恶性入侵植物豚草的繁育系统特性
2015-03-13郝建华金洁洁陈国奇王立新
郝建华, 金洁洁, 陈国奇, 王立新
1 常熟理工学院生物与食品工程学院, 常熟 215500 2 广东省农业科学院植物保护研究所/广东省植物保护新技术重点实验室, 广州 510640
恶性入侵植物豚草的繁育系统特性
郝建华1,*, 金洁洁1, 陈国奇2, 王立新1
1 常熟理工学院生物与食品工程学院, 常熟 215500 2 广东省农业科学院植物保护研究所/广东省植物保护新技术重点实验室, 广州 510640
豚草(AmbrosiaartemisiifoliaL.)是原产于北美的一年生恶性入侵植物,目前已广泛分布在我国东北、华北、华中和华东等地,威胁农业、生态和民众健康。该种雌、雄头状花序同株,以种子繁殖,因而其有性繁殖特性对其扩散和爆发具有重要影响。运用套袋授粉和联苯胺-过氧化氢法等实验方法,对入侵江苏常熟的豚草种群的繁育系统特性进行了研究。结果表明,豚草花粉活力在开花后第4天开始出现,第8天到第10天花粉活力比较高;柱头可授性在开花第2天开始出现,第5天到第8天柱头可授性较高;同植株上同一时期开花的雌花的柱头可授性和雄花的花粉活力有5d左右的重叠期。不套袋处理(自然条件)和异株授粉处理下,豚草的结实率都比较高,分别达48.4%和44.4%,两者间无显著差异(P> 0.05);而同株授粉处理的结实率较低,仅3.4%,极显著低于不套袋处理和异株授粉的结实率(P<0.01)但显著大于0(P< 0.05)。分析表明,豚草属于自交不亲和种,但又可部分自交亲和,不具有无融合生殖特性。因此,对在新分布区的零星豚草植株进行及时防除,将取得事半功倍的效果。
豚草; 菊科; 繁育系统; 外来入侵植物; 自交不亲和性
豚草(Ambrosiaartemisiifolia)是原产于北美的一年生恶性杂草,该种在我国21个省、市、自治区的1038个县分布,发生面积达0.0247 亿hm2,在入侵地形成优势群落,急剧降低生物多样性,导致的生态损失无法估算[1]。该种在欧洲、亚洲和大洋洲广泛入侵,并对农田造成了严重危害[2]。Fumanal 等[3]对法国豚草种群进行了野外调查,结果表明该种的生态适应幅极其广泛,能在不同土壤类型、植被群落结构组成和类型的生境下大量爆发,并且该种对春季作物和各种半自然生境具有潜在的威胁。此外,豚草极易产生除草剂的抗性,除草剂抗性治理委员会(Herbicide Resistance Action Committee,HRAC)数据库纪录的抗除草剂豚草种群有30个,其明显具有抗性的除草剂活性成份涉及30多种[4]。豚草以有性生殖产生的种子繁殖,据文献报道,豚草的种子产量巨大,一株较小的豚草植株即能产生超过30000枚种子,而高大的植株甚至能产生62000枚种子[5]。豚草种子具有明显的休眠性,部分种子在土壤中埋藏39a仍能萌发[5]。
豚草的繁育系统等有性繁殖特性与其种子的产量和质量密切相关,对其能否在新侵入地区成功定植和快速广泛扩散具有重要影响。前人的多项研究曾报道豚草能通过自交和异交产生种子[5-8]。然而Friedman 和Barrett[2]利用分子生物学方法和花粉-柱头亲和性反应对豚草原产地种群的研究表明,豚草自交不亲和。因此,还需要更多的研究实例,明确豚草的繁育系统特性及其与入侵性之间的关系。
