Ddx1基因在不同发育阶段小鼠睾丸组织中的表达
2015-03-07赵秀军河北医科大学基础医学院组织胚胎学教研室河北石家庄050017
李 莉,张 雷,龚 淼,赵秀军,赵 昱(河北医科大学基础医学院组织胚胎学教研室,河北 石家庄 050017)
·论著·
Ddx1基因在不同发育阶段小鼠睾丸组织中的表达
李莉,张雷,龚淼,赵秀军,赵昱*(河北医科大学基础医学院组织胚胎学教研室,河北 石家庄 050017)
[摘要]目的系统观察不同日龄小鼠睾丸组织的发育特点,检测ddx1基因与蛋白在不同发育阶段小鼠睾丸组织中的表达模式,探讨ddx1基因在精子发生过程中的作用。方法采用石蜡切片HE染色的方法观察不同发育阶段小鼠睾丸组织的结构特征;采用实时荧光定量PCR法检测ddx1 mRNA在不同日龄小鼠睾丸组织中的表达;采用免疫印迹和免疫组织化学法检测DDX1蛋白在不同日龄小鼠睾丸组织中的表达特性与细胞内的定位。结果石蜡切片HE染色显示5、15、23、35、42和60日龄小鼠睾丸组织结构特点能代表生精上皮组织发生发育阶段;实时荧光定量PCR法显示ddx1 mRNA在各日龄小鼠睾丸组织中均有表达,于15日龄开始增高,随后维持稳定水平;免疫印迹法显示DDX1蛋白在35、42、60日龄小鼠睾丸组织中特异性表达并逐渐增高;免疫组织化学法显示在少量精原细胞、大量精母细胞和圆形精子细胞胞质与细胞核中存在DDX1蛋白阳性表达。 结论Ddx1基因和蛋白在小鼠精子发生过程中呈现阶段与细胞特异性表达, 提示ddx1基因可能在精子发生过程中发挥作用。
[关键词]精子发生;ddx1基因;睾丸;精母细胞;小鼠
doi:10.3969/j.issn.1007-3205.2015.12.001
精子发生是生精细胞特殊分化的复杂过程,各级生精细胞具有特异的基因表达与调控方式,此过程不仅需要多种基因正确表达,也需要多种RNA正确执行功能[1]。DEAD box家族蛋白是一个三磷酸腺苷(adenosine tripho-sphate,ATP)依赖的RNA解旋酶家族,参与RNA各种功能的执行[2]。研究显示该家族成员DDX3、DDX4、DDX25等与精子生成和男性不育有密切关系[3-6]。本实验前期研究发现正常成年男性睾丸内有ddx1基因与蛋白的表达。而有些进行睾丸穿刺的非梗阻性无精子症(non-obstructive azoospermia,NOA)患者的睾丸组织中没有ddx1基因与蛋白的特异表达。目前国内外尚未见到关于ddx1基因在人和小鼠睾丸组织中动态表达情况的报道。本研究连续观察了小鼠出生后2个月内多个时点的睾丸组织形态特征,确定小鼠精子发生过程与小鼠出生后日龄的具体对应关系,选取与首个精子发生波的时间点完全一致的5、15、23、35、42、60日龄小鼠,观察ddx1 基因与蛋白在小鼠精子发生过程中的表达情况,探讨ddx1基因在小鼠精子发生过程中可能发挥的功能,旨在为阐明生精障碍的病因和探索男性不育的靶向治疗提供理论与实验基础。
1材料与方法
