西辽河河岸草本植物物种特征及其与环境因子的关系
2015-03-01孔维静夏会娟3张远侯利萍孙丽慧张
孔维静夏会娟, 3张 远侯利萍孙丽慧张 楠
(1. 中国环境科学研究院流域水生态保护技术研究室, 北京 100012; 2. 环境标准与风险评估国家重点实验室, 北京 100012; 3. 北京林业大学林学院森林培育与保护教育部重点实验室, 北京 100083)
研究简报
西辽河河岸草本植物物种特征及其与环境因子的关系
孔维静1, 2夏会娟1, 2, 3张 远1, 2侯利萍1, 2孙丽慧1, 2张 楠1, 2
(1. 中国环境科学研究院流域水生态保护技术研究室, 北京 100012; 2. 环境标准与风险评估国家重点实验室, 北京 100012; 3. 北京林业大学林学院森林培育与保护教育部重点实验室, 北京 100083)
河岸带是水体与陆域之间的过渡区域[1], 受到洪水等河流过程以及来自邻近高地的影响, 形成多样、动态的复杂生境和特殊的结构功能特征。河岸植被是河岸带生态系统的重要组成部分, 具有污染物消减、微气候调节、生态廊道以及提供栖息地等生态功能[2], 在河岸带功能的发挥中具有重要作用。受到河谷地貌、洪水强度、频率、地形和土壤性质的小尺度变化, 河流自上游向下游不同生物区系的气候变化以及陆地对河流的影响, 河岸植物在物种组成、空间分布和群落多样性等生态系统特征中具有区域特性[3]。
已有研究表明, 河岸带植被以草本植物为主[4, 5]。草本植物是群落的先锋植物, 有较宽的生态位, 能较快地适应河岸带环境[6]。而河岸带生境的复杂性以及人为和自然干扰的影响使草本植物的物种组成具有不稳定性和多变性, 群落空间分布更加不规则[6]。另外, 草本植物比乔木和灌木对干扰、干旱胁迫或地貌异质性更加敏感[7—10]。因此, 河岸带草本植物群落特征能反映河岸带生态系统受自然或人为因素干扰的影响。国际上已有学者从不同角度对河岸带植物群落特征进行了研究, 阐明了河岸带群落的物种组成[11]、植被演替过程[12]、植被功能[1]和生物多样性[13, 14]等特征, 而在我国河岸带植被的研究开展较少, 仅在少数河流开展了河岸带草本植物功能群划分[15]、物种组成[16, 17]和群落物种多样性[6, 18]等研究。
辽河流域是我国东北地区南部最大的河流, 已有的研究曾涉及辽河流域[19, 20]、辽河三角洲的植被结构[21]、辽河太子河大型水生植物群落特征及其与环境的关系[22],而对西辽河河岸带植被的研究鲜有涉及, 仅在东辽河[18]和辽河干流[17]河岸带开展了少量研究。西辽河流域是辽河流域的重要组成部分, 本文基于野外调查, 研究了西辽河及其两条支流老哈河和西拉木伦河的河岸带草本植物物种特征, 包括植物物种组成、物种生活型和水分生态类型、物种多样性, 分析了支流间河岸带草本物种特征的差异, 以及影响物种多样性的主要环境因素。通过本研究以期为河岸带恢复中物种的筛选以及河流保护和管理提供依据。
1 研究地区与研究方法
1.1 研究区域概况
西辽河是我国七大江河之一辽河的支流, 位于119°04′—125°01′E, 42°00′—45°00′N, 干流全长 829 km,流域面积1.48×105km2, 占辽河流域总面积的65%[23]。西辽河发源于河北省, 但水系绝大部分流经内蒙古东北部的赤峰和通辽市境内。西辽河有二源: 一是河北平泉县光头岭的老哈河; 另一源是内蒙古克什克腾旗白岔山的西拉木伦河(图 1)。老哈河干流全长约 443 km, 流域面积2.74×104km2, 海拔280—1500 m。从河源至黑里河口, 为老哈河的上游段, 两岸山高谷深, 河网比较发育, 河床多为砂砾石; 由黑里河至红山水库为老哈河中游段, 河槽呈宽浅型, 河流两岸比较开阔, 水量增加, 河床摆动较多;由红山水库至海力吐汇合口处是老哈河的下游段, 该河段两岸多为沙质丘陵台地。西拉木伦河干流全长约333 km, 流域面积3.22×104km2, 海拔280—1900 m。西拉木伦河从源头到百岔河口为上游段, 地貌多为海拔高于1000 m的山地, 河道弯曲, 河谷狭窄, 落差集中, 水流湍急, 坡度较大, 河床稳定; 从百岔河口至海拉苏为中游河段, 河槽呈宽浅型, 主流摆动不定, 多处出现河心沙洲,沿河两岸多为沙坨台地, 流域地貌绝大部分为海拔在500—1000 m 的低山丘陵; 海拉苏至西辽河干流为下游,河道较为平缓, 地势比较平坦, 其地貌为平原。西辽河流域地处中高纬度, 大部分地区气温均在 4℃以上, 日照时数较长。年降水量343—451 mm, 年内分配不均匀, 集中在 7、8月份, 占全年的 60%左右, 冬季降水量少。西辽河多年平均蒸发量达2000 mm以上, 年内月份最大蒸发量出现在5月, 达240—450 mm左右。