本研究通过同质园栽培和观察实验,对豚草入侵地种群(江苏常熟)的花粉活力、柱头可授性、自交亲和性等繁育系统特性进行了研究,为阐明其繁育系统特性与其入侵性之间的关系提供基础资料,并为深入了解豚草在入侵地的扩散规律和趋势及制订科学的防控措施提供科学依据。
1 实验方法
1.1 实验材料
豚草为菊科豚草属植物,其花序是菊科植物中高度特化的类型,传粉类型已由菊科植物典型的虫媒传粉转化成风媒传粉[9-10]。它们的雄花和雌花分别组成雄头状花序和雌头状花序。雄头状花序排列在茎和小枝末端,由10—100(-200)朵雄花组成,呈葇荑花序状。当花粉成熟时释放大量风媒花粉。雌头状花序发生在叶腋内,无柄,聚成簇状。每一雌头状花序中仅包含一朵雌花,成熟后仅能产生一粒瘦果[5, 9]。
本研究的豚草种群为采自江苏常熟的野外植株。2009年5月从常熟虞山移栽了40株豚草幼苗至常熟理工学院东湖校区,置于花盆中进行露天种植。除土壤过分干燥时浇水外,未进行其它人工管理干预,以尽量模拟其野外生活条件。
1.2 花粉活力的动态
在豚草开花盛期的2009年7月21日—8月5日期间,采用联苯胺法测定雄花序的花粉活力[11]。具体步骤为:在10棵植株的各株上随机标记36个花药开放前一天的雄花序(以总苞片张开作为开花的依据);于开花后1—12d中每天10:00—11:00从每株各取3个花序,分别放入加有试剂Ⅰ(0.5%联苯胺、0.5%甲萘酚和0.25%碳酸钠各取10mL混合均匀)1mL和一滴0.3%过氧化氢溶液的离心管中带回实验室;将整个花序放在洁净的比色板上,用剪刀剪下花药及其相连部分放回离心管中,用玻璃棒捣碎花药,用旋涡混合器旋转1—2 min;把旋转好的溶液倒入比色板中,用吸管吸取混合液,分别滴在3个载玻片上;载玻片放在铺有湿滤纸的培养皿中,皿外用遮光纸袋糊住,放在35 °C的培养箱中温育30 min后,在显微镜下观察;统计每个载玻片上的有活力花粉占总花粉数的比例,每样本统计的总花粉粒数约为100粒。
1.3 柱头可授性的动态
采用联苯胺-过氧化氢法测定豚草雌花的柱头可授性[12]。具体步骤为:在10棵植株的各株上随机标记36个开花前一天的雌花序(以总苞片张开作为开花的依据;开花后每天10:00—11:00从每株上各取3个新鲜花序,将其浸入含有联苯胺-过氧化氢的离心管中带回实验室;将雌花放入含有反应液的比色板凹穴中,在解剖镜下统计柱头可授性。以呈现蓝色并有大量气泡出现的柱头作为有活力的柱头,而无活力的柱头不呈现蓝色且无或有少量气泡出现。
1.4 繁育系统的鉴定
于豚草开花盛期的7月16日—8月16日,在10个植株上随机选55朵雌花序,分别进行同株授粉、异株授粉、不授粉不套袋(自然条件)和套袋不授粉4种处理。同株授粉、异株授粉和套袋不授粉3种处理的雌花均在总苞片张开前进行套袋;同株授粉和异株授粉两种处理的授粉时间为开花后第4天,在授粉后再行套袋。当瘦果成熟后,统计有饱满种子的雌花序占所有雌花序的百分比。