1.1实验动物与主要试剂清洁级C57BL/6j雄性小鼠,购自北京华阜康生物科技股份有限公司。取出生后 1、3、5、7、9、11、13、15、18、23、27、30、33、35、39、42、45、48、51、54、58、60日龄小鼠,每组7只。兔抗鼠DDX1多克隆抗体(sc-134752,美国Santa Cruz公司);兔抗鼠β-actin多克隆抗体(sc-7210,美国Santa Cruz公司);二步法通用型免疫组织化学试剂盒(上海基因科技有限公司);ECL化学发光试剂盒(美国Abcam公司);PrimeScript®Ⅱ High Fidelity RT-PCR Kit(DR023A,宝生物公司);SYBR®Premix DimerEraser®试剂盒(DRR091A,宝生物公司)。
1.2石蜡切片制备用颈椎脱臼法将不同日龄小鼠处死,冰上快速取出睾丸放入4% 多聚甲醛固定4 h,取出切开睾丸组织后继续固定12~36 h。常规石蜡包埋,4 μm连续切片,60 ℃烤箱3 h,装盒备用。
1.3HE染色切片常规脱蜡、梯度酒精水合,苏木精染液染色3~10 min,自来水冲洗3 min,1% 盐酸酒精分化2 s,自来水冲洗3 min,5%氨水返蓝30 s,伊红染色3 min ,梯度酒精逐级脱水,二甲苯透明,中性树胶封片,Leica光学显微镜下观察并摄片。
1.4实时荧光定量PCR(Quantitative Real-Time PCR)法检测ddx1 mRNA的水平取液氮冻存的生后5、15、23、35、42、60日龄小鼠睾丸组织每份20 mg,应用TRIzol 法提取 RNA,选取凝胶电泳验证提取质量良好的RNA应用PrimeScript®Ⅱ High Fidelity RT-PCR试剂盒进行反转录,所得cDNA作为模板,参照SYBR®Premix DimerEraser®试剂盒说明书,将反应体系放置于 ABI-7300 Real-Time PCR仪中进行扩增反应。反应结束后确认扩增曲线和熔解曲线,证实无非特异性扩增,ddx1的相对表达量计算采用ΔΔCT法,以Gapdh (管家基因)、ddx25(已证实在小鼠睾丸不同发育阶段均有表达的基因)、Stra8(显示减数分裂起始的标志基因)、Prm1(显示长形精子细胞形成的特异性基因)作为对照,引物序列见表 1。

表1 实时荧光定量PCR所用引物序列
1.5免疫印迹(Western blotting)法检测DDX1蛋白在睾丸组织中的表达取液氮冻存的不同日龄小鼠睾丸组织,提取总蛋白并测定浓度,各组蛋白上样量为40 μg,SDS-PAGE胶分离,转膜,5%牛血清蛋白(bovine serum albumin,BSA)室温摇床封闭 2 h,去除封闭液,分别加入3%BSA稀释的兔抗鼠DDX1多克隆抗体(工作浓度为1∶100),兔抗鼠β-actin多克隆抗体(工作浓度为1∶1 000),4 ℃过夜,次日,室温平衡1h,加入用TBST稀释的辣根酶过氧化物标记的羊抗小鼠IgG(1∶200),室温垂直摇床孵育2 h,常规化学发光显色,在Gene-box成像系统中显影。
1.6免疫组织化学检测DDX1蛋白在睾丸组织中的表达与定位切片经脱蜡、水化后用10 mmol/L枸橼酸钠缓冲液微波加热至沸腾10 min进行抗原修复,用3% H2O2室温孵育 10 min 封闭内源性过氧化物酶的活性,滴加兔抗鼠DDX1多克隆抗体工作液(工作浓度为1∶50)避光孵育,4 ℃过夜,次日取出平衡至室温,滴加二抗复合物室温孵育 45 min,0.05%二氨基联苯胺(diaminoben zidine,DAB)溶液显色5~10 min,部分切片进行苏木精复染 3 min。阴性对照用 磷酸盐缓冲液替代一抗。常规脱水、透明、封片,显微镜下观察并摄片。