1.2 野外调查
根据水系布设样点, 获得老哈河 14个样地(自上而下依次编号为 1—14)和西拉木伦河 26个样地(自上而下依次编号为 15—40)的河岸植被数据, 采样点分布如图 1所示。2012年8月20日—9月19日植物生长季, 采用样线法进行植物群落调查。调查时在每个样地平行于河流流向紧邻河水边缘设置一条长50 m宽1 m的样线, 通常样线距河水边缘 0—5 m, 记录样地的经纬度及海拔, 并描述样地受到的人为干扰程度和河岸基质类型; 调查记录样线内的草本物种名称、个体数和高度, 估测样线内物种的盖度。根据《内蒙古植物志》[24]鉴定物种, 对暂时不能确定的物种采集标本带回实验室, 请专家鉴定。
1.3 数据处理与分析
植物物种组成 根据《内蒙古植物志》[24]鉴定植物的科属种信息, 统计物种的科属组成。
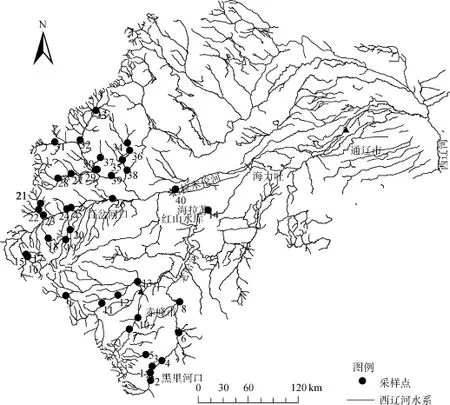
图1 西辽河流域及采样点分布图Fig. 1 Overview of the West Liaohe River watershed and sampling sites
物种生活型和水分生态型划分 根据《中国植被》[25]对物种生活型和水分生态型的定义, 参考植物志, 并根据野外调查物种生境进行划分; 参考《内蒙古植物志》将植物物种划分为中旱生、旱中生、中生、湿中生、湿生和水生6种水分生态型[24]; 参考《中国植物志》将植物物种分为1年生、1—2年生、2年生和多年生4种生活型[26]。
物种多样性指数计算 频度和盖度可以反映物种在研究区的重要性, 计算公式为:

物种多样性指标选用 Patrick丰富度指数、Shannon-Wiener多样性指数、Simpson多样性指数和Pielou均匀度指数, 其计算公式如下[27]:

式中: S为样线内的物种数; Pi为某种的个体数占总个体数的比例。
冗余分析(RDA) 排序选用的模型根据DCA排序轴长度确定, 如果第 1轴的长度>4.0, 就选单峰模型(CCA、CA、DCA); 如果在3.0—4.0, 那么选单峰和线性模型(PCA、RDA)均可; 如果<3.0, 则选线性模型[28]。本文第一轴的长度<3.0, 选用 RDA排序分析物种多样性与环境因子之间的关系, 用偏蒙特卡罗置换检验评估每个环境因子对物种多样性的解释贡献。排序在CANOCO4.5软件下进行。
环境因子的获取 选择海拔、河流等级、河道宽度和湿宽(河流水面宽度) 4个环境因子, 分析其与植物群落特征的关系。在调查中, 使用 GPS获取样地的海拔信息, 用测距仪测定河道宽度和湿宽。基于ArcGIS软件下的西辽河水系分布图, 根据Strahler河流等级分级方法划分河流等级[30]。
数据分析 使用Excel软件对物种、群落数据进行基本统计分析。
单因素方差分析(one-way ANOVA)、相关性分析等统计分析在SPSS 17.0下完成。显著性水平为α=0.05。
2 结果
2.1 植物物种组成
西辽河河岸带共记录到109种草本植物, 分属25科68属。蕨类植物只有木贼科(Equisetaceae)木贼属(Equisetum)问荆(Equisetum arvense)1种, 其余全为被子植物; 被子植物中双子叶植物63种, 分属17科38属, 单子叶植物45种, 分属7科29属。老哈河河岸带记录到草本植物57种, 分属19科42属。西拉木伦河河岸带记录到草本植物90种, 分属22科57属。
植物科的物种组成 西辽河河岸带草本植物科的物种组成以禾本科(Gramineae)、菊科(Compositae)、蓼科(Polygonaceae)和莎草科(Cyperaceae)物种数较多, 其中禾本科所含属数和种数最多, 主要为稗属(Echinochloa)植物、芦苇(Phragmites australis)和拂子茅(Calamagrostis epigeios)等 23种。老哈河和西拉木伦河河岸草本植物中禾本科植物分别有12和19种, 占各自总物种数的21.05% 和 21.11%。虽然含种数较多的科仅有 4科, 但它们在西辽河、老哈河和西拉木伦河所含属分别占各自总属数的51.47%、50.00%和 56.14%; 所含种分别占各自总种数的57.80%、57.89%和60.00%(表1)。只出现1种物种的科较多, 在西辽河有苋科(Amaranthaceae)、葫芦科(Cucurbitaceae)、马齿苋科(Portulacaceae)和眼子菜科(Potamogetonaceae)等10科, 占本流域内全部科的40.00%; 老哈河包括藜科(Chenopodiaceae)、毛茛科(Ranunculaceae)、苋科和十字花科(Cruciferae)等9科, 占本流域内全部科的47.37%; 西拉木伦河包括玄参科(Scrophulariaceae)、马齿苋科、桑科(Moraceae)和大戟科(Euphorbiaceae)等9科, 占本流域内全部科的40.91%。