此外,于授粉处理48 h后、从5个植株的4种处理中各取一朵雌花,立即放入FAA固定液中,带回实验室。利用常规的苯胺蓝荧光显微镜方法,用Nikon ECLIPSE 90i显微镜镜中的荧光装置,检测不同授粉条件下的花粉-柱头亲和性反应,并用连接在显微镜上的DXM1200C型CCD进行观察。
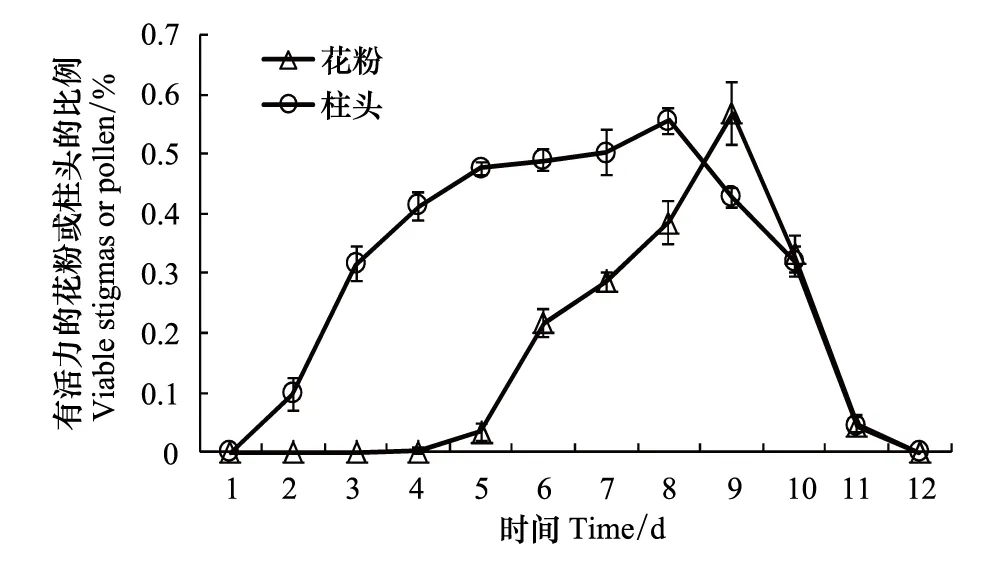
图1 豚草开花后有活力的花粉和柱头所占比例Fig.1 Percentage of viable pollen and stigmas of Ambrosia artemisiifolia after flowered
1.5 数据统计分析
采用SPSS13.0统计软件检验变量之间是否存在差异,并用Excel软件进行绘图。
2 结果
2.1 花粉活力和柱头可授性的动态
豚草开花第4天花粉开始有活力(图1);开花第8天和第10天花粉活力比较高;开花第9天花粉活力最高,有活力花粉数可达56.80%;之后花粉活力逐渐下降,到开花第12天花粉活力为0。开花第2天柱头就开始有活力(图1);开花第5天、第6天和第7天柱头可授性都比较高;开花第8天柱头可授性最高,有活力柱头可达55.56%;之后柱头可授性逐渐下降,到开花第12天柱头可授性为0。
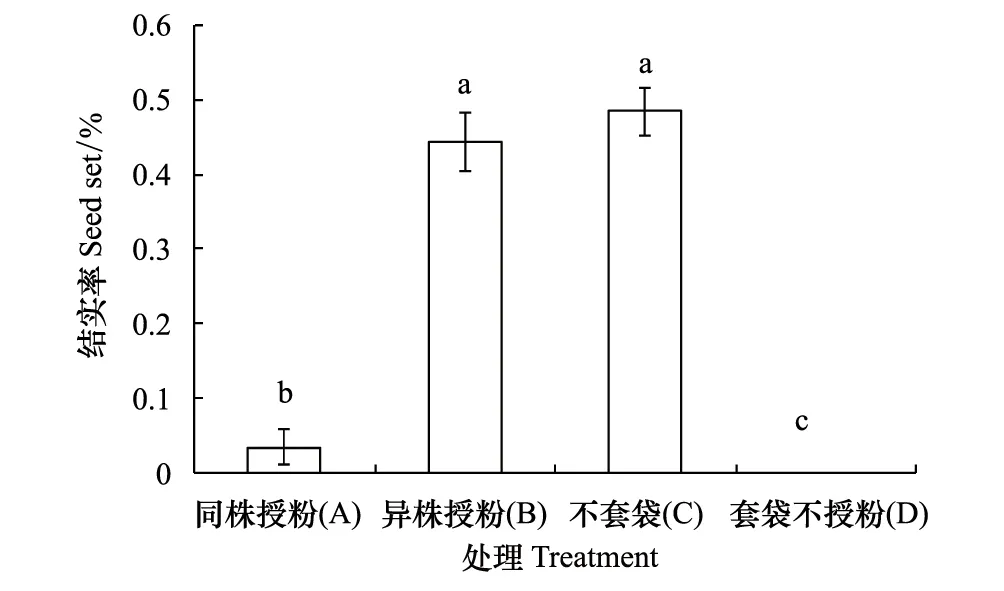
图2 豚草同株授粉(A)、异株授粉(B)、不套袋(C)和套袋不授粉(D)4种处理的结实率(平均值±标准差)Fig. 2 Mean (±SD) seed set of Ambrosia artemisiifolia for geitonogamy (A), xenogamous (B), open-pollinated (C) and bagged capitula (D). Significant differences (P < 0.05) in seed set between treatments are indicated by different letters above the bars for each species separately
2.2 繁育系统
同株授粉、异株授粉、不套袋和套袋不授粉4种处理的结实率的统计结果表明(图2),异株授粉处理和不套袋处理(自然条件)下,豚草都可以正常结实,且两者结实率都比较高, 分别达44.4%和48.4%;同株授粉处理的结实率很低,仅3.4%;套袋不授粉处理的结实率为0。对处理所得数据进行单因素方差分析,结果表明:异株授粉和不套袋处理(自然条件)之间无显著差异,且都极显著高于同株授粉(P<0.01);而同株授粉处理的结实率显著大于0(P< 0.05)。
同株授粉、异株授粉、不套袋和套袋不授粉4种处理的雌花柱头的荧光镜检结果表明,同株授粉处理的雌花柱头上有花粉粒,但不萌发;异株授粉处理的雌花柱头上有花粉粒,且萌发;不套袋处理的雌花柱头上有花粉粒,也萌发;但套袋不授粉处理的花序柱头上无花粉粒,不萌发。
3 结论与讨论
豚草具有极高的入侵性,在原产地以及欧洲,豚草在多种生境类型下泛滥成灾,是当地农田中危害最为严重的主要恶性杂草之一[3, 6, 11]。本文对其开花动态及繁育系统进行了实验研究,为深入了解其交配系统特性等繁殖特性与入侵性的关系提供了基础资料。