2结果
2.1不同日龄小鼠睾丸组织的形态结构5日龄小鼠睾丸生精小管的上皮内细胞层数2~3层,由大量支持细胞和少量精原细胞组成;睾丸间质细胞(Leydig 细胞)数量较少(图1A)。15日龄小鼠睾丸生精小管之间的间隙变小,部分小管可见管腔;生精上皮细胞层数为3~5层,初见前细线期精母细胞,体积较大,染色质松散;精原细胞位于基膜内侧,核染色质致密深染,可见有丝分裂相;Leydig细胞数量增多(图1B)。23日龄小鼠睾丸内生精上皮增厚,细胞层数明显增多,初见圆形精子细胞;精原细胞紧贴基膜,相对数量减少,核深染;初级精母细胞数量明显增多,粗线期精母细胞所占比例增大,细胞体积大,胞质嗜酸性,胞核呈绒球状;近管腔侧上皮内开始出现圆形精子细胞,体积较小,胞核呈均质状;偶见间质细胞成团分布,体积增大(图1C)。35日龄小鼠睾丸生精小管上皮明显增厚,可见精原细胞、精母细胞、圆形精子细胞和长形精子细胞,其中圆形精子细胞所占比例较大,可见较多处在形态改变过程中的精子细胞和处于各期的精母细胞;相邻生精小管之间可见成群分布的Leydig细胞,胞质嗜酸性强(图1D)。42日龄小鼠睾丸生精小管排列紧密,管腔明显;生精上皮内可见各种类型的生精细胞,其中长形精子细胞数量明显增多,偶见发育成熟的精子(图1E)。60日龄小鼠睾丸生精小管排列更紧密,间隙变小,管腔中精子增多;Leydig 细胞成群分布,细胞轮廓清晰,胞质嗜酸性增强(图1F)。

图1不同日龄C57BL/6j小鼠睾丸组织光镜结构(HE ×400)
A.5日龄;B.15日龄;C.23日龄;D.35日龄;E.42日龄;F.60日龄
Figure 1Morphological structure of normal C57BL/6j mouse spermatogenesis at different postnatal stages of the developing testis(HE ×400)
2.2Ddx1 mRNA在不同日龄小鼠睾丸组织中的表达Ddx1 mRNA在5日龄小鼠睾丸组织内已有微量表达,在15日龄即生精细胞开始进入减数分裂时期的小鼠睾丸组织中转录水平略有升高,随后逐渐升高,直到60日龄维持稳定水平;ddx25 mRNA 自生后即有持续表达;Stra8 mRNA 从出生后15日龄小鼠即有一定转录水平,到23 d龄转录水平有显著升高,随后维持较低水平;Prm1 mRNA自23 d出现微量特异性转录,至35日龄转录水平达最高,随后维持较高水平(图2)。
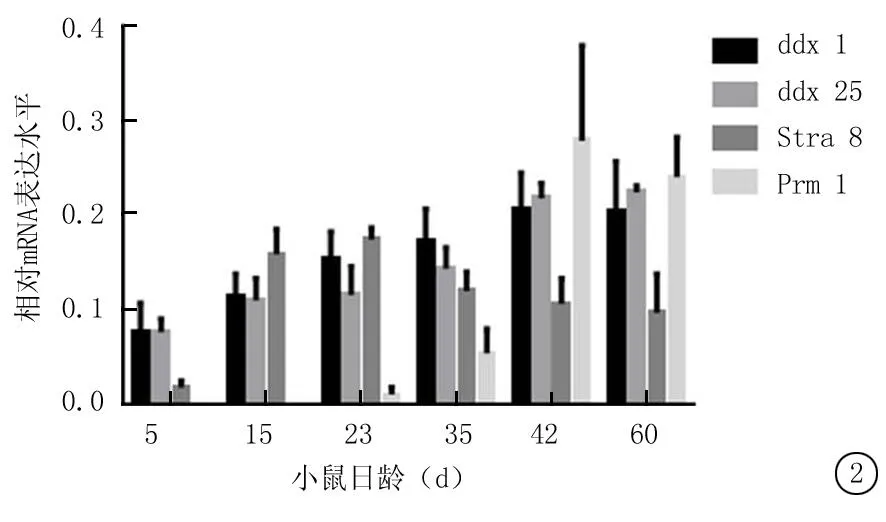
图2实时荧光定量PCR法检测ddx1 mRNA在不同日龄小鼠睾丸组织中的表达
Figure 2The expression of ddx1 mRNA by quantitative real-time PCR in mouse testis at different ages