植物属的物种组成 西辽河、老哈河和西拉木伦河河岸带草本植物只出现 1种物种的属最多, 分别占各自总属数的 73.53%、80.95%和 71.93%, 如菵草属(Beckmannia)、芦苇属(Phragmites)、水莎草属(Juncellus)、旋覆花属(Inula)和苍耳属(Xanthium)等。在西辽河出现物种数大于 5的属仅有蒿属(Artemisia)和蓼属(Polygonum);老哈河和西拉木伦河的蓼属植物分别有6种和7种; 老哈河仅有1种蒿属植物, 西拉木伦河有7种。大多数属所出现的物种数在2—5之间, 包括莎草属(Cyperus)、酸模属(Rumex) 藨、 草属(Scirpus)和车前属(Plantago)等(表2)。
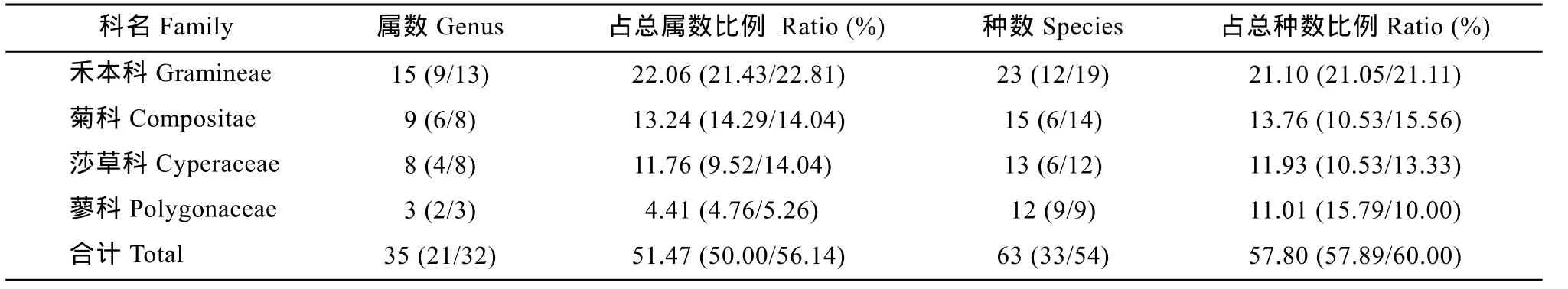
表1 西辽河河岸带草本植物主要科内属和种的组成Tab. 1 Riparian herbaceous genera and species composition among dominant families in the West Liaohe River
2.2 草本植物物种的频度和盖度
西辽河河岸带109种草本植物物种中, 频度≥30%的有15种, 占全部物种的13.76%; 频度≥50%的有6种, 占全部物种的5.50%, 其中桃叶蓼(Polygonum persicaria)和无芒稗(Echinochloa crusgalli var. mitis)的频度≥70%; 频度≥30%的物种中平均盖度≥5%的有8种, 其中桃叶蓼、拂子茅和芦苇的平均盖度≥10% (表3)。
老哈河河岸带 57种草本植物物种中, 频度≥30%的有13种, 占全部物种的22.81%; 频度≥50%的有5种, 占全部物种的 8.77%, 其中桃叶蓼的频度≥70%; 频度≥30%的物种中平均盖度≥5%的有8种, 平均盖度≥10%的有7种, 其中桃叶蓼和蒌蒿(Artemisia selengensis)的平均盖度≥20%(表3)。西拉木伦河河岸带90种草本植物物种中, 频度≥30%的有 16种, 占全部物种的 17.78%; 频度≥50%的有9种, 占全部物种的10%, 其中桃叶蓼和无芒稗的频度≥70%; 频度≥30%的物种中平均盖度≥5%的仅有4种, 其中拂子茅和芦苇的平均盖度≥10%(表3)。
2.3 草本植物生活型和水分生态型
西辽河河岸带草本植物的生活型以多年生草本植物为主, 占全部物种的50.46%, 其次为1年生植物, 占全部物种的 42.20%。对比两条支流的物种生活型发现, 老哈河以 1年生植物为主, 所占比例为 47.37%; 西拉木伦河以多年生植物为主, 所占比例为52.22%。
西辽河河岸带 109种草本植物中, 中生植物物种数最多, 达62种, 占全部物种的56.88%; 其次为湿生植物, 有25种, 所占比例为22.94%; 水生、湿中生、旱中生和中旱生植物较少, 分别有5、9、6和2种。对比两条支流河岸带草本植物水分生态型发现二者均以中生植物占优势, 所占比例分别为56.14%和57.78%; 其次为湿生植物,所占比例分别为24.56%和23.33%; 老哈河无中旱生和旱中生2种水分生态型。
2.4 草本植物物种多样性及其与环境因子的关系
老哈河和西拉木伦河河岸带草本植物物种多样性之间无显著差异(P>0.05)。老哈河的Shannon-Wiener指数、Simpson多样性指数、Pielou均匀度和丰富度指数的变化范围分别是1.23—2.21、0.55—0.87、0.54—0.86和5—14,各指数在源头区较低, 且自上游至下游呈增加的趋势(图2)。西拉木伦河的Shannon-Wiener指数、Simpson多样性指数、Pielou均匀度和丰富度指数的变化范围分别是1.14—2.26、0.49—0.84、0.47—0.81和 11—20, 各指数在源头区最低, 但在自上游至下游无显著变化。
西辽河各样地的环境因子数据如表 4所示, 物种多样性与环境因子的RDA排序结果表明(图3): 物种丰富度与海拔显著正相关(P<0.05); Shannon-Wiener指数、Simpson多样性指数和Pielou均匀度指数与河道宽度和湿宽负相关(P<0.05), 各多样性指数与河流等级无显著相关性(P>0.05)。湿宽对物种多样性的解释率最高, 为 50%;其次为海拔, 其解释率为25.4%; 河宽对物种多样性的解释率为15.5%; 河流等级的解释率最低, 仅为9.2%。