本文的研究结果和Friedman 与Barrett[2]的研究结果均表明,豚草的交配系统以自交不亲和为主,豚草的种子主要是通过不同植株之间的异交产生的。本研究的结果还表明,同株豚草的花粉活力和柱头可授性在第6天到第10天均在20%以上,有长达5d的自交结实的机会,但自交结实率仅有3.4%,不能进行无融合生殖。
Hao等[13]的研究表明,入侵我国的大多数一年生或越年生菊科外来植物明显倾向于自交亲和,并且菊科入侵植物在中国的分布范围与自交产生种子占全部种子的比例显著正相关。在已报道过繁育系统的菊科外来入侵植物中,一些多年生植物具有自交不亲和特性,如南美蟛蜞菊(Wedeliatrilobata)[14]、薇甘菊(Mikaniamicrantha)[15]、加拿大一枝黄花(Solidagocanadensis)[13, 16]和剑叶金鸡菊(Coreopsislanceolata)[13,17]等。这些自交不亲和的入侵种均既可通过地下茎进行营养繁殖, 又可通过有性生殖产生大量种子,两种繁殖方式的能力均很强。有性生殖主要负责这些入侵种的长距离扩散和种群的初期建群,而无性繁殖则更多地负责局域种群的生存壮大。
豚草作为一年生外来入侵植物,主要通过有性生殖繁殖后代,它与其他一年生或越年生菊科外来入侵植物常具有自交亲和性的繁育系统不同,通过自交不亲性保持了较高的异交率。这可能与其高度特化的头状花序类型、特殊的风媒传粉方式、巨额的花粉量、较大的种子量和较长的种子休眠期有关[2]。豚草为雌雄同株植物,它的雌花和雄花分别着生在不同的头状花序中,其花序结构与大多数菊科植物的头状花序的不同,这样高度特化的单性花头状花序类型与其它由两性花或雌花——两性花组成的头状花序类型相比,更有利于异交的进行。豚草的花和花序已由菊科植物典型的虫媒传粉演化为适于风媒传粉,其散粉过程既不同于其它菊科植物, 又有别于杨树等典型的风媒植物,是菊科植物中高度特化的传粉类型[10]。豚草所具有自交不亲和特性,使其交配方式以异交为主。异交有利于子代产生新的遗传变异,避免自交衰退,从长远效应,有利于豚草的种群发展壮大。但对于新入侵的豚草新植株和小种群而言,不利于其繁殖保障的短期效应。豚草在繁殖保障和避免自交衰退的权衡中,选择了后者作为生存策略,有利于其在入侵地种群的长期生存和壮大。
外来种在新的分布区入侵和扩散的首要环节便是成功定植和建群[18],而对于以种子繁殖的入侵植物而言,能否产生大量有活力的种子对外来种在新侵入地区成功定植和快速广泛扩散具有重要影响[19]。本文的研究结果表明,豚草主要通过异交产生大量有活力的种子,另一方面,豚草通过自交能产生少量种子(自交结实率达3.4%),因此,对在新分布区的零星豚草植株进行及时防除,将取得事半功倍的效果。
[1] 周忠实, 郭建英, 李保平, 孟玲, 傅建炜, 陈红松, 马明勇, 史梦竹, 李敏, 郭薇, 罗敏, 郑兴汶, 郑海燕, 罗源华, 万方浩. 豚草和空心莲子草分布与区域减灾策略. 生物安全学报, 2011, 20(4): 263-266.