2.3DDX1蛋白在不同日龄小鼠睾丸组织中的表达5、15和23日龄小鼠睾丸组织中无DDX1蛋白条带出现;35、42、60日龄小鼠睾丸组织中可见相对分子质量为82 000处有条带出现(图3)。
2.4DDX1蛋白在不同日龄小鼠睾丸组织中的表达与定位5、15和23日龄小鼠睾丸组织中未见DDX1蛋白的表达(图4B、C、D);DDX1蛋白的阳性
表达首先出现于35日龄小鼠睾丸生精细胞的胞质中,呈弱阳性表达(图4E),主要在精母细胞与圆形精子细胞胞质中表达(图4F);在42日龄小鼠睾丸组织中DDX1蛋白则分别在睾丸间质细胞胞质,精原细胞、精母细胞和圆形精子细胞的胞质与胞核中表达,其中精母细胞和圆形精子细胞中的阳性表达较强,长形精子细胞的胞核无表达(图4G、H);60日龄小鼠睾丸组织中DDX1蛋白的表达与42日龄小鼠睾丸组织中的表达情况一致(图4I、J)。对照组为阴性(图4A)。

图3免疫印迹法检测DDX1蛋白在不同日龄小鼠睾丸组织中的表达
Figure 3The expression of DDX1 by Western blotting in mouse testis at different ages
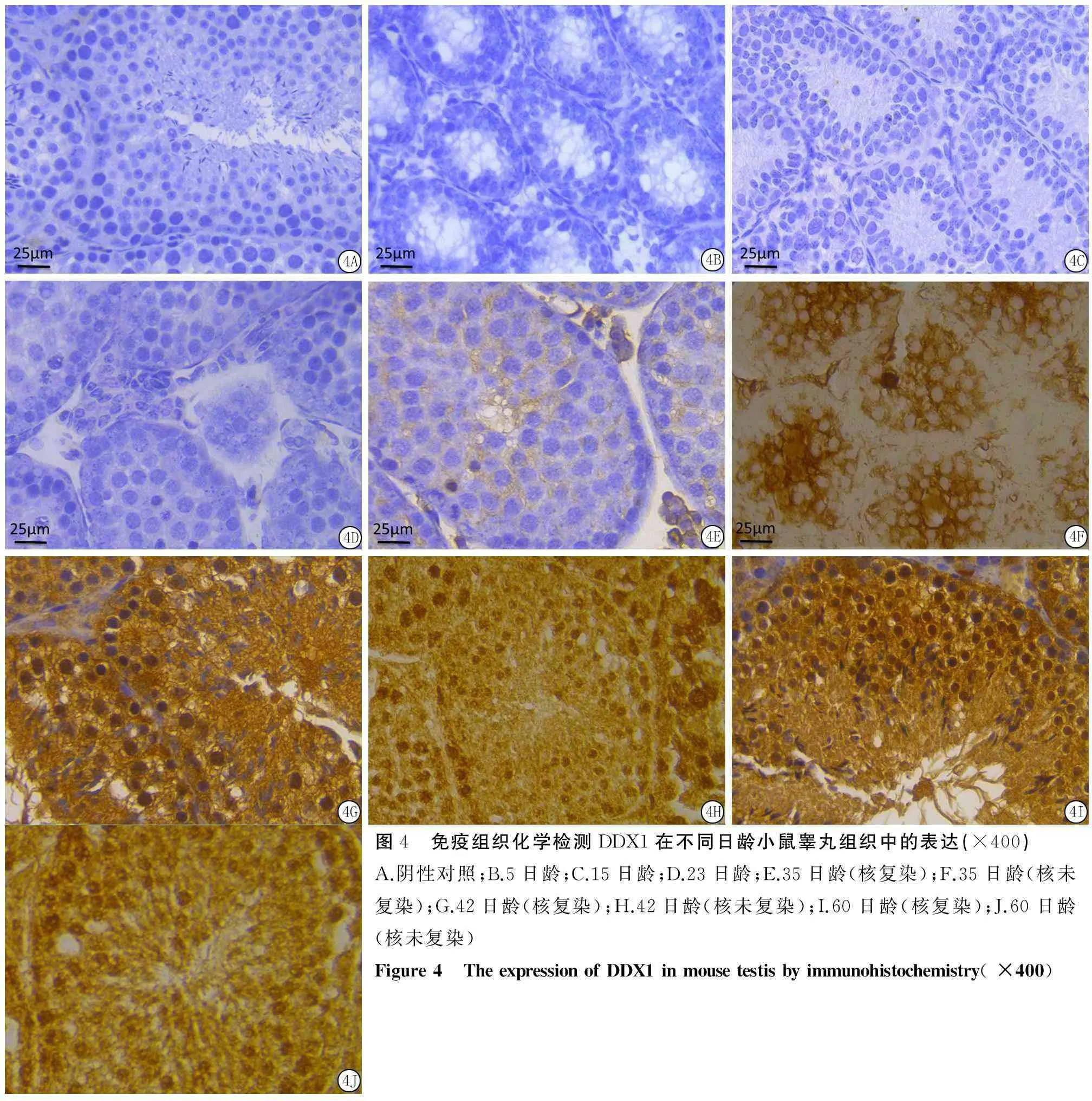
图4 免疫组织化学检测DDX1在不同日龄小鼠睾丸组织中的表达(×400)A.阴性对照;B.5日龄;C.15日龄;D.23日龄;E.35日龄(核复染);F.35日龄(核未复染);G.42日龄(核复染);H.42日龄(核未复染);I.60日龄(核复染);J.60日龄(核未复染)Figure4 TheexpressionofDDX1inmousetestisbyimmunohistochemistry(×400)
3讨论
随着生殖健康研究的深入开展,男性不育的病因研究取得了较大进步。男性不育症患者的精液中没有精子,通常被称为NOA。中国有25%的NOA患者的长辈中有NOA患者。研究证实,相当比例的NOA患者的病因是基因突变[7]。近年来与男性不育相关的基因突变或多态现象日益成为研究热点。因此,研究精子发生相关基因的特性与功能,将为男性生精障碍发病机制的阐明、临床诊断、靶向治疗与男性避孕奠定基础。本研究前期发现ddx1基因在小鼠的脑、肝、肾和睾丸组织中均有表达,但DDX1蛋白仅在脑和睾丸组织中检测到,为进一步研究ddx1基因与蛋白在小鼠睾丸组织中的确切表达与其在精子发生中的作用,本研究先从显微组织学水平入手,将生后0~60日龄的C57BL/6j小鼠的睾丸组织进行石蜡包埋并连续切片,系统观察该小鼠睾丸组织结构与功能的变化规律,较完整地揭示小鼠精子发生过程各阶段与小鼠生后日龄的具体对应关系,旨在为后续基因与蛋白分子水平的功能研究提供实验基础。本研究形态学结果显示,C57BL/6j小鼠在精子发生过程中,生精细胞的减数分裂在出生后第15天左右开始,生精上皮内偶见前细线期精母细胞,到第18天左右,生精上皮内的粗线期精母细胞的数量增加明显,到第23天时,可见少量减数分裂完成而形成的圆形精子细胞。第25天圆形精子细胞进入精子形成阶段,形态开始发生改变,到第35天生精上皮内可见到少量形态改变即将完成的长形精子细胞。第42天在生精上皮的管腔侧可见到长出鞭毛的精子,提示小鼠睾丸组织中的第1个精子发生波结束,第60天小鼠生精小管内可见大量精子提示小鼠进入成年期。因此,本研究选取第1个精子发生波相关的时间点,即生后第5、15、23、35、42天以及成年期生后第60天作为对比,对ddx1基因与蛋白在睾丸组织中的动态表达进行研究,探讨其在小鼠精子发生过程中的作用。