3 讨论
3.1 植物物种组成、生活型和水分生态型
西辽河河岸带草本植物物种组成呈现单种科和单种属较多, 出现物种较多的科只有菊科、禾本科、莎草科和蓼科植物, 它们在物种组成中占优势。在本研究中菊科和禾本科的物种占优势, 这与其他地区的河岸带植物物种组成一致[4—6]。本研究中出现的蓼科和莎草科植物多为湿生或水生物种, 这与河岸紧邻水缘, 土壤水分充足, 生境适宜有关; 而且两科植物物种种子体积小、重量轻、数量多, 利于传播和定居有关, 因此在物种组成中占有较大的比例[18]。
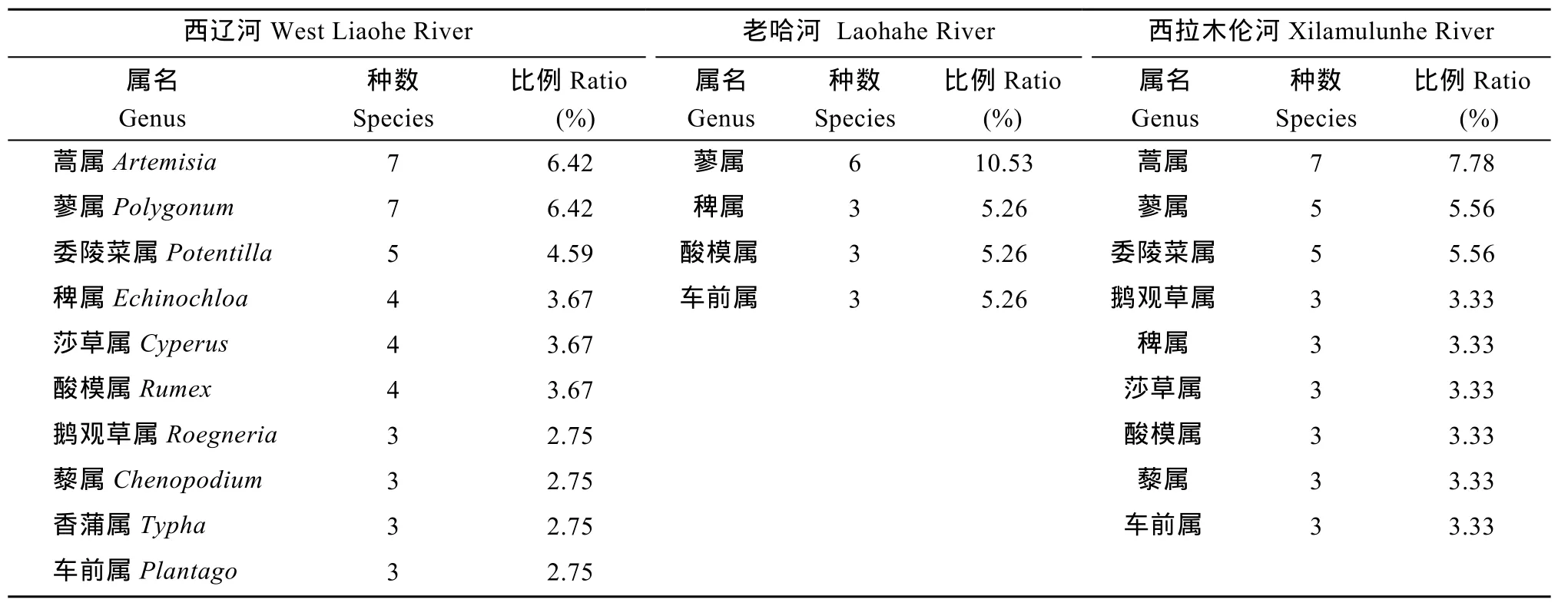
表2 西辽河河岸带草本植物主要属(物种数≥3)的物种组成Tab. 2 Riparian herbaceous species composition among genera in the West Liaohe River
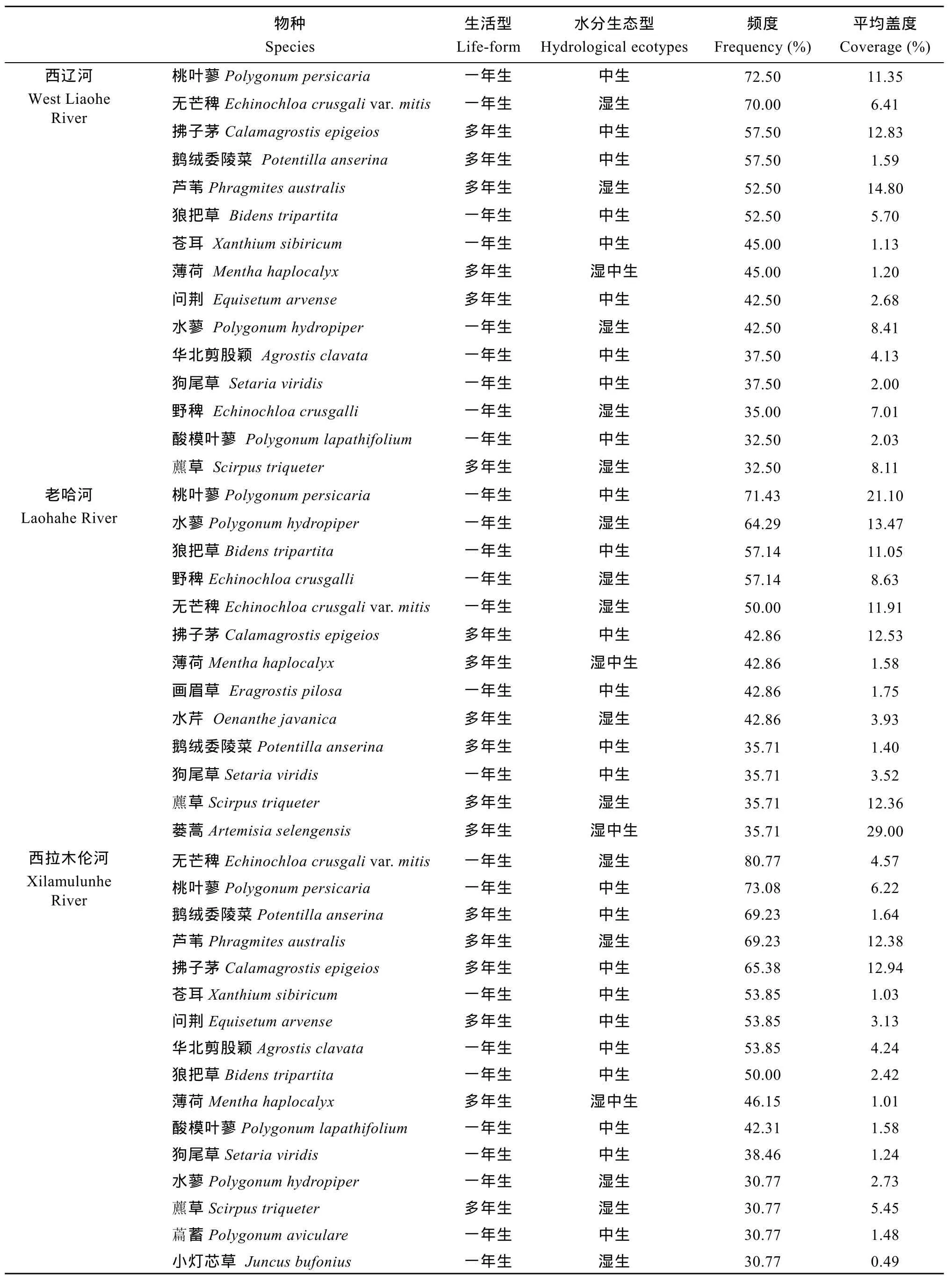
表3 西辽河及其支流河岸草本植物物种(频度≥30%)的基本信息Tab. 3 Information of riparian herbaceous species (frequency≥30%) in the West Liaohe River and its tributaries
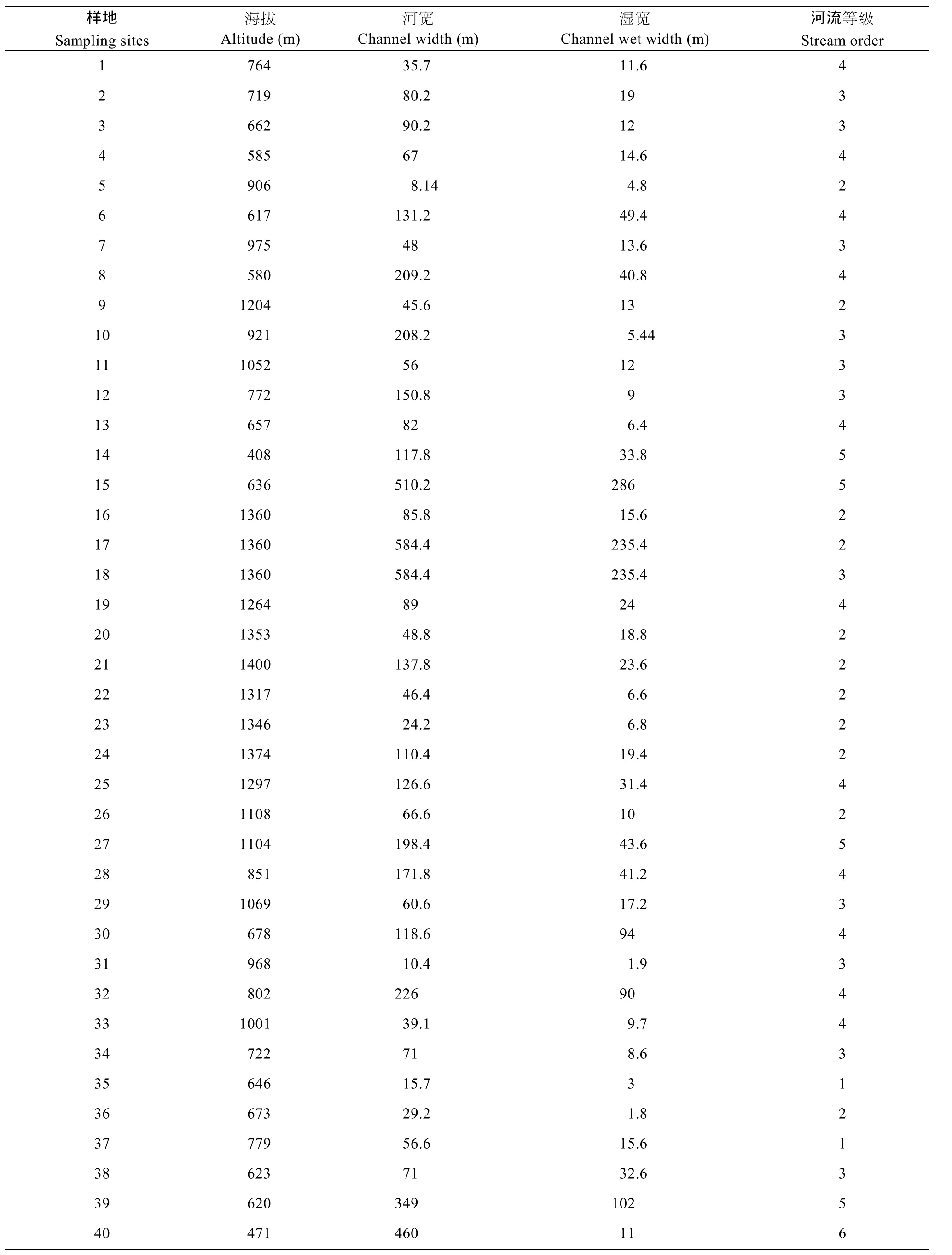
表4 调查样地的环境因子信息Tab. 4 Environmental factors of different investigated sites
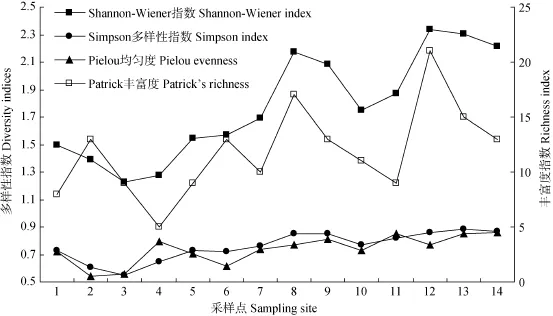
图2 老哈河河岸带草本植物物种多样性自上游至下游的变化Fig. 2 Riparian herbaceous species diversity along the Laohahe River

图3 西辽河河岸带草本植物物种多样性与环境因子的RDA排序图Fig. 3 RDA ordination of riparian herbaceous species diversity in relation to environmental factors
老哈河和西拉木伦河河岸带草本植物水分生态型均以中生植物占优势, 其次为湿生植物。老哈河河岸带草本植物群落中出现频度高且平均盖度大的物种有蒌蒿、桃叶蓼、水蓼、藨 草、拂子茅、无芒稗和狼把草, 而在西拉木伦河仅有拂子茅和芦苇2种。这些物种除拂子茅为中生植物外, 其余均为湿生植物。这说明虽然中生植物在物种组成中占有较大比例, 但在草本植物群落中多以湿生植物为优势种, 这和东辽河的研究结果一致[18]。所以在河岸植被恢复时, 出现频度高且盖度大的湿生植物可以作为优选物种。根据孟伟等[29]的辽河流域水生态分区, 西拉木伦河全流域属于辽西北缺水区, 老哈河中下游虽然也属于缺水区, 但其源头区属于辽中辽南少水区, 陆地水分条件是影响河岸物种水分生态型的主要因素, 在缺水区西拉木伦河河岸带的土壤水分条件允许部分中旱生和旱中生植物生长, 而在老哈河河岸带则无中旱生和旱中生植物生长。
3.2 草本植物物种多样性及其与环境因子的关系
老哈河和西拉木伦河的物种多样性均在源头区较低,源头区各样地草本植物群落的物种组成以禾本科植物占优势, 禾本科植物通过根孽繁殖或根茎繁殖等方式繁殖[24],植物群落多呈集群分布[27], 易于形成单优群落, 因此源头区草本植物群落的物种多样性降低。同时, 源头区多为海拔600—1300米的山地, 河谷狭窄, 坡度较大[23], 河岸带较窄, 这些条件均不利于河岸草本植物发育。在从河流源头到下游的纵向梯度上, 环境异质性导致河流的河岸带物种多样性由上游向下游存在差异。有研究认为, 河流中游洪水的干扰强度和持续时间处于中等水平, 其物种较上游和下游的丰富[13, 18, 31], 但也有研究认为自上游至下游物种多样性呈降低的趋势[17]。在本研究中老哈河自源头至与西拉木伦河交汇处, 不断有支流汇入, 水量增加, 河岸坡度变缓[23], 河岸环境变得多样, 提供的生境类型增加, 植物群落的复杂性增高, 从而使得老哈河的物种多样性自上游至下游呈增加趋势。而西拉木伦河不同河段的物种多样性无显著差异, 原因可能是西拉木伦河分支较多, 且在中游有较大支流查干木伦河的汇入, 使得河岸生境更加复杂多变, 物种多样性不稳定。