[2] Friedman J, Barrett S C H. High outcrossing in the annual colonizing speciesAmbrosiaartemisiifolia(Asteraceae). Annals of Botany, 2008, 101(9): 1303-1309.
[3] Fumanal B, Girod C, Fried G, Bretagnolle F, Chauvel B. Can the large ecological amplitude ofAmbrosiaartemisiifoliaexplain its invasive success in France? Weed Research, 2008, 48(4): 349-359.
[4] Heap, I. The international survey of herbicide resistant weeds. Online. Internet. Tuesday, September 16, 2014. Available from: http://www.weedscience.org/Summary/Species.aspx?WeedID=18
[5] Bassett I J, Crompton C W. The biology of Canadian weeds: 11.AmbrosiaartemisiifoliaL. andA.psilostachyaDC. Canadian Journal of Plant Science, 1975, 55(2): 463-476.
[6] Genton B J, Shykoff J A, Giraud T. High genetic diversity in French invasive populations of common ragweed,Ambrosiaartemisiifolia, as a result of multiple sources of introduction. Molecular Ecology, 2005, 14(14): 4275-4285.
[7] McKone M J, Tonkyn D W. Intrapopulation gender variation in common ragweed (Asteraceae,AmbrosiaartemisiifoliaL.), a monoecious, annual herb. Oecologia, 1986, 70(1): 63-67.
[8] Lundholm J T, Aarssen L W. Neighbor effects on gender variation inAmbrosiaartemisiifolia. Canadian Journal of Botany, 1994, 72(6): 794-800.
[9] 关广清. 豚草花序及其演化. 沈阳农业大学学报, 1992, 23(4): 329-334.
[10] 关广清. 豚草花器结构及排粉过程. 植物检疫, 1993, 7(3): 167-170.
[11] King J R. The peroxidase reaction as an indicator of pollen viability. Stain Technology, 1960, 35(4): 225-227.
[12] Dafni A, Maués M M. A rapid and simple procedure to determine stigma receptivity. Sexual Plant Reproduction, 1998, 11(3): 177-180.
[13] Hao J H, Qiang S, Chrobock T, van Kleunen M, Liu Q Q. A test of baker′s law: breeding systems of invasive species of Asteraceae in China. Biological Invasions, 2011, 13(3): 571-580.
[14] 吴彦琼, 胡玉佳, 陈江宁. 外来植物南美蟛蜞菊的繁殖特性. 中山大学学报: 自然科学版, 2005, 44(6): 93-96.
[15] Hong L, Shen H, Ye W H, Cao H L, Wang Z M. Self-incompatibility inMikaniamicranthain South China. Weed Research, 2007, 47(4): 280-283.
[16] 郝建华, 钱海军, 姜雯, 沈宗根. 加拿大一枝黄花有性生殖特征研究. 生态环境学报, 2009, 18(6): 2278-2282.
[17] 曾建军, 肖宜安, 孙敏. 入侵植物剑叶金鸡菊的繁殖特征及其与入侵性之间的关系. 植物生态学报, 2010, 34(8): 966-972.
[18] Williamson M H, Fitter A. The characters of successful invaders. Biological Conservation, 1996, 78(1/2): 163-170.
[19] van Kleunen M, Manning J C, Pasqualetto V, Johnson S D. Phylogenetically independent associations between autonomous self-fertilization and plant invasiveness. American Naturalist, 2008, 171(2): 195-201.