在精子发生的多种调控因素中起决定性作用的是生精细胞内基因水平的调控[8]。许多特异性基因均具有各发育阶段的特异性表达和不同生精细胞的特异性表达的特征[9]。研究特定基因在精子发生中的作用需从该基因在精子发生过程中的基因与蛋白的表达入手。因此,本研究选取与第1个精子发生波时间点相对应的不同生后日龄,减数分裂开始对应15日龄,染色质的浓缩对应23日龄,形态改变过程对应35日龄,精子形成对应42日龄。一般情况基因特异表达于精子发生过程中的某种生精细胞,则该基因的转录产物将出现在睾丸发育的某个特定时期,并与精子发生的特定阶段一致。本研究从基因水平开始,首先探讨ddx1 mRNA是否表达于不同日龄C57BL/6j小鼠睾丸组织内。实时荧光定量PCR检测结果显示不同日龄小鼠睾丸组织中均有ddx1 mRNA的表达,表达水平呈现逐渐升高的趋势,于42日龄后趋于稳定,表达水平与60日龄无明显差异,提示ddx1基因在精子发生过程中未表现出明显的时空特异性,但在睾丸组织内确实有ddx1 mRNA的表达。同时,本研究将ddx25、Stra8和Prm1作为对照[10-12],结果显示,在不同日龄小鼠睾丸组织中均可检测到ddx25的表达,15日龄小鼠睾丸内Stra8 mRNA的表达开始升高,35日龄小鼠睾丸组织中开始出现Prm1 mRNA的表达,于42日龄达到最高。提示小鼠睾丸组织RNA的提取与检测方法正确,小鼠生精细胞于15日龄左右开始进入减数分裂阶段,35日龄小鼠睾丸组织中开始出现长形精子细胞,于42日龄长形精子细胞数量明显增加,进一步验证了对小鼠睾丸发育阶段组织学观察的正确性。有研究显示,在精子发生过程中仅有20%左右的基因表达与蛋白表达具有相关性,大多数基因与蛋白表达的特异性并不一致[13]。同时,精子形成后不再进行转录,精子发生的相关基因在精子发生早期即开始进行转录,并可在睾丸组织中检测到该基因的表达,但转录产物没有立即翻译形成蛋白,在数日内呈抑制状态,直到精子发生至特定阶段需要特定蛋白表达时才进行翻译[14],即基因表达水平并不代表蛋白水平的表达。因此,本研究考虑ddx1基因很可能以上述这种方式对精子发生进行调控,于是接着进行DDX1蛋白在睾丸组织与生精细胞中表达与定位的研究。
本研究分别应用免疫印迹法检测不同日龄小鼠睾丸组织中DDX1蛋白的总体表达情况和免疫组织化学方法检测DDX1蛋白在不同生精细胞中的表达定位。免疫印迹结果显示,5、15和23日龄小鼠睾丸组织中未见DDX1蛋白的表达,35日龄小鼠睾丸组织内开始出现DDX1蛋白的表达,随着日龄的增加,蛋白的表达水平逐渐增高。提示ddx1基因的表达与蛋白的表达情况确实不一致,转录后直到减数分裂后期才进行翻译形成蛋白产物表达于睾丸组织。