有研究表明, 河流湿宽越小, 河岸植被物种越少[32],而本研究中物种多样性与河道宽度和湿宽显著负相关,与其研究结果不一致。调查时发现较窄的河流河岸植被较窄, 由于河岸多受到农田干扰, 边缘效应[27]以及人类活动带来的种子增加河岸物种多样性[33]; 田间施肥管理时,加强了河岸的营养富集作用[33],也能引起物种多样性的增加。而在较宽的河流人为干扰多远离水体, 河岸主要为洪水泛滥形成较宽的河漫滩, 而在河漫滩中通常为优势物种形成的优势群落, 因此通常物种多样性低; 而调查时为洪水期过后的平水期, 某些样点河岸植被重新演替,也导致物种多样性低。除河道宽度和湿宽外, 海拔对物种多样性也具有较高的解释率。海拔是影响河岸带植被分布的主导因素,海拔的差异影响洪水周期、水位、土壤基质类型、土壤水分含量、土壤有机质含量和生长季的长短等生境因子[18], 这些因素的综合作用影响河岸带物种多样性。
3.3 东、西辽河河岸带草本物种多样性比较
与东辽河河岸带草本群落的 Shannon-wiener指数(1.58—2.80)、Simpson多样性指数(0.68—0.90)、Pielou均匀度(0.59—0.89)和丰富度指数(8—33)相比[18], 西辽河的物种多样性较低, 这与它们所处的生态区和植被区有关。西辽河流域主要位于辽西北缺水区[29], 植被类型为温带草原[25]; 而东辽河主要位于辽中辽南少水区[29], 植被类型跨温带针阔叶混交林(中上游)和温带草原(下游)[25]。温带针阔叶混交林区地处欧亚大陆东缘, 濒临日本海,受海洋季风的影响, 降雨量较多, 夏季气温较高, 生长季也较长, 气候适宜, 形成独特的长白植物区系, 区域内植物种类丰富; 而温带草原区域海洋季风气候受到山地的阻挡, 成了森林区与草原区之间的分水岭[25], 导致温带草原区的物种多样性较森林区的低。因此, 区域植被背景影响大尺度上河岸带植物的物种多样性格局。
[1] Gregory S V, Swanson F J, Mckee W A, et al. An ecosystem perspective of riparian zones [J]. BioScience, 1991, 41(8): 540—551
[2] Naiman R J, Décamps H. The ecology of interfaces: Riparian zones [J]. Annual Review of Ecology and Systematics, 1997, 28(1): 621—658
[3] Naiman R J, Décamps H, Pollock M. The role of riparian corridors in maintaining regional biodiversity [J]. Ecological Applications, 1993, 3(2): 209—212
[4] Fan M C, Guo R Z, Wu J Q, et al. Floristic analysis of vascular plants in riparian zone of Wujiang River [J]. Journal of Wuhan Botanical Research, 2010, 28(3): 286—293 [樊明策,郭润正, 吴金清, 等. 乌江河岸带维管植物区系的初步研究. 武汉植物学研究, 2010, 28(3): 286—293]
[5] Shen Y Q, Zhang X K, Zhao W H, et al. 2011. Riparian plants in the mainstream of the Yellow River: Assemblage characteristics and influencing factors [J]. Acta Hydrobiologica Sinica, 2011, 35(1): 51—66 [沈亚强, 张晓可, 赵伟华, 等. 黄河干流河岸带植物群落特征及其影响因子分析.水生生物学报, 2011, 35(1): 51—66]
[6] Chen J, Guo Y L, Lu X L, et al. Species diversity of herbaceous communities in the Yiluo River Basin [J]. Acta Ecologica Sinica, 2012, 32(10): 3021—3030 [陈杰, 郭屹立,卢训令, 等. 伊洛河流域草本植物群落物种多样性. 生态学报, 2012, 32(10): 3021—3030]
