52.
[21] TRAVERSO N,MENINIS,MAINERIE P,et al.Malondialdehyde,a lipoperoxidation-derived aldehyde,can bring about secondary oxidative damage to proteins[J].The Journals of Gerontology Series A:Biological Sciences and Medical Sciences,2004,59(9):B890-B895.
[22] CHEN JJ,YU B P.A lterations inm itochondrialmembrane fluidity by lipid peroxidation products[J].Free Radical Biology and Medicine,1994,17(5):411-418.
[23] EDWARDS R,DIXON D P,WALBOT V.Plant glutathione S-transferases:enzymes w ith multiple functions in sickness and in health[J].Trends in Plant Science,2000,5(5):193-198.
[24] 裴冬丽.谷胱甘肽还原酶在植物防御中的研究进展[J].中国农学通报,2012,28(18):185-188.
[25] 张媛.产黄青霉谷胱甘肽转移酶基因的克隆、表达与功能研究[D].硕士学位论文.石家庄:河北师范大学,2007.
[26] TEW K D.G lutathione-associated enzymes in anticancer drug resistance[J].Cancer Research,1994,54(16):4313-4320.
[27] CHO SG,LEE Y H,PARK H S,et al.Glutathione S-transferase mu modulates the stress-activated signals by suppressing apoptosis signal-regulating kinase 1[J].Journal of Biological Chem istry,2001,276(16):12749-12755.
[28] RUSCOE J E,ROSARIO L A,WANG T L,et al.Pharmacologic or genetic manipulation of glutathione S-transferase P1-1(GSTπ)influences cell proliferation pathways[J].Journal of Pharmacology and Experimental Therapeutics,2001,298(1):339-345.
[29] WU Y,FAN Y,XUE B,et al.Human glutathione S-transferase P1-1 interacts w ith TRAF2 and regulates TRAF2-ASK1 signals[J].Oncogene,2006,25(42):5787-5800.
[30] AYALA A,MUÑOZM F,ARGÜELLESS.Lipid peroxidation:production,metabolism,and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal[J].Oxidative Medicine and Cellular Longevity,2014,2014(2014):1-31.
[31] LENGQVIST J,SVENSSON R,EVERGREN E,et al.Observation of an intact noncovalent homotrimer of detergent-solubilized rat microsomal glutathione transferase-1 by electrospray mass spectrometry[J].Journal of Biological Chemistry,2004,279(14):13311-13316.
[32] SHINNO E,SHIMOJIM,IMAIZUMI N,et al.Activation of rat livermicrosomal glutathione S-transferase by gallic acid[J].Life Sciences,2005,78(1):99-106.
[33] IMAIZUM IN,M IYAGIS,ANIYA Y.Reactive nitrogen species derived activation of rat liver m icrosomal glutathione S-transferase[J].Life Sciences,2006,78(26):2998-3006.
[34] MORGENSTERN R,LUNDQVIST G,ANDERSSON G,et al.The distribution of m icrosomal glutathione transferase among different organelles,different organs,and different organisms[J].Biochem ical Pharmacology,1984,33(22):3609-3614.
[35] SCHAFFERT C S.Role of MGST1 in reactive intermediate-induced injury[J].World Journal of Gastroenterology,2011,17(20):2552-2557.