为了进一步探讨DDX1蛋白的表达是否具有阶段与细胞特异性,本研究采用免疫组织化学法检测不同日龄小鼠睾丸组织中DDX1蛋白的表达与定位,结果与免疫印迹结果一致,5、15和23日龄小鼠睾丸组织中未见DDX1蛋白的表达;阳性表达首先出现于35日龄小鼠精母细胞与圆形精子细胞胞质中;在42日龄小鼠睾丸组织中DDX1蛋白则分别表达于精原细胞、精母细胞和圆形精子细胞的胞质与胞核,其中精母细胞和圆形精子细胞中的阳性表达较强,长形精子细胞的胞核未见表达。提示ddx1基因在小鼠睾丸圆形精子细胞的形成与形态改变过程中发挥作用,虽然不参与减数分裂的起始,但对维持减数分裂的正常进行可能发挥作用。RNA解旋酶家族几乎参与全部与RNA相关的生命活动,如RNA二级结构的改变,转录起始与调控,线粒体RNA剪接,核糖体和剪接体装配,mRNA降解与稳定性维持,翻译调控等[15]。Ddx1作为该家族的成员之一,也具有上述一些功能。本研究结果显示DDX1蛋白在生精细胞的表达中有时空特异性,但与其基因的表达不完全一致。提示ddx1基因可能在转录后翻译水平在精子发生过程中发挥作用。本研究结果还显示42日龄小鼠睾丸间质细胞的胞质中有DDX1蛋白的表达。提示ddx1基因可能与睾丸间质细胞合成分泌雄激素的功能发挥有关。
综上所述,ddx1基因在不同日龄小鼠睾丸组织中表达,但无阶段特异性,DDX1蛋白在不同日龄小鼠睾丸组织中的表达有时空特异性与生精细胞特异性。提示ddx1基因在精子发生过程中调控作用可能发生在转录后翻译水平,可为临床生精障碍的诊治提供新指标。为了探讨ddx1基因在精子发生过程中的精确作用,本研究后期将把RNA干扰技术与生精细胞体外培养技术结合进行深入研究。
[参考文献]
[1]Senti KA,Jurczak D,Sachidanandam R,et al.piRNA-guided slicing of transposon transcripts enforces their transcriptional silencing viaspecifying the nuclear piRNA repertoire[J].Genes Development,2015,29(16):1747-1762.
[2]Cherry AA,Ananvoranich S.Characterization of a homolog of DEAD-box RNA helicases in Toxoplasma gondii as a marker of cytoplasmic mRNP stress granules[J].Gene,2014,543(1):34-44.
[3]Singh P,Schimenti JC.The genetics of human infertility by functional interrogation of SNPs in mice[J].Proc Natl Acad Sci USA,2015,112(33):10431-10436.
[4]Celik H,Sajwan KP,Selvanathan SP,et al.Ezrin binds to dead-box rna helicase ddx3 and regulates its function and protein level[J].Mol Cell Biol,2015,35(18):3145-3162.