Breeding system of a noxious invasive alien plant,AmbrosiaartemisiifoliaL.
HAO Jianhua1,*, JIN Jiejie1, CHEN Guoqi2, WANG Lixin1
1CollegeofBiologyandFoodEngineering,ChangshuInstituteofTechnology,Changshu215500,China2GuangdongProvincialKeyLaboratoryofHighTechnologyforPlantProtection,PlantProtectionResearchInstitute,GuangdongAcademyofAgriculturalSciences,Guangzhou510640,China
Ragweed (AmbrosiaartemisiifoliaL.), a member of the Asteraceae (Compositae), is an annual species native to North America. The species is a noxious invasive plant worldwide, with infestation by ragweed frequently posing challenges to protection and functioning of local ecosystems, crop production in agricultural areas, and public health. In North America, Europe, and Australia, ragweed is a major nuisance in arable areas and other habitats. At present, ragweed is relatively rare in agricultural areas of China compared with corresponding regions of other countries; nevertheless, the species is broadly distributed throughout many regions of China, including 21 of 34 provinces. Because ragweed is a monoecious annual that reproduces with high fecundity via seeds, its mating patterns may be closely related to its invasiveness and dissemination ability. In China, alien invasive plants in the family Asteraceae that are characterized by high fecundity tend to retain a high potential for self-compatibility, while many self-incompatible invasive alien plant species are perennials with high vegetative reproduction ability. Although ragweed does not reproduce vegetatively, its breeding system has not been fully elucidated. Some studies have indicated that ragweed can readily generate mature seeds autogamously and allogamously, whereas other experimental evidence implies that this species is self-incompatible. To shed further light on the breeding system of ragweed, we transplanted plants from an invasive ragweed population in Changshu, Jiangsu Province, China, and used them in a series of common garden experiments. We studied patterns of pollen viability and stigma receptivity. We also investigated the ragweed breeding system and apomixis by measuring seed sets of different individual plants subjected to four different treatments: self-pollination, cross-pollination, open-pollination, and bagging of female capitula. Viable ragweed pollen was first detected four days after flowering, with viability peaking eight to ten days post-anthesis. Viable stigmas were observed on the second day, with viable levels reaching a maximum five to eight days after flowering. The overlapping period of pollen viability and stigma receptivity was thus about five days. Seed set of ragweed individuals was highest under open-pollination (48.4%, on average) and cross-pollination (44.4%, on average) treatments, although the difference between these two treatments was not significant (P> 0.05). Seed set of self-pollinated individuals was much significantly lower (P< 0.01) than values obtained under open pollination and cross pollination, but the average value, 3.4%, was significantly higher than zero (P< 0.05). In addition, no seed set was observed from bagged female capitula. Taken together, our results suggest that ragweed is mainly self-incompatible, but possesses slight self-compatibility and no apomictic ability. Over the long run, self-incompatibility enhances ragweed diversity and adaptation to newly invaded areas; initially, however, this characteristic does not promote its spread, as it is obviously difficult for a few individual ragweed plants to form a large population via seeds relying solely on hybridization. Self-incompatibility may thus be one of the major reasons why ragweed seldom infests small croplands subjected to intensive weed management practices. Consequently, removal of sporadic individuals from new ragweed distributional areas should be made a priority to prevent serious invasion.
Ambrosiaartemisiifolia; asteraceae; breeding system; invasive alien plants; self-incompatibility
江苏省自然科学基金面上项目(SBK201320980); 国家自然科学基金面上项目(31370548); 苏州市科技计划项目(SYN201304, YJG0911); 国家科技部国际合作与交流专项: 中国-东盟重大农业外来有害生物预警与防控平台(2011DFB30040)
2013-06-10;
日期:2014-05-16
10.5846/stxb201306101590
*通讯作者Corresponding author.E-mail: jhhao@cslg.cn
郝建华, 金洁洁, 陈国奇, 王立新.恶性入侵植物豚草的繁育系统特性.生态学报,2015,35(8):2516-2520.
Hao J H, Jin J J, Chen G Q, Wang L X.Breeding system of a noxious invasive alien plant,AmbrosiaartemisiifoliaL..Acta Ecologica Sinica,2015,35(8):2516-2520.