[7] Huenneke L F. Wetland forests of Tompkins County, New York [J]. Bulletin of the Torrey Botanical Club, 1982, 109(1): 51—63
[8] Sagers C L, Lyon J. Gradient analysis in a riparian landscape: contrasts among forest layers [J]. Forest Ecology and Management, 1997, 96(1): 13—26
[9] Lyon J, Sagers C L. Structure of herbaceous plant assemblages in a forested riparian landscape [J]. Plant Ecology, 1998, 138(1): 1—16
[10] Nichols W F, Killingbeck K T, August P V. The influence of geomorphological heterogeneity on biodiversity II. A landscape perspective [J]. Conservation Biology, 1998, 12(2): 371—379
[11] Dwire K A, Kauffman J B, Brookshire E N, et al. Plant biomass and species composition along an environmental gradient in montane riparian meadows [J]. Oecologia, 2004, 139(2): 309—317
[12] Menges E S, Waller D M. Plant strategies in relation to elevation and light in floodplain herbs [J]. The American Naturalist, 1983, 122(4): 454—473
[13] Jiang M X, Deng H B, Tang T, et al. On spatial pattern of species richness in plant communities along riparian zone in Xiangxi River watershed [J]. Acta Ecologica Sinica, 2002, 22(5): 629—635 [江明喜, 邓红兵, 唐涛, 等. 香溪河流域河岸带植物群落物种丰富度格局. 生态学报, 2002, 22: 629—635]
[14] Brown R L, Peet R K. Diversity and invasibility of southern Appalachian plant communities [J]. Ecology, 2003, 84(1): 32—39
[15] Guo Y L, Lu X L, Ding S Y. The classification of plant functional types based on the dominant herbaceous species in the riparian zone ecosystems in the Yiluo River [J]. Acta Ecologica Sinica, 2012, 32(14): 4434—4442 [郭屹立, 卢训令, 丁圣彦. 伊洛河河岸带生态系统草本植物功能群划分.生态学报, 2012, 32(14): 4434—4442]
[16] Pan X L, Zhang Y, Sun J X, et al. Research on characteristics of riparian herbs in Taizi River, Liaoning [J]. Ecological Science, 2013, 32(5): 616—623 [潘雪莲, 张远, 孙建新, 等.辽宁太子河河岸带草本物种特征. 生态科学, 2013, 32(5): 616—623]
[17] Xia H J, Zhang Y, Zhang W L, et al. Characteristics of plants during early restoration stage of abandoned riparian croplands in Liaohe River reserve [J]. Chinese Journal of Eco-logy, 2014, 33(1): 41—47 [夏会娟, 张远, 张文丽, 等. 辽河保护区河岸农田撂荒恢复初期植物物种特征. 生态学杂志, 2014, 33(1): 41—47]