[5]Leng L,Tan Y,Gong F,et al.Differentiation of primordial germ cells from induced pluripotent stem cells of primary ovarian insufficiency[J].Human Reprod,2015,30(3):737-748.
[6]Kavarthapu R,Tsai-Morris CH,Fukushima M,et al.A 5′-flanking region of gonadotropin-regulated testicular RNA helicase (GRTH/DDX25) genedirects its cell-specific androgen-regulated gene expression[J].Endocrinology,2013,154(6):2200-2207.
[7]Vloeberghs V,Verheyen G,Haentjens P,et al.How successful is TESE-ICSI in couples with non-obstructive azoospermia? [J].Hum Reprod,2015,30(8):1790-1796.
[8]Yuan Y,Zhou Q,Wan H,et al.Generation of fertile offspring from Kit(w)/Kit(wv) mice through differentiation of gene corrected nuclear transfer embryonic stem cells[J].Cell Res,2015,25(7):851-863.
[9]Coutton C,Escoffier J,Martinez G,et al.Teratozoospermia:spotlight on the main genetic actors in the human[J].Hum Reprod Update,2015,21(4):455-485.
[10]Takebe M,Onohara Y,Yokota S.Expression of MAEL in nuage and non-nuage compartments of rat spermatogenic cells and colocalization with DDX4,DDX25 and MIWI[J].Histochem Cell Biol,2013,140(2):169-181.
[11]Sahin P,Sahin Z,Gungor-Ordueri NE,et al.Inhibition of mammalian target of rapamycin signaling pathway decreases retinoic acid stimulated gene 8 expression in adult mouse testis[J].Fertil Steril,2014,102(5):1482-1490.
[13]Hu JR,Liu M,Hou CC,et al.Gene expression pattern of KIFC3 during spermatogenesis of the skink Eumeces chinensis[J].Gene,2015,556(2):206-212.
[14]Krieger T,Simons BD.Dynamic stem cell heterogeneity[J].Development,2015,142(8):1396-1406.
[15]Desai SS,Roy BS,Mahale SD.Mutations and polymorphisms in FSH receptor:functional implications in human reproduction[J].Reproduction,2013,146(6):R235-248.
(本文编辑:许卓文)
Developmental expression of ddx1 in mouse testis
LI Li,ZHANG Lei,GONG Miao,ZHAO Xiu-jun,ZHAO Yu*
(Department of Histology and Embryology,the School of Basic Medical Science,Hebei Medical University,Shijiazhuang 050017,China)
[Abstract]ObjectiveTo identify the different developmental stages of testis in mice.To investigate the dynamic expression of ddx1 mRNA and protein in mouse testis as well as the function of ddx1 during spermatogenesis.MethodsParaffin sections were used with H&E staining to show morphological structure of normal mouse spermatogenesis at postnatal stages of the developing testis.Quantitative real-time PCR was used to establish the ddx1 mRNA expression in the testis of mice at different ages.Immunohistochemistry and Western blotting were carried out to detect the expression and distribution of DDX1 protein in mouse testis and spermatocytes.ResultsHistological analysis showed morphological structure of normal mouse spermatogenesis at different postnatal stages of the developing testis from 5,15,23,35,42,60-day old mice.Quantitative real-time PCR showed that express of ddx1 mRNA situate at low relatively at postnatal P5,then increased at P15 and retained high afterwards.Western blotting confirmed that the specific expression of DDX1 protein in mouse testis.The DDX1 antigen was detected in testis of 35,42 and 60-day old and located mainly in the cytoplasm and nuclear of spermatocyte and round spermatid by immunohistochemistry.ConclusionThe ddx1 mRNA and protein expressed in the testis of mice at different stage during spermatogenesis.It indicated that ddx1 may be involved in the regulation of spermatogenesis and spermiogenesis.
[Key words]spermatogenesis;ddx1;testis;spermatocyte;mice
[中图分类号]R321.1
[文献标志码]A
[文章编号]1007-3205(2015)12-1365-06
[作者简介]李莉(1976-),女,河北石家庄人,河北医科大学基础*通讯作者。E-mail:441678544@qq.com
[基金项目]国家自然科学基金项目(81373787)
[收稿日期]2015-09-14;[修回日期]2015-11-21
医学院讲师,医学博士,从事生殖生物学与组织学研究。