[18] Zhang W L, Xia H J, Zhang Y, et al. Herbaceous species diversity and community quantitative analysis in the riparian zone of East Liaohe River [J]. Chinese Journal of Ecology, 2014, 33(5): 1142—1149 [张文丽, 夏会娟, 张远, 等. 东辽河河岸带草本植物物种多样性及群落数量分析. 生态学杂志, 2014, 33(5): 1142—1149]
[19] Yu D. Study on the geography of aquatic plants in northeast China [J]. Bulletin of Botanical Research, 1994, 14(2): 169—178 [于丹. 东北水生植物地理学的研究. 植物研究, 1994, 14(2): 169—178]
[20] Yu D. Division of aquatic plants in northeast China [J]. Acta Hydrobiologica Sinica, 1996, 20(4): 322—332 [于丹. 东北水生植物区划. 水生生物学报, 1996, 20(4): 322—332]
[21] Ji Y H, Zhou G S. Transformation of vegetation structure in China’s Liaohe Delta during 1988—2006 [J]. Chinese Journal of Plant Ecology, 2010, 34(4): 359—367 [汲玉河, 周广胜. 1988—2006年辽河三角洲植被结构的变化. 植物生态学报, 2010, 34(4): 359—367]
[22] Zhang Y, Chen L B, Qu X D, et al. Environmental factors and community characteristics of aquatic macrophytes in Taizi River tributaries of Liaoning province [J]. Plant Science Journal, 2011, 29(5): 552—560 [张远, 陈立斌, 渠晓东, 等. 辽宁太子河大型水生植物的群落特征及其与环境的关系. 植物科学学报, 2011, 29(5): 552—560]
[23] Ministry of Water Resources of Songliao Water Resources Commission. Liaohe Zhi [M]. Changchun: Jilin People’s Press. 2004 [水利部松辽水利委员会. 辽河志. 长春: 吉林人民出版社. 2004]
[24] Inner Mongolia Flora Editorial Board. Flora of Inner Mongolia [M]. Hohhot: Inner-Mongol People’s Press. 1998 [内蒙古植物志编辑委员会. 内蒙古植物志. 呼和浩特: 内蒙古人民出版社. 1998]
[25] Chinese Vegetation Editorial Board. Chinese Vegetation [M]. Beijing: Science Press. 1980, 772, 956 [中国植被编委会.中国植被. 北京: 科学出版社. 1980, 772, 956]
[26] Chinese Academy of Sciences, China Flora Editorial Board. Flora of China [M]. Beijing: Science Press. 2004 [中国科学院中国植物志编辑委员会. 中国植物志. 北京: 科学出版社. 2004]
[27] Sun R Y, Li B, Zhuge Y, et al. General Ecology [M]. Beijing: Higher Education Press. 1993, 135—138 [孙儒泳, 李博, 诸葛阳, 等. 普通生态学. 北京: 高等教育出版社. 1993, 135—138]
[28] Lepš L, Šmilauer P. Multivariate Analysis of Ecological Data Using CANOCO [M]. New York: Cambridge University Press. 2003, 51
[29] Meng W, Zhang Y, Zheng B H. Study of aquatic ecoregion in Liao River Basin [J]. Acta Scientiae Circumstantiae, 2007, 27(6): 911—918 [孟伟, 张远, 郑丙辉. 辽河流域水生态分区研究. 环境科学学报, 2007, 27(6): 911— 918]
[30] Strahler A N. Quantitative analysis of watershed geomorphology [J]. Transactions, American Geophysical Union, 1957, 38: 913—920
[31] Nilsson C, Grelssin G, Johansson M, et al. Patterns of plant species richness along riverbanks [J]. Ecology, 1989, 70(1): 77—84
[32] Chen J Q. Riparian vegetation characteristics and their functions in ecosystems and landscapes [J]. Chinese Journal of Applied Ecology, 1996, 7(4): 439—448 [陈吉泉. 河岸植被特征及其在生态系统和景观中的作用. 应用生态学报, 1996, 7(4): 439—448]
[33] Li L T. Distribution pattern of plant diversity and vegetation construction in field margins and homegarden [D]. Doctoral Dissertation. China Agricultural University, Beijing. 2014 [李良涛. 农田边界和居民庭院植物多样性分布格局及植被营建. 博士学位论文, 中国农业大学, 北京. 2014]
CHARACTERISTICS OF RIPARIAN HERBACEOUS VEGETATION AND THE RELATIONSHIP WITH ENVIRONMENTAL FACTORS IN THE WEST LIAOHE RIVER, CHINA
KONG Wei-Jing1, 2, XIA Hui-Juan1, 2, 3, ZHANG Yuan1, 2, HOU Li-Ping1, 2, SUN Li-Hui1, 2and ZHANG Nan1, 2
(1. Laboratory of Riverine Ecological Conservation and Technology, Chinese Research Academy of Environmental Sciences, Beijing 100012, China; 2. State Key Laboratory of Environmental Criteria and Risk Assessment, Beijing 100012, China; 3. MOE Key Laboratory for Silviculture and Conservation of Ministry of Education, College of Forest Science, Beijing Forestry University, Beijing 100083, China)
河岸带; 物种多样性; 水分生态型; 生态区; 西辽河
Riparian zone; Species diversity; Water ecotype; Ecoregion; West Liaohe River
Q948.1
A
1000-3207(2015)06-1266-09
10.7541/2015.166
2015-01-16;
2015-05-11
水体污染控制与治理科技重大专项(2012ZX07501-001-02)资助
孔维静(1978—), 男, 山东泰安人; 博士; 主要从事河岸植被生态和流域生态学研究。E-mail: kongwj@craes.org.cn
张楠(1980—), 男, 江苏徐州人; 博士; 主要从事流域生态学研究。E-mail: zhangnan@craes.org.cn